
高山生态关键带植物群落多样性格局与系统发育结构
2023-10-08 08:57王俊伟明升平
草地学报 2023年9期
王俊伟, 明升平, 许 敏, 拉 琼*
(1. 西藏大学生态环境学院, 西藏 拉萨 850000; 2. 青藏高原生物多样性与生态环境保护教育部重点实验室, 西藏 拉萨 850000; 3. 西藏雅尼湿地生态系统定位观测研究站, 西藏 林芝 860000; 4. 中国科学院昆明植物研究所云南丽江森林生态系统野外科学观测研究站, 云南 丽江 674100; 5. 西藏自治区林业调查规划研究院, 西藏 拉萨 850000)
沿海拔梯度的植物物种丰富度分布格局一直是群落生态学研究的重要内容。山地生态系统受人为干扰影响较小和栖息地高度异质性,能在有限的垂直地理空间范围内高度压缩地形、海拔和温度等多种环境因子和地形因子于一体,孕育丰富的垂直植被带谱和植物多样性,而且生物多样性热点地区也几乎都在山地生态系统中[1-5]。另外,山地生态系统中往往会表现出沿海拔梯度多种环境变量(如温度、水分、土壤)的梯度变化,因而有很大潜力作为研究气候变化对生物分布影响的天然试验场[4-6]。到目前为止,对山地植物多样性的海拔垂直分布格局还存在很多争论,一直是一个未解决的谜团。普遍得到的植物垂直分布格局是单调递增、单调递减或单峰格局,且这些分布格局在不同的地理区域、不同的研究类群和不同的取样尺度会表现出不同的结果,同时还受到小气候变化和人为干扰强度等很多局部环境因子互作的强烈影响[7-10]。
群落系统发育研究是将物种的演化历史整合到群落生态学研究中,从而可以从多维度探讨物种多样性的垂直分布格局,得出更加准确的研究结果。目前,生态学家的研究都集中于热带、亚热带及温带的森林生态系统和亚寒带的高山草甸。另外随着研究地区的不同、或者大尺度到小尺度的研究系统发育多样分布格局往往表现出不一样的结果。如Qian等对中国森林群落的被子植物树种丰富度与系统发育多样性进行了大尺度格局的研究,发现物种丰富度与系统发育多样性跟温度有密切的关系,且表现出强烈的纬度梯度格局[11]。Cai等对中国木本植物系统发育多样性的地理格局进行大尺度的研究,发现从南到北木本植物物种丰富度与系统发育多样性都下降[12]。如Yue等对中国西南部的亚热带独龙江流域木本被子植物进行了系统发育多样性的研究,发现物种丰富度与系统发育多样性随着海拔的增加而显著降低,系统发育结构在高海拔聚集[13]。Zhang等对中国东部的亚热带森林植物群落沿海拔梯度的分类与系统发育研究,发现不论物种多度加权与否,物种α-多样性均沿海拔升高而增加,对于群落系统发育结构而言,沿海拔增加并无明显变化,得出地形和微气候是多样性格局和群落系统发育结构的主要驱动力[14]。也有在东南亚热带地区的研究发现,物种丰富度随着海拔的升高而下降,系统发育多样性却上升,在高海拔系统发育结构显示出过度分散的格局[15]。黄建雄等对古田山植物群落进行系统发育研究时发现高海拔区域表现出谱系发散格局,而低海拔区域保持谱系聚集[16]。徐璐等对青藏高原东北部五个不同海拔的高寒草甸进行了谱系结构与多样性格局的分析,发现从低海拔到高海拔群落谱系结构由发散到聚集,群落构建由物种趋同进化与生境过滤有关[17]。综上可以得出沿海拔梯度的物种多样性与系统发育多样性分布格局没有一致的结论,对于高海拔生态关键带的山地灌丛群落系统发育的研究更是较为欠缺,特别是青藏高原的高寒山地。
雅鲁藏布江中下游流域是典型的山地生态系统,具有河谷至高山的较为完整的垂直带,独特的自然地理孕育了丰富的植被类型,从河谷山地热带到高山冰缘带的自然植被带在这一区域都有分布[18-20]。因此,这一地区是研究山地植物多样性垂直分布格局的理想场所,也一直倍受众多生物地理学家和生态学家的钟爱。拉琼等[21]对雅鲁藏布江河岸植物群落物种丰富度分布格局做了系统研究,研究结果表明雅鲁藏布江河岸物种丰富度沿海拔和经度呈浅“U”形分布格局,物种丰富度与年均降水量和植被盖度成显著正相关;弓莉等[22]通过群落样地调查与查阅文献相结合对南迦巴瓦兰科植物多样性及垂直分布格局进行了研究,结果发现该区域的兰科植物垂直分布格局呈单峰型,在海拔1 500~2 000 m的区间段上物种丰富度最高;王喜龙等[23]对藏东南兰科植物多样性及沿海拔梯度的分布格局进行了系统研究,结果发现藏东南兰科植物主要集中分布在1 500~3 500 m的海拔段,呈“中间高度膨胀型”的分布格局,从生活型上看也出现同样的分布格局。这些研究极大地丰富了雅鲁藏布江流域的植物物种丰富度分布格局研究基础,但少见有结合物种的进化历史与群落系统发育来研究这一区域植物海拔垂直分布格局的报道。因此,为了了解广泛的物种多样性海拔垂直分布格局,需要更多精细尺度的案例研究做支撑,更是为得出沿海拔梯度的物种与系统发育多样性的普适性格局做基础。
布丹拉山是雅鲁藏布江中下游流域藏南谷地的典型山地生态系统代表,位于藏南河谷东部半湿润区和西部半干旱区的生态群落过渡带[18]。生态过渡带也称群落交错区,由于其受多种要素的联合作用使其物种多样性高而复杂,小地形小环境作用影响强烈。高海拔生态过渡带的植物多样性垂直分布特征也是否像其他常见格局一样,是否也受海拔等关键环境因子的显著影响等这些问题的回答并无一致的结论。为此,本研究选择布丹拉山南坡为研究对象,通过植物群落样方调查分析了布丹拉山南坡种子植物物种多样性、系统发育多样性与系统发育结构的海拔格局,从而推测高海拔山地植物群落构建的生态学过程。旨在探究以下问题:高海拔山地关键生态区植物物种多样性指数与群落系统发育多样性指数之间相关性如何?以及沿海拔梯度多样性指数及群落系统发育结构特征如何?这些分布特征与常见的海拔分布特征是否一致?本研究试图从系统发育的角度探索高山植物多样性的海拔分布格局,结果丰富了雅鲁藏布江中游谷地区山地植物多样性海拔分布格局的研究。
1 材料与方法
1.1 研究区域概况
布丹拉山海拔跨度3 800~5 127 m,正好位于西藏自治区山南市曲松县与加查县行政区划的地理分界线上,南坡地理位置为29°02′~29°03′ N,92°22′~92°36′ E。研究区域是靠近加查县的一侧,山脚是著名大河雅鲁藏布江的小型支流之一拉绥河流域,其分布有密集的农业农事生产活动,行政区划上属于加查县拉绥乡拉索自然村。此区域气候温和,年平均气温8.9℃,年变化相对较小、昼夜日温差变化大,累计年较差14.4℃。平均气温最高月16.4℃(7月),最低月-1.0℃(1月)。气候显著特点是降水集中且雨热同期,蒸发量大[24]。
根据西藏自然地理关于西藏气候、西藏地形和西藏植被分区的描述,此区域地理位置特殊,正好处于好几个自然地理分带的生态过渡交错区,从西藏地形分区上看处于喜马拉雅山区向雅鲁藏布江中游谷地区的过渡带上;从西藏气候地域类型分区上看处于藏东南温暖半湿润高原季风气候地区向藏南温暖半干旱高原季风气候地区的过渡带上;从西藏植被区划上看处于藏东山地针叶林区向藏南山地灌丛草原区过渡的区域[18]。另外此区也是青藏高原重要山脉冈底斯山—念青唐古拉山向喜马拉雅山的东部延伸的地理缝合区[18],使这一区域地形复杂多样,海拔高低悬殊,气候条件多样,水热再分配的垂直地带性规律明显,孕育了复杂的垂直自然植被带和丰富的山地植物多样性。
1.2 植物群落野外本底调查
调查时间分2018年9月和2019年9月分两次对布丹拉山南坡进行植物群落野外样方调查,2020年5月至2020年7月分两次又对布丹拉山南坡进行野外植物标本的采集与鉴定的补充调查工作。调查范围从山脚农田边缘到山顶高山流石滩,在1 327 m的海拔跨度上,每隔50 m设置一个海拔样带,在每个海拔样带上随机设置6个5 m×5 m的平行样方,共调查了162个样方(27个海拔样带×6块样地)。调查内容详细记录每个样方内的种子植物物种组成及海拔样带内未在样方内的周围物种,目测样方内每个物种的盖度和样方的总盖度,禾本科(Poaceae)与莎草科(Cyperaceae)等禾草类植物用盖度代替多度。用GPS全球定位系统仪和罗盘仪记录每个样方的海拔、经纬度与坡度等地形因子。
1.3 数据处理与分析
1.3.1系统发育多样性与群落系统发育结构计算 基于162个调查样方记录的298种植物名录数据为蓝本,整理成科/属/种格式的植物名录,再利用Jin和Qian开发的R语言V. PhyloMaker 2软件包[25-26],使用phylo. maker函数构建本研究的超级系统发育树,此合成树是基于现有的超级系统发育树裁剪拼接而成(图1)。基于合成的超级系统发育树再利用R语言Picante软件包计算系统发育多样性指数(Phylogenetic diversity,PD)、系统发育结构指数用净亲缘关系指数(Net relatedness index,NRI)和净最近亲缘关系指数(Net nearest taxa index,NTI)来表征[27]。系统发育结构指数计算具体采用的方法是:通过群落物种库合成的超级系统发育树,再将实际的每个群落与标准化的零模型作比较,统计实际观测值与零假设期望值间的差异。零模型的构建采用群落中所有物种于系统进化树末端分支随机置换999次的方法,保持每次零模型和生成的系统进化树具有相同的物种数目。NTI指数侧重于从分支末端衡量群落系统发育结构,受系统发育树末端多歧分支的影响较大,而NRI指数侧重于从系统发育树整体上来衡量群落谱系结构[27]。具体的计算公式如下:

图1 布丹拉山南坡海拔梯度298个种子植物的系统发育树
PD=∑Lb
式中,Lb为系统发育树量化后的分支长度,MPDs和MNTDs分别表示实际观测的MPD值和群落内亲缘关系最近种间的平均系统发育距离,MPDmds和MNTDmds分别表示999个零模型模拟下随机群落的MPD值和最近相邻系统发育距离的平均值,SD为标准差。若NRI>0,NTI>0,则小样方的群落系统发育结构聚集,表明亲缘关系近的物种倾向于在同一个群落中出现,群落中环境过滤作用占主导;若NRI<0,NTI<0,则小样方的群落系统发育结构发散,表明群落中的物种亲缘关系较远,竞争排斥的生态过程占主导作用;若NRI=0,NTI=0,则表明群落的系统发育结构是随机的。所有的NRI值和NTI值计算时都进行物种多度加权。
1.3.2物种多样性指数计算 根据调查得到的样方物种矩阵数据,采用物种丰富度(Species richness,S)、Shannon-Wiener多样性性指数(Shannon-Weiner diversity index,H)、Simpson优势度指数(Simpson index,D)和Pielou均匀度指数(Species evenness index,E)来分析和评价物种多样性沿海拔梯度的差异[28]。计算公式如下:
S=样方内物种总数
式中:Pi表示第i个物种的多度比例。
1.3.3海拔分布格局分析 广义线性模型、回归分析与Pearson’s相关性分析物种丰富度(S)、Shannon-Wiener指数(H)、Simpson优势度指数(D)和Pielou均匀度指数(E),以及群落系统发育多样性指数(PD)、系统发育结构净亲缘关系指数(NRI)与净最近亲缘关系指数(NTI)的海拔梯度变化趋势。
2 结果与分析
2.1 植物群落物种组成特征
162个样方共记录到了种子植物298种,隶属于45科149属,以草本植物为主,学名以英文版中国植物志FloraofChina为准(Editorial Committee of Flora of China,1994—2013)。不同群落样方间物种丰富度差异很大,物种数最少的为10种,物种数最多的为49种。出现频率最高的物种是钉柱委陵菜(Potentillasaundersiana),总共162块样方,100块样方中都有记录;93块样方记录了高山嵩草(Carexparvula);83块样方记录了圆穗蓼(Polygonummacrophyllum);80块样方记录了矮羊茅(Festucacoelestis);77块样方记录了吉拉柳(Salixgilashanica);75块样方记录了倒披针叶风毛菊(Saussureanimborum)。
2.2 群落物种多样性与系统发育多样性的海拔格局
通过162个样方的种子植物物种丰富度、Shannon-Wiener多样性指数、Simpson优势度指数和Pielou均匀度指数与海拔梯度的回归分析与显著性检验(图2),发现布丹拉山南坡种子植物物种丰富度与海拔梯度(图2a)没有显著的相关性,相关系数r值为0.118,也无明显的海拔分布格局。群落物种Shannon-Wiener多样性指数(图2b)、Simpson优势度指数(图2 d)和Pielou均匀度指数(图2c)与海拔梯度均呈现显著的正相关(P<0.05),相关系数r值分别为0.242,0.272,0.301,其变化规律随着海拔的增加而呈显著单调递增的格局。群落系统发育多样性指数沿海拔梯度的变化规律不明显,值范围在1 173.514~2 994.893之间,其与海拔梯度无显著相关关系(图3c),相关系数r值为-0.029。
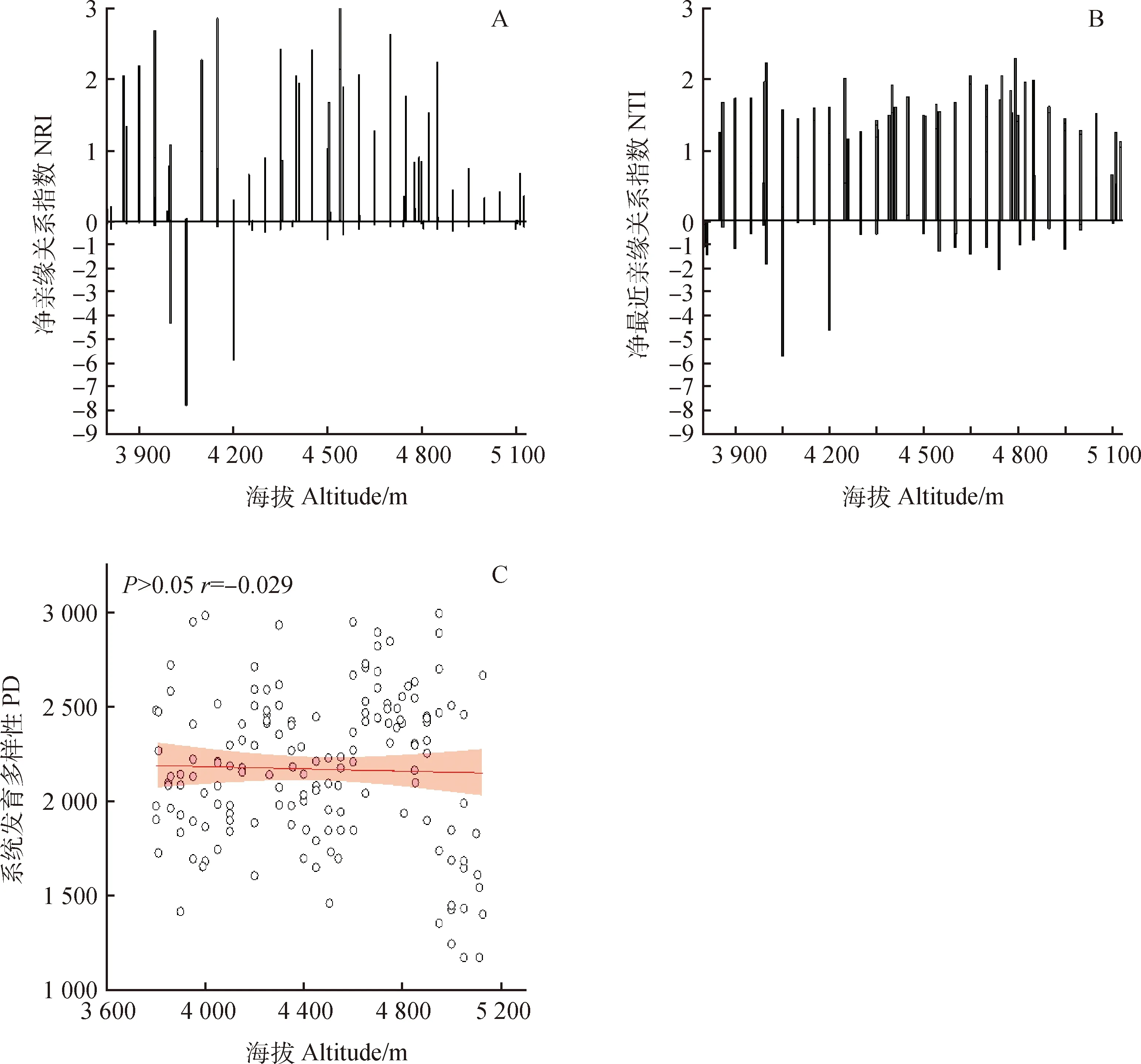
图3 沿海拔梯度的群落系统发育结构(a,b)、群落系统发育多样性(c)变化图
2.3 群落系统发育结构沿海拔梯度的变化规律
图1表示布丹拉山植物群落系统发育关系,描述了物种间进化亲缘关系。净亲缘关系指数(NRI)和净最近亲缘关系指数(NTI)是表征群落系统发育结构最常用的两个指标。结合表1和图3,布丹拉山南坡沿海拔梯度的群落系统发育结构聚类或过度分散都没有明显的线性变化趋势,可以看出布丹拉山南坡整个山脚到山顶的海拔梯度上所调查的162个样方中,大多数的样方植物群落系统发育结构指数NRI与NTI两者都大于0,即在整个海拔梯度群落系统发育结构主体上都呈现聚集的状态,个别样方系统发育结构指数NRI与NTI都小于0,出现群落系统发育结构发散的格局。其中,低海拔3 800~4 000 m的区间有6个、中海拔4 000~4 500 m的区间有8个、高海拔4 500~5 000 m的区间有12个、5 000 m以上的极高海拔有4个植物群落样方表现系统发育发散的结构,即组成这些植物群落的物种之间亲缘关系都相对较远,其群落构建中相似性竞争的作用更强烈。如表1中统计可知,每一个海拔梯度上都有一半以上的植物群落样方系统发育结构指数NRI>0,NTI>0,都是群落系统发育聚集的格局,即亲缘关系较近的物种共存组成植物群落,表明环境过滤的生态作用对布丹拉山南坡植物群落的构建起主导作用。

表1 沿海拔梯度的样方系统发育结构指数
2.4 物种多样性与系统发育多样性指数间的相关性分析
布丹拉山南坡植物群落物种多样性指数(S,H,D)与群落系统发育多样性指数(PD)均极显著正相关;物种多样性指数之间除了均匀度指数外都表现出高度的协同性;另外,物种多样性指数(S)与群落系统发育多样性指数(NTI)也有显著正相关的关系(表2)。Shannon-Weiner多样性(H)指数与S,E和D间极显著正相关,即物种丰富度、均匀度、优势度均对物种多样性具有极显著的贡献。系统发育多样性结构指数NRI和NTI与物种多样性(S,E,H,D)之间均无显著相关性。系统发育多样性结构指数NRI和NTI之间有极显著的正相关性。
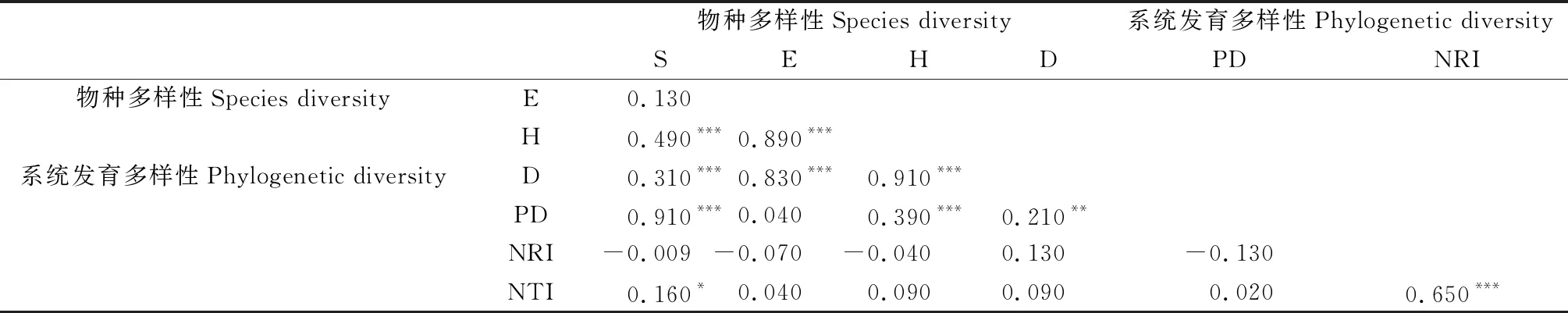
表2 布丹拉山南坡植物群落α多样性指数间的Pearson相关系数
3 讨论
物种海拔分布格局的关系在很大程度上是依赖于环境变量之间的变异程度(小地形和小生境),特别是地形复杂多样,微环境作用显著的山地生态系统中尤为明显。小地形对海拔梯度变化所引起的水热条件有强烈的再一次修饰的作用[29]。处于许多自然地理分界的生态过渡带和山脉缝合带的布丹拉山,地形复杂多样,气候条件变化多端,直接影响了这一区域水热因子的再一次分布变化,因此造成了这一区域特殊的植物分布格局及成分复杂多样。有很多研究也发现不同的生活型植物表现出的物种丰富度海拔格局都不一样[30-31],而草本植物对环境梯度变化是最敏感的[29,32],因此草本植物受局域小环境的影响比较大,甚至没有一个气候变化因子与草本植物丰富度有显著相关[33],也有研究者发现总的物种丰富度或草本植物物种丰富度与海拔没有显著相关性[33-35]。草本植物物种丰富度与环境的关系是非常复杂的[36],因其短暂的生活史[7]。本研究中总的物种丰富度随海拔的关系没有显著相关性,可能与布丹拉山南坡的种子植物群落中草本植物占据绝对优势,近90%的物种是草本植物的物种组成结构有一定关系。另外,从结果分析可知样方之间的物种丰富度也存在显著差异,群落系统发育结构沿海拔梯度的分析也无明显的变化规律,说明影响物种多样性分布的因素非常复杂,可知此区域的植物在空间垂直分布格局上可能更多的受小环境和小地形因子作用(低洼、岩石缝、人为干扰和放牧等)。
多重环境指标和能量指标是物种分布的重要驱动因素,当前的气候变化也对物种丰富度空间分布格局影响显著[37-38]。土壤的特性,如全氮、全钾、全磷、土壤含水量、有机质、pH值等土壤环境因子也对物种分布有着重要影响[39],而本研究中只分析了物种多样性、系统发育多样性与海拔之间的相关性,环境因子与地形因子的测度较少。因此在今后的研究中应该尽可能多的考虑多元环境指标对山地植物物种多样性分布格局潜在机制的影响,可为今后评估全球气候变化对物种分布可能的影响提供相关基础资料。高海拔地区较为恶劣的生境条件使环境过滤占据主要地位,环境筛选出布丹拉山南坡杜鹃属、柳属为优势的植物群落,系统发育结构主体上表现为相对聚集[17,20]。但在布丹拉山南坡4 500 m的高海拔及以上也有16个植物群落在样方尺度上出现群落系统发育发散的格局,我们猜测布丹拉山南坡作为生态过渡带生境高度异质性,高海拔地区形成了更多的小生境小地形为更多的物种生存提供生境条件,促使群落物种间采取不同的资源利用策略,再加上高海拔植物可利用资源有限,种内竞争更加激烈,使得高海拔一些小区域群落系统发育结构发育为发散的格局,倾向于更多亲缘关系远的物种共存组成群落。目前,基于生态位理论的环境过滤和竞争排斥被认为是群落构建过程中的主要驱动力。一般而言,环境过滤的作用会使得相同生境出现亲缘关系较近、适应能力相似的物种,系统发育群落结构会表现为聚集。相反,竞争排斥是不同的物种竞争同一资源,因为亲缘关系较近、生态位相似的物种无法共存于同一环境中,因此群落内的物种会表现为系统发育发散的格局[40-41]。我们的研究结果布丹拉山南坡中低海拔有几个样方群落系统发育呈现异常发散的格局,几个样方中NRI与NTI达到-6甚至-8,是因为这些样方中的植物群落共存有亲缘关系较远的物种有关,野外调查也发现这几个样方中有裸子植物、禾本科等单子叶植物和超菊类分支的植物共存一个空间,而这些植物在系统发育关系上与伴生植物的系统位置关系都较远,因此会导致这种系统发育异常发散的格局。
群落系统发育结构沿海拔梯度由发散到聚集的变化,已被很多研究证实[13,15,17],而我们的研究在海拔梯度上并没有表现出明显的变化规律,但群落系统发育结构主体是聚集的。我们分析推测可能与我们研究区域的海拔起点就很高的原因有关。以往报道的群落系统发育结构由发散到聚集的变化格局大多是在海拔3 000 m以下的研究中报道。而我们在布丹拉山进行的群落系统发育的研究海拔跨度为3 800~5 127 m,所以我们没有发现这种由发散到聚集的变化格局,整体都是聚集的格局。这也证明了青藏高原高海拔地区的植物群落构建机制以生境过滤作用为主,致使高海拔地区的植物群落系统发育结构聚集。
另外,我们的研究中系统发育结构NRI与NTI指数不一致的样方也占据一定的比例,从一定程度上也说明了布丹拉山南坡的植物群落还在不断的演化与发展中,植被结构还没有达到相对稳定的程度[42]。群落中存在一些易受威胁影响或狭域特有分布的物种,当这些物种丧失后,群落极其脆弱,群落稳定程度会下降,致使系统发育结构没有表现出明显的聚集和发散趋势[42]。同时,目前选择的零模型也不完善,应该全面考虑群落中的物种丰富度和多度以及生态学特征,并将进化特征和群落生态学功能性状特征结合起来[43],因此,将来需要更多的研究来验证NRI和NTI指数的准确性并寻找更加合理的模型,增加统计分析和解决问题的能力。另外,有研究也表明不同的群系由于不同建群种对环境适应策略的差异会导致不同群系的分布范围和物种组成有显著不同,这种会在群落的系统发育结构上体现出差异[44],甚至会得出相反的群落构建机制[45]。因此后续应该分不同的植被类型或群系、不同的垂直结构层与不同生活型植物来研究群落系统发育结构及其分布格局。
群落系统发育多样性与物种丰富度之间存在极显著的正相关关系[14]。我们的研究结果也得到了一致的结果,其中Shannon-Wiener多样性指数与其他物种多样性指数、系统发育多样性指数都显著相关,因此在有些时候可以代替其它指数对植物群落结构进行描述。但是系统发育结构NRI指数、NTI指数与Shannon-Wiener多样性指数、Simpson优势度指数和Pielou均匀度指数都没有显著相关,与吕自立等在巴音布鲁克高寒草甸植物群落研究得出系统发育结构指数净最近种间亲缘关系指数(NTI)和净谱系亲缘关系指数(NRI)与物种多样性指数均呈显著负相关关系(P<0.05)的研究结果不一致[46]。这可能是由于布丹拉山物种多样性高,很多物种还在处于分化初期阶段导致的[47],另外我们推测也通过拼接合成的超级系统发育树研究物种丰富多样的生态群落交错区可能效果不是很理想。也有研究提出了这样的观点,Jin等人对中国亚热带常绿阔叶林的580种木本植物进行群落系统发育研究表明,通过合成的系统发育树来进行群落系统发育结构的研究可能会造成偏差,尤其是属种一级的分支关系,因此可能得出错误的结论[48]。
海拔和地形(坡度和坡向等)是影响植物丰富度的最重要因素[49],并且各植物生活型类群的响应变量不同[50]。对物种丰富度和海拔之间关系的确切形式的变化的一些可能的原因进行了很多的研究评论,但是我们对海拔和物种丰富度之间的关系的理解仍然是不够成熟的[51]。本研究反映出环境梯度复杂的高原山地生态系统中,特别是处于这种生态群落交错带的过渡区,海拔对山地植物多样性的分布格局可能没那么重要了,而是与所处的小生境类特征、植被类型和大量复杂的生态环境因子有着很大的联系,坡度等地形异质性引起的小生境作用是布丹拉山南坡种子植物物种丰富度分布格局强有力的解释变量。我们的研究结论正好也验证了经典生态学教材《基础生态学》中关于生态过渡带是多种要素的联合作用和转换区,各要素相互作用强烈,常是非线性现象显示区和突变发生区,是生物多样性较高的区域的观点[52]。
鉴于本研究中有些样方系统发育结构NRI和NTI反映出来的趋势存在差异,加之本研究缺少环境因子、土壤因子的取样,系统发育也是基于合成的系统发育树来研究的。因此对于物种高度多样且丰富的布丹拉山地区需要借助质体基因组学、DNA条形码技术等更加精准的手段来构建分辨率更好更高的系统发育关系树来研究群落生态构建的过程,以加速了解高海拔地区群落的系统发育结构及其驱动因素,特别是青藏高原地区,后续还需要通过加大更多的取样,以及研究尺度的增加并结合植物功能性状进行多维度综合的研究,进一步深入探讨高山关键带生态群落交错区植物群落构建的形成与机制。我们的研究以布丹拉山南坡为研究对象,通过大量的野外植物群落样方调查数据,从物种多样性、系统发育多样性与系统发育结构等方面探讨了山地植物群落多样性的海拔格局及其群落构建机制,但本研究还存在一些不足,有研究发现影响高寒草甸植物群落多样性的主要环境因子是土壤水分、速效磷和铵态氮等土壤因子[53],我们的研究正是缺少对应的土壤因子、环境气候变量数据,限制了定量的进行深入分析研究,展望这些不足将在后续研究中进一步探索和改进。
4 结论
布丹拉山南坡种子植物群落物种多样性丰富,其中草本植物占据优势。沿海拔梯度群落系统发育结构主体上表现为聚集,说明高海拔山地植物物种多样性高,但都是由亲缘关系较近的物种组成,表明生境过滤的生态作用是高海拔生态过渡带的山地植物群落构建的主要驱动力。高海拔地区物种丰富度、群落系统发育多样性与系统发育结构的沿海拔分布格局受许多环境变量共同影响,与海拔都没有显著的相关性。高海拔关键生态区的物种多样性海拔垂直分布格局与常见的物种单调递增或递减与单峰分布格局不同。
相关性分析表明关键生态区物种多样性Shannon-Wiener指数与其他物种多样性指数、系统发育多样性指数(PD)之间都显著存在相关性,系统发育结构指数NRI和NTI与物种多样性(S,E,H,D)之间均无显著相关。
猜你喜欢
林业勘查设计(2022年1期)2022-02-15
散文诗(2021年22期)2022-01-12
乡村科技(2021年17期)2021-10-20
安顺学院学报(2021年4期)2021-09-16
林业调查规划(2020年3期)2020-06-03
今日农业(2019年13期)2019-08-12
环球时报(2019-05-23)2019-05-23
人民调解(2019年5期)2019-03-17
少儿美术(快乐历史地理)(2018年7期)2018-11-16
小说月刊(2015年12期)2015-04-23
