
晋北农牧交错带草地土壤净氮矿化速率对氮磷添加的响应
2023-10-08 09:24杨倩雯赵芳草刁华杰董宽虎王常慧
草地学报 2023年9期
韩 菲, 苏 原, 杨倩雯, 王 袼, 常 婕,赵芳草, 刁华杰, 董宽虎*, 王常慧*
(1.山西农业大学草业学院, 山西 太谷 030801; 2.草地生态保护与乡土草种质创新山西省重点实验室, 山西 太谷 030801;3.山西右玉黄土高原草地生态系统国家定位观测研究站, 山西 右玉 037200)
氮(N)和磷(P)是生命元素,参与陆地生态系统生物地球化学循环关键过程,同时也是限制植物生长的大量元素[1]。氮磷养分的平衡对于养分限制地区植物的生长至关重要,同时氮、磷可用性的高低对于维持陆地生态系统的结构和功能具有重要作用[2]。根据最新报道,全球氮沉降量持续增加,已经超过了90 Tg N·a-1[3]。土壤中合适的氮磷比可以维持较高的生产力[4]。大量的研究已经报道了外源氮添加引起的氮磷失衡通过影响土壤微生物[5]影响生物地球化学过程[6],以及土壤氮的转化过程[7]。但是单独的外源磷输入和氮磷的交互作用如何影响农牧交错带草地生态系统土壤氮的转化过程目前还不清楚,由于农牧交错带草地氮沉降的量高于天然牧区草地[8],同时受农田P输入的影响,因此研究农牧交错带草地外源N,P输入对土壤氮转化速率的影响对于科学管理和利用该区域草地具有重要意义。
土壤中N素可利用性的高低可以用土壤净N矿化速率表征[9]。已有的大量研究表明早期N添加促进土壤净N矿化速率[10-13],长期N添加对净N矿化速率无显著影响[14-16],或抑制净N矿化[17]。外源P输入可以通过影响土壤微生物的活性和功能直接影响土壤无机氮库和氮的矿化过程[18]。在热带森林生态系统[19]、草原生态系统[20]、和湿地生态系统中的研究结果表明P添加促进了土壤净氮矿化速率[19-22]。也有研究报道在热带低地雨林生态系统P添加对土壤净N矿化速率没有显著影响[23],新西兰草地生态系统P添加导致净N矿化速率降低[24]。但是,目前关于农牧交错带草地土壤净N矿化作用对P添加及N和P同时添加响应方面的研究仍非常有限,限制了我们对不同区域、不同类型草地生态系统养分循环的认知和管理水平的提升。
北方农牧交错带草地受人为干扰影响较大,同时对气候变化极其敏感[25]。有关外源N,P添加对农牧交错带草地土壤净N矿化速率影响的研究报道还不多见,由于其所处地理位置的特殊性,因此需要深入了解农牧交错带草地受氮沉降和农田P输入的影响,从而科学管理该区域的草地。我们依托2018年在山西右玉黄土高原草地生态系统国家定位观测研究站建立的全球变化联网试验——养分添加试验平台,研究N,P添加及其交互作用对农牧交错带草地土壤净N矿化速率的影响。
1 材料与方法
1.1 研究区域概况
本研究依托山西右玉黄土高原草地生态系统国家定位观测研究站(112.29°E,39.28°N) 2018年建立的全球变化联网试验平台(右玉站)展开,该实验地点海拔 1 348 m,年平均降水量425 mm,年平均蒸发量1 000~1 500 mm,年平均气温4.6℃,属于温带大陆性季风气候,位于北方典型农牧交错带,植物群落以赖草(Leymussecalinus)、碱茅(Puccinelliadistans)、碱蒿(Artemisiaanethifolia)等植物为优势种。
1.2 试验设计
试验采用完全随机区组设计,每个处理6个重复,包括4个处理,对照(CK)、N添加(N)、P添加(P)和N,P同时添加(NP),共计24个小区,小区面积6 m×6 m,小区间间隔为2 m。土壤N矿化速率测定时间分为非生长季(2020年10月至2021年4月)和生长季(2021年5月至2021年9月)。在生长季初期(5月初)将10 g·m-2·a-1的缓释尿素和10 g·m-2·a-1的Ca(H2PO4)2分别均匀的撒施到处理小区。
1.3 取样与处理

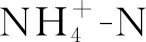
Δt=t1-t0
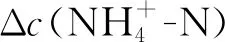
采用收获法测定植物地上生物量,2021年8月在每个处理小区内随机取0.2 m ×1 m的固定样方,用剪刀将植物地上部分齐地面剪下,装入信封并收集凋落物,带回实验室在65℃烘箱中烘干48小时候称重,计为地上生物量。
使用便携式土壤水分速测仪(TDR-300,Spectrum)测定土壤含水量,将探针插入土壤10 cm处记录数值;土壤温度用数字显示温度计(JM-224,天津今明)测定,将探针插入土壤10 cm处测定并记录数值,生长季每周测定一次,同时测定土壤含水量与土壤温度。空气温度及降雨量数据从实验站的小型气象站获得(HOBO U30-NRC,USA,Onset)。
1.4 数据处理
用双因素重复测量方差分析法(Two-way repeated ANOVA)分析N添加和P添加对土壤温度、土壤含水量、铵态氮、硝态氮、无机氮,净铵化速率、净硝化速率、净氮矿化速率的影响。采用单因素方差分析法(One-way ANOVA)检验,分析不同月份、生长季与非生长季N添加和P添加对无机氮,净氮矿化速率的影响,N添加和P添加对植物地上净初级生产力的影响。采用相关性分析检验无机氮、净氮矿化速率与地上净初级生产力的相关性,数据分析使用SPSS 16.0(SPSS,Chicago,USA),用Origin 2021作图。
2 结果与分析
2.1 环境因子
2021年生长季(5—9月)月平均空气温度呈单峰曲线,最高值出现在7月(24.7℃),最低温出现在5月(6.7℃)。2021年降雨量分布不均匀,八月降雨量最高为94.6 mm(图1)。
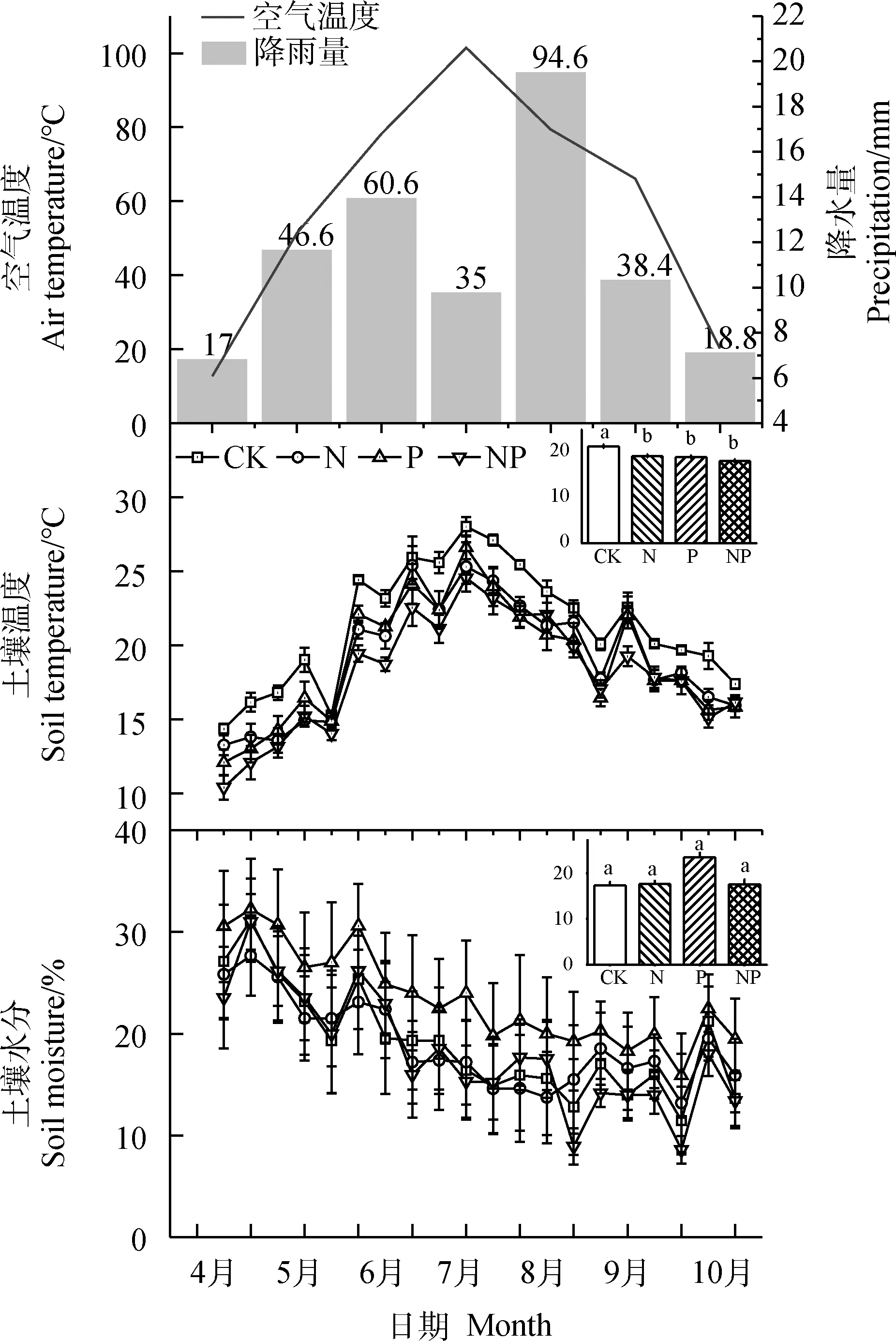
图1 2021年空气温度、降雨量、土壤温度和土壤水分的季节变化
0~10 cm的土壤温度和土壤含水量呈现出明显的季节波动(图1),季节之间差异显著(P<0.01;表1)。N,P单独添加和同时添加显著降低了生长季土壤的平均温度(P<0.05;图1)。土壤含水量月份之间无显著差异(表1),2021年生长季平均土壤含水量为19.84%。各处理间土壤含水量无显著差异(表1)。

表1 氮磷添加对土壤温度、土壤含水量、无机氮含量、氮转化速率月份变化影响的双因素重复测量方差分析结果
2.2 氮、磷添加对土壤铵态氮、硝态氮和无机氮含量的影响
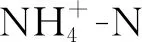
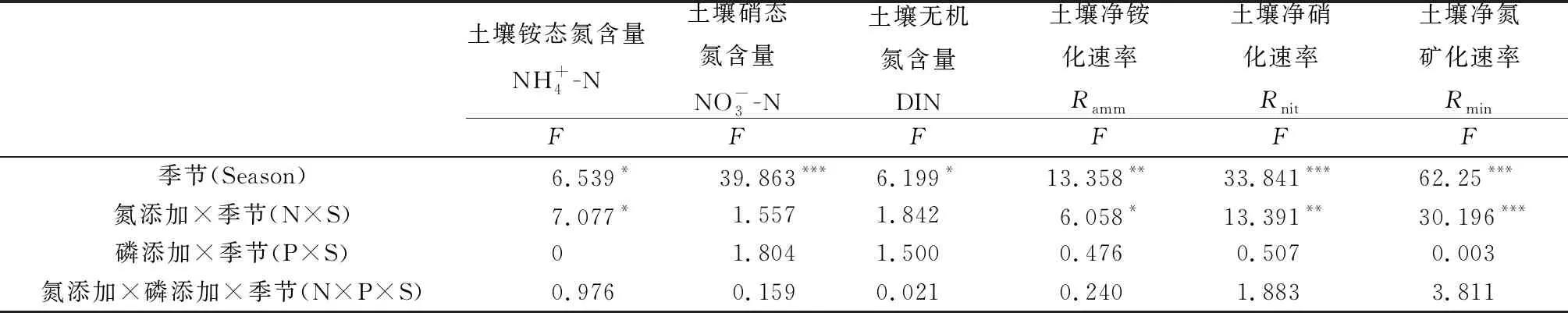
表2 生长季和非生长季氮磷添加对土壤无机氮含量、氮转化速率季节变化影响的双因素重复测量方差分析结果
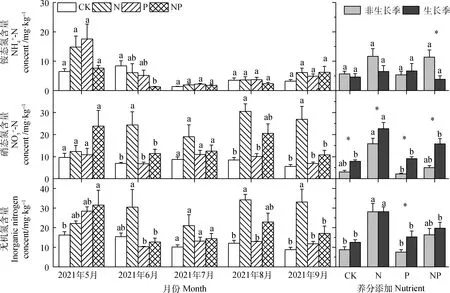
图2 2021年不同处理下土壤无机氮含量的季节变化

图3 2021年不同处理下土壤氮转化速率的季节变化
氮添加极显著地影响土壤无机氮含量(P<0.001;表1),P添加对土壤无机氮含量影响不显著(表1)。NP添加对土壤无机氮含量影响显著(P<0.05;表1),8,9月N添加显著增加了土壤无机氮含量(P<0.05),比对照分别增加了298.4%,194.9%。不同的N,P添加处理在非生长季和生长季对土壤无机氮含量的影响差异显著(P<0.05;表2)。非生长季P添加处理的土壤无机氮含量显著低于生长季(P<0.05),低于生长季205%。
2.3 氮、磷添加对土壤净铵化速率、净硝化速率和净氮矿化速率的影响
净铵化速率、净硝化速率和净氮矿化速率表现出明显的季节动态(图2),生长季(5月—9月)内单独N,P添加处理对土壤净铵化速率没有显著影响(表1)。7月N添加显著增加了土壤净铵化速率(P<0.05);5月P添加显著降低了土壤净铵化速率(P<0.05);6月氮磷添加使得土壤净铵化速率显著提高了327.5%(P<0.05)。不同的N,P添加处理在非生长季和生长季对土壤净铵化速率的影响显著(P<0.01;表2)。生长季NP同时添加处理的土壤净铵化速率显著高于非生长季(P<0.05)。
氮添加极显著地提高了土壤净硝化速率(P<0.001;表1)。P添加、NP同时添加对土壤净硝化速率没有产生显著影响(表1)。非生长季、5、7月N添加显著提高了土壤净硝化速率(P<0.05);生长季末期的9月,氮磷同时添加显著提高了土壤净硝化速率(P<0.05);非生长季和生长季土壤净硝化速率的差异在单独N,P添加处理下达到极显著水平(P<0.001;表2)。在生长季,不同处理的土壤净硝化速率均显著高于非生长季(P<0.05)。
N添加极显著地提高了土壤净氮矿化速率(P<0.001;表1)。但是单独P添加和N,P同时添加对土壤净N矿化速率影响不显著(表1)。7月N添加显著提高了土壤净N矿化速率(P<0.05)。6月和9月NP处理显著增加了土壤净N矿化速率,比对照分别增加了1 195.8%,469.8%(P<0.05)。单独的N,P添加处理在非生长季和生长季对土壤净N矿化速率的影响差异显著(P<0.001;表2)。生长季单独添加N,P和NP同时添加处理的土壤净N矿化速率均显著高于非生长季(P<0.05)。
2.4 氮、磷添加对地上净初级生产力的影响
不同处理之间的土壤净初级生产力差异显著(P<0.05),单独的N,P添加处理均对地上净初级生产力没有产生显著影响,但是N,P同时添加处理显著增加了地上净初级生产力(P<0.05,图4)。N添加处理下,土壤铵态氮的含量与地上净初级生产力呈显著正相关关系(P<0.05,图5),土壤硝态氮含量与土壤无机氮含量与地上净初级生产力相关性均不显著(图5)。且在N添加处理下,土壤净铵化速率与地上净初级生产力呈负相关关系(P<0.05,图5);N,P同时添加处理下,土壤净硝化速率与净N矿化速率与地上净初级生产力呈负相关关系(图5)。
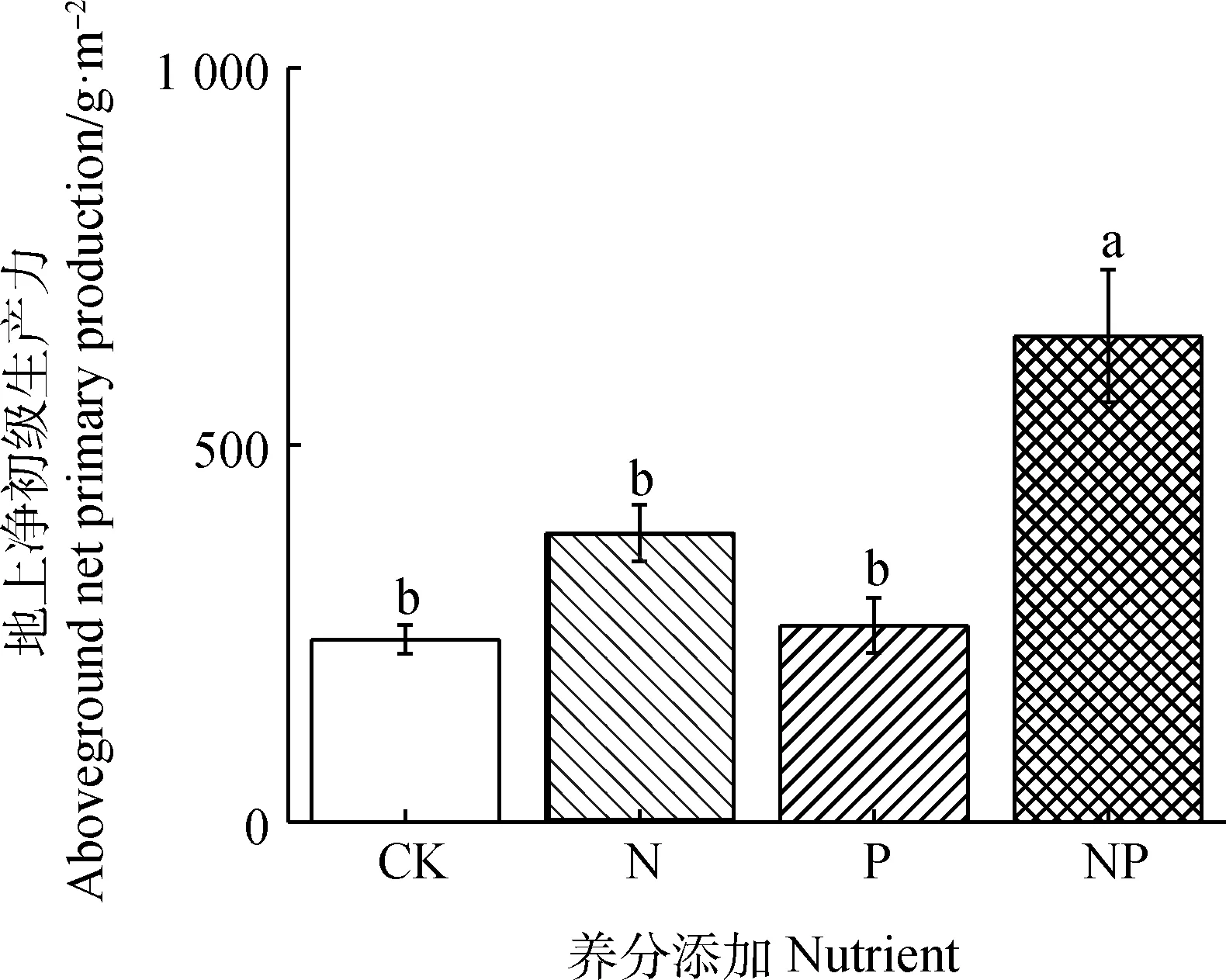
图4 2021年氮磷添加处理对植物地上生物量的影响

图5 地上生物量与氮转化速率的相关关系
3 讨论
3.1 氮添加对土壤净氮矿化速率的影响
我们发现N添加显著增加了生长季土壤无机氮含量,尤其是硝态氮含量,且净硝化速率、净氮矿化速率显著增加,这与大部分草地生态系统的研究结果一致[10]。在受氮限制的生态系统中,氮沉降能够直接提高土壤可利用氮含量[27],增强硝化作用[28],刺激土壤氮矿化[23],在松嫩平原[26]和南非稀树草原[12]的研究也得出相同的结论。一方面N添加导致土壤氮富集,当土壤中的无机氮相对于微生物的需要量过剩时,可能短暂增加了土壤微生物活性[29],土壤微生物释放无机氮[30],导致了土壤有机质的矿化。另一方面N添加导致土壤C:N比减少,尤其是当基质碳氮比趋于微生物量碳氮比时,净氮矿化量显著增加[31]。也有研究表明N添加对土壤净氮矿化速率无显著影响,或起抑制作用[31]。我们的试验处理开始于2018年,在进行试验处理之前,没有外源养分输入。由于短期N添加与长期N添加对土壤净N矿化速率的影响存在差异[19],随着N添加时间的不断延长,土壤净氮矿化速率逐渐降低[32]。
3.2 磷添加对土壤净氮矿化速率的影响
3.3 氮磷交互作用对土壤净氮矿化速率的影响
在本研究中,NP同时添加对土壤无机氮含量均没有产生显著影响,与对照相比N,P同时添加导致土壤净N矿化速率和净硝化速率增强,但低于单独N添加的处理。该研究结果与已有的研究结果不一致,氮磷的互作效应使土壤氮矿化和硝化速率显著高于单独N添加处理[1]。造成这种差异的原因:当生态系统达到N饱和时,P成为植物和土壤微生物的主要限制因子[32];植物生长和微生物代谢从氮限制转向P限制时,尤其是当植物生长的氮素限制解除后,土壤磷酸酶活性可以通过N添加而提高,因此,添加P会增加植物生长和土壤微生物活性[38]。植物生长的加快会刺激植物对氮的吸收,微生物生物量和活性的增加会增加氮的矿化和其他转化过程。当生态系统氮含量较低时,即使同时添加氮和磷,植物生长[24]和微生物代谢仍可能受氮限制[39],PII(2015)等的研究也得出相似的结论,在氮限制的生态系统中,N添加主要用来维持植物和微生物的生长,而不是磷酸酶的产生[40]。我们研究区土壤养分贫瘠,不仅受氮限制,同时也受到磷的限制。在酸性土壤中的研究发现,在氮磷同时添加和单独磷添加的处理小区,土壤pH值高于单独N添加小区,表明P添加减轻了N添加引起的土壤酸化,土壤酸化的降低为微生物和植物的生长提供了更有利的条件[19]。另外,N添加导致的土壤pH值降低有助于金属离子如铝、钙、铁等激发磷的活性,从而随着时间的推移减轻磷限制,但是对植物和微生物没有影响[41],所以磷限制的减缓只是暂时的[21]。Luisin等[42]在巴塔哥尼亚草原的实验证明了这一观点。另一方面本研究P添加的量较低或添加时间较短,短期氮磷添加还不能完全反映出其对土壤净氮矿化速率的影响[11],需要进一步研究揭示草地土壤氮素净矿化量特征及其影响机理。值得注意的是,氮磷同时添加显著增加了植物净初级生产力,这与何利元等2015年在内蒙古温带草地发现氮磷同时添加提高了植物地上生物量90%以上的研究结果相似[43]。一方面可能是磷添加缓解了植物对氮素吸收的限制[44],Eisele和Crews等发现增加氮的可利用性,导致植物生产力显著增加[45-46]。另一方面可能是磷添加提高了土壤磷的供给能力,缓解了氮沉降而引起的土壤和植物间养分不平衡[47]。同时在氮磷交互作用下,地上净初级生产力与净硝化速率和净氮矿化速率呈负相关。这与Schleuss等发现植物生物量与净氮矿化速率呈负相关相似,分析原因,土壤可溶性碳(DOC)与地上净初级生产力呈正相关,当土壤碳氮比过高时,土壤微生物对氮的固定增强[30]。
4 结论
单独N添加显著提高了晋北农牧交错带草地土壤无机氮含量及土壤氮转化速率;单独P添加对土壤无机氮含量和氮转化速率均没有产生显著影响;而N和P同时添加虽然显著增加了土壤无机氮含量,但没有显著影响土壤氮转化速率,增加了地上净初级生产力。表明该地区草地土壤受N、P同时限制。未来需要在长的时间尺度研究N和P的互作效应对该类型草地生态系统结构与功能的影响,为农牧交错带草地管理提供科学依据。
猜你喜欢
今日农业(2022年14期)2022-11-10
北京大学学报(自然科学版)(2022年4期)2022-08-18
草业学报(2021年9期)2021-09-22
珠江水运(2017年17期)2017-10-10
农业环境科学学报(2017年2期)2017-03-20
中央民族大学学报(自然科学版)(2015年4期)2015-06-11
新疆大学学报(自然科学版)(中英文)(2014年2期)2014-11-06
水土保持通报(2014年5期)2014-06-09
湖南农业科学(2014年14期)2014-02-27
植物营养与肥料学报(2012年1期)2012-10-26
