
半灌木扩张草原生长季内土壤氮矿化特征及其影响因素
2023-10-08 09:24:00拓行行黄建弟田慧慧李美慧王子临杨梦茹王亦波叶发明
草地学报 2023年9期
拓行行, 黄建弟, 白 玥, 田慧慧, 李美慧, 王子临, 杨梦茹, 王亦波, 叶发明, 李 伟
(1.西北农林科技大学草业与草原学院, 陕西 杨凌 712100; 2.宁夏师范学院化学化工学院, 宁夏 固原 756000; 3.西部机场集团榆林机场有限公司, 陕西 榆林 719000; 4.中国科学院水利部水土保持研究所, 陕西 杨凌 712100; 5.中国科学院大学, 北京 100049; 6.西北农林科技大学水土保持研究所, 陕西 杨凌 712100)
全球变化和人类活动加剧的背景下,干旱半干旱区天然草原常发生灌木/半灌木植物密度、高度及生物量增加,且随时间的推移呈逐渐上升趋势,这种现象被定义为草原灌木扩张或灌丛化(Shrub encroachment)[1,2]。灌丛化可降低植物多样性,影响草地生态产品供给,是生态系统功能下降或荒漠化的表现之一,其已成为不可忽视的全球生态问题之一[3]。但是,灌从化也可使植物群落结构发生变化,影响土壤养分的水平和垂直分布,将土壤养分聚集在其冠层下形成“沃岛效应”[4],从而增加土壤水分下渗,有利于生态系统水分储存和养分的转化[3]。
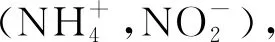
目前关于土壤氮矿化的研究主要集中在氮添加、土壤水分、温度以及微生物活性等方面[12-14];但是不同半灌木扩张强度及生长季内土壤水分、温度变化如何影响土壤氮矿化的机制尚未明晰。宁夏云雾山典型草原作为天然草原中独特的一种类型,其处在一个对自然和人类活动干扰十分敏感的地带,与其他草原类型相比,其状态转化的恢复力阈值很小,且具有常年干燥的气象特征,植被组成更是种类贫乏且结构简单[15,16]。本研究旨在探究生长季内半灌木扩张草原如何通过改变植物群落组成结构以及土壤水温的变化,进而如何影响土壤氮矿化过程。本研究以黄土高原宁夏云雾山不同半灌木扩张强度(对照、轻度、中度和重度)的天然草原为研究对象,深入分析生长季前后期土壤氮矿化过程及其影响因素,揭示了半灌木扩张草原土壤氮矿化的潜在机制,以期为天然草原稳态转变过程理论研究和草原生态保护、恢复和可持续利用提供实践指导。
1 材料和方法
1.1 研究区域概况
本研究在西北农林科技大学宁夏云雾山草原生态系统国家长期科研基地进行,研究区位于宁夏固原市东北部,属黄土高原中偏西部梁状丘陵区(36°13′~36°19′ N;106°24′~106°28′ E),海拔1 800~2 148 m,中温带半干旱气候区,年均气温6.9℃,年均降水量为425 mm。土壤类型以山地灰褐土和黑垆土为主,植被属暖温带典型草原,主要建群种和优势种有本氏针茅(Stipabungeana)、白莲蒿(Artemisiasacrorum)、大针茅(Stipagrandis)等[17]。
1.2 试验设计
根据半灌木扩张强度,选取轻度半灌木扩张(Light subshrub expansion,LSE)、中度半灌木扩张(Moderate subshrub expansion,MSE)、重度半灌木扩张(Heavy subshrub expansion,HSE)和未发生灌木扩张的天然草地作为对照(CK),共4个样地,每个样地的面积为1 ha。在每个样地中随机选择12个10 m×10 m的小区作为重复,每个小区之间至少相距50 m以小区四角用铁桩固定。分别在每个小区内测定土壤氮矿化速率、群落学调查等。

表1 样地土壤特性

表2 样地概况
1.3 试验方法
本研究采用顶盖PVC管法研究植物不同生长期内半灌木扩张对土壤氮矿化的影响,分别测定生长季前期和生长季后期的土壤氮矿化速率,生长季前期的培养时间为2022年5月1日—6月30日,生长季后期的培养时间为2022年7月1日—8月31日。在每个处理样方内砸下直径7 cm、高度12 cm的两个PVC矿化管(露出地面2 cm),每个生长期前用直径3 cm的土钻取两钻土,分别装在自封袋中作为该生长期培养前的土,同时将矿化管的土取出装入自封袋作为前一生长期培养后的土,取出后将PVC管再次砸下用于下一生长期的矿化速率的测定[18]。用AA-3流动分析仪(SEAL Auto Analyzer 3)测定铵态氮和硝态氮浓度。
生长季内土壤温度和土壤含水量由黄土高原生态过程联网监测系统提供(0~10 cm)。同时在每个10 m×10 m的小区内随机选取1个1 m×1 m的样方进行群落学调查,将样方中的所有植物按种分类,本研究用物种丰富度和植物密度作为物种多样性的衡量指标[17]。调查结束后将样方内植物地上部分剪下,清除植物粘附异物后装袋,并带回实验室内在105℃烘箱内杀青,之后在70℃下烘干至恒量,称重记为地上生物量。将样方内的白莲蒿测定其高度、盖度等群落特征,计算白莲蒿的相对高度、相对多度、相对盖度和相对频度,从而计算白莲蒿的重要值。
1.4 数据处理及分析
R硝=(NB硝-NA硝)/t
R铵=(NB铵-NA铵)/t
RN=(NB硝+ NB铵)-(NA硝+ NA铵)/t
式中:NB硝为培养后硝态氮含量(mg·kg-1);NA硝为培养前硝态氮含量(mg·kg-1);NB铵为培养后铵态氮含量(mg·kg-1);NA铵为培养前铵态氮含量(mg·kg-1);t为培养时间(mg·kg-1);R硝为土壤硝化速率(mg·kg-1·d-1);R铵为土壤铵化速率(mg·kg-1·d-1);RN为土壤氮净矿化速率(mg·kg-1·d-1)。文中土壤氮矿化速率统指硝化速率、铵化速率和净矿化速率。
物种丰富度=样方内所有物种的数目(种·m-2)
植物密度=样方内所有物种的个体总数(株·m-2)
重要值=
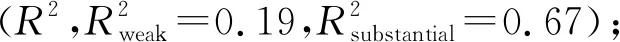
2 结果与分析
2.1 半灌木扩张下草原群落特征的变化
随着半灌木扩张强度增大,草原群落结构发生了显著变化,物种丰富度及植物密度随着半灌木扩张显著下降(图2A、B,P<0.05);在生长季前、后期,地上生物量均随着半灌木扩张的增大而显著增加(图2C,P<0.05)。
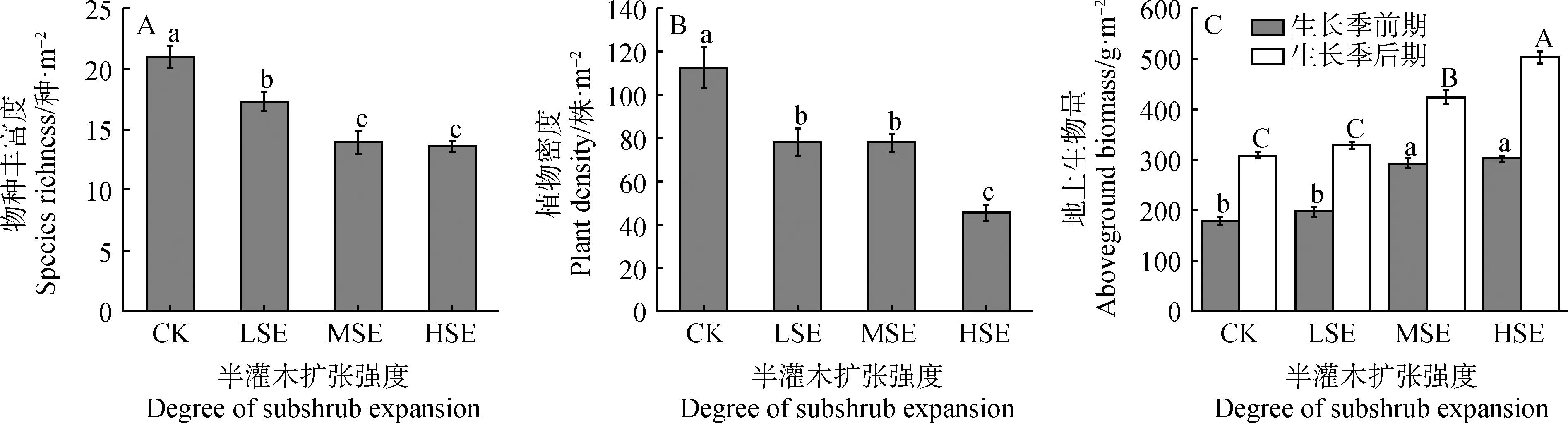
图2 不同扩张强度下草原群落特征的变化
2.2 不同生长期土壤水温的变化
随着半灌木扩张的增强,土壤含水量在生长季前、后期均为显著增加的趋势,并且生长后期土壤含水量显著高于生长季前期(图3A,P<0.05);而土壤温度在生长季前、后期并不随这半灌木扩张的变化而改变(图3B)。
2.3 半灌木扩张对不同生长时期土壤氮矿化速率
生长季前期土壤氮矿化主要表现为矿化形式,主要以土壤硝化为主,且在中度扩张和重度扩张下,土壤硝化速率显著高于对照和轻度扩张;在生长季后期主要表现为氮固持,且对照、轻度扩张和中度扩张下土壤固氮速率显著高于重度扩张草原(图4)。土壤氮矿化速率受半灌木扩张强度和生长期变化的交互影响(表3)。
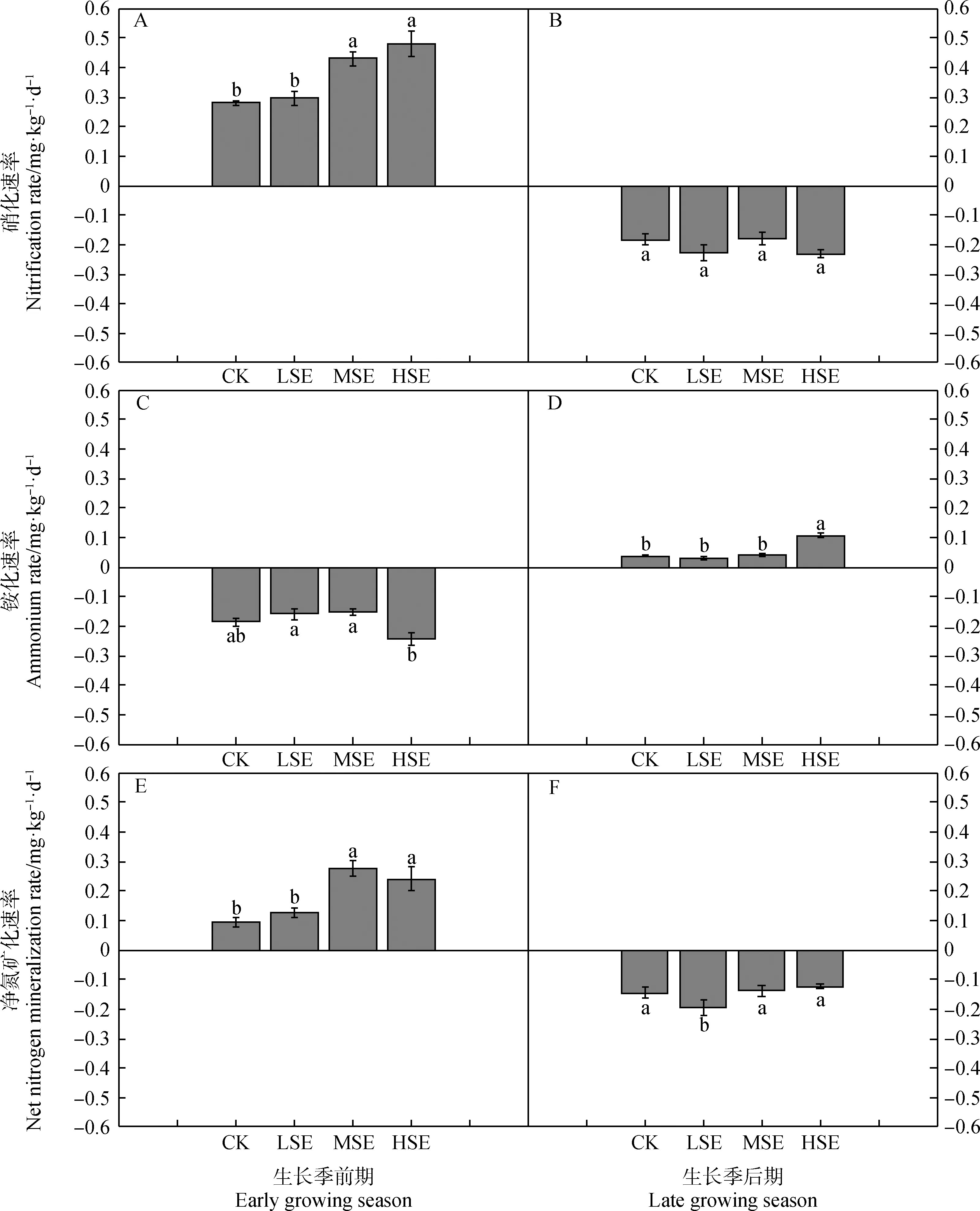
图4 不同扩张强度下土壤硝化、铵化和净氮矿化速率
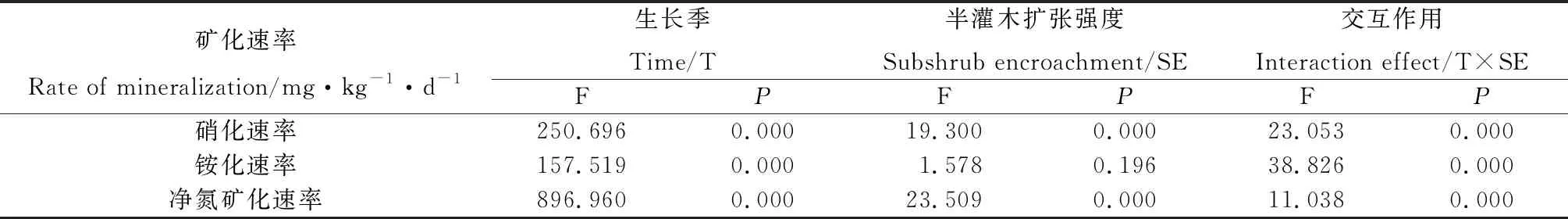
表3 土壤氮矿化速率在不同扩张强度和生长季内双因素方差分析
2.4 物种多样性、土壤水温与土壤矿化的关系
在生长季前期物种丰富度和植物密度与硝化速率和净矿化速率有显著负相关关系(图5 A、C;P<0.05),与铵化速率无显著关系;在生长季后期仅物种丰富度对铵化速率有显著负相关(图5B;P<0.05)。在生长季前期地上生物量与硝化速率和净矿化速率有显著正相关(图6A;P<0.05),生长季后期地上生物量仅与土壤铵化速率有显著正相关关系(图6B;P<0.05)。生长季前期土壤含水量与土壤硝化速率有显著正相关(P<0.05),生长季后期土壤含水量与铵化速率为显著正相关(图7A、B;P<0.05);从整个生长季来看,矿化速率随着土壤含水量的增加呈现下降趋势(P<0.05),随土壤温度的增加呈现先增大后降低的趋势(图7C、F;P<0.05)。
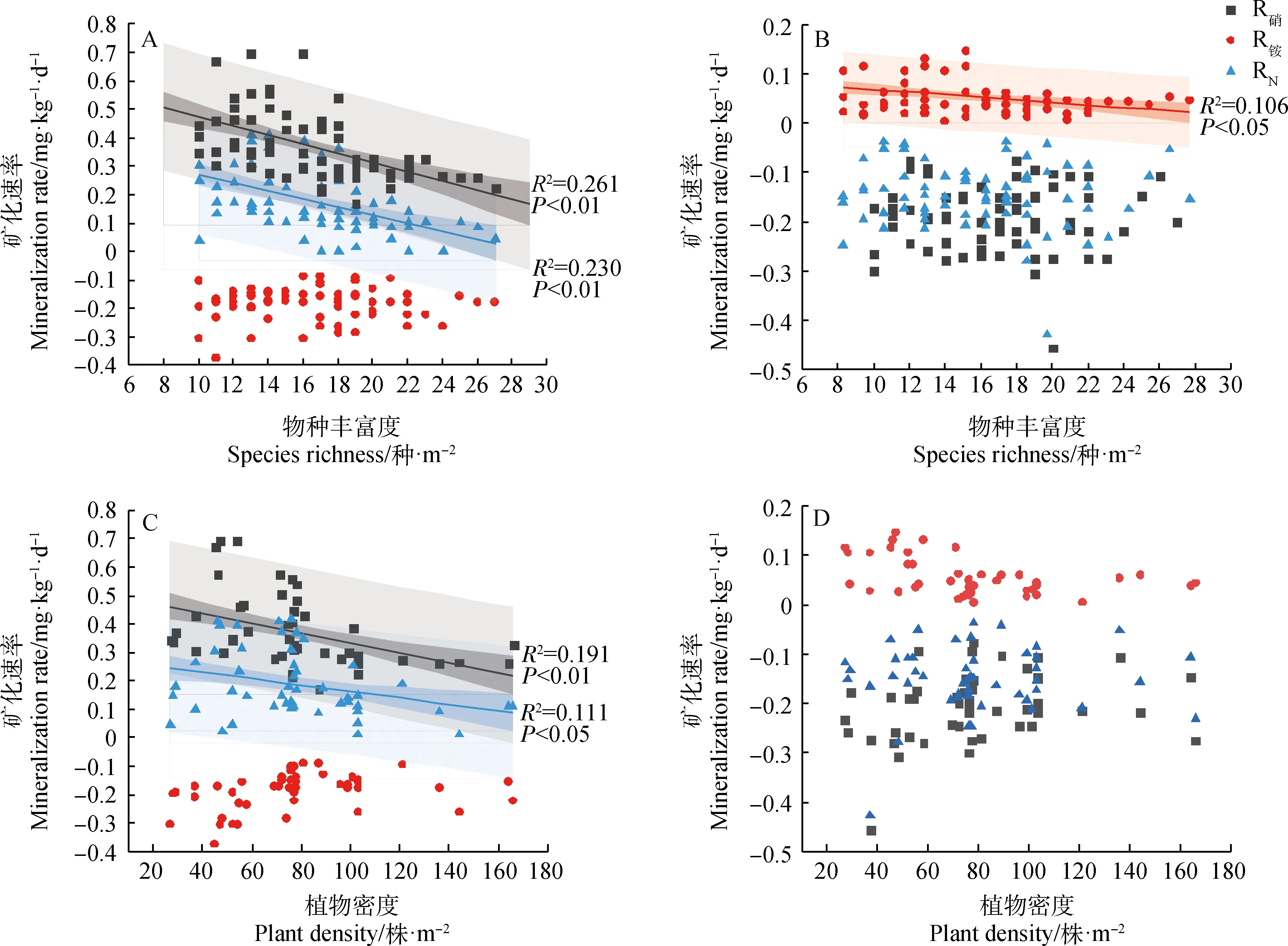
图5 物种多样性与土壤矿化速率的关系
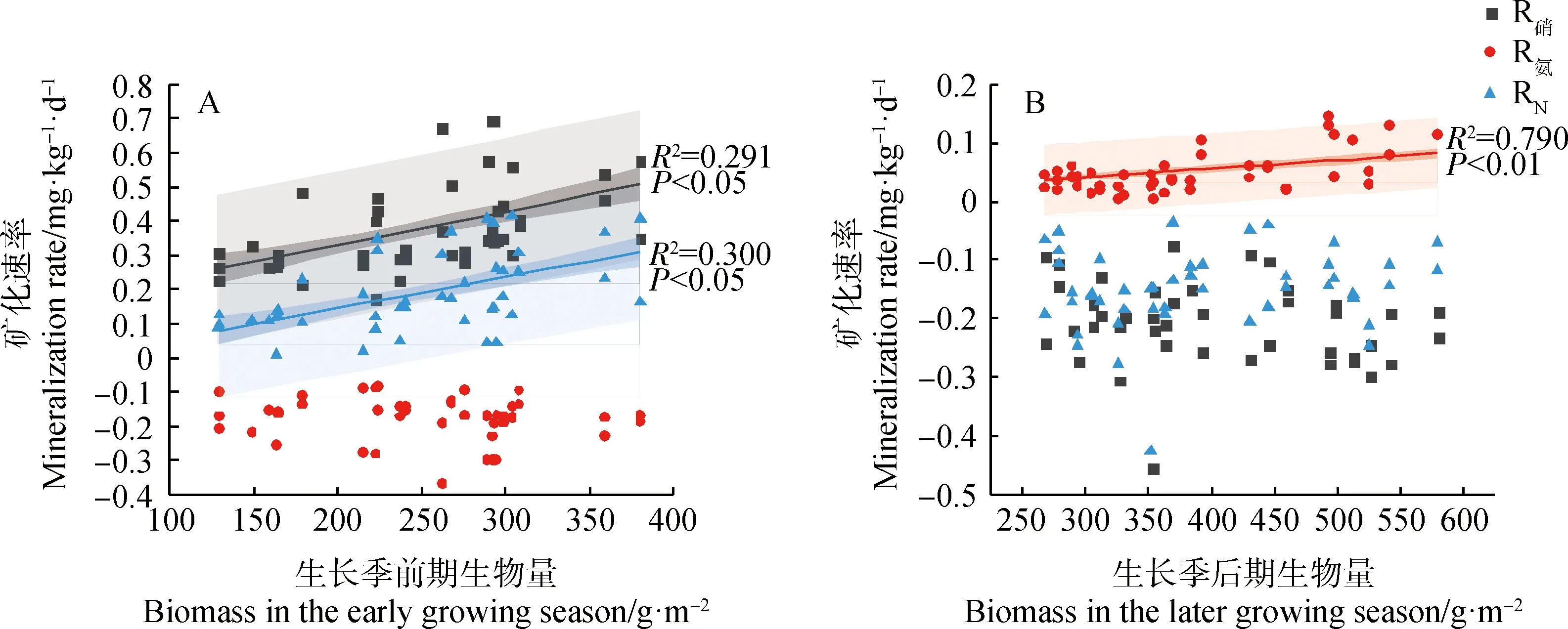
图6 地上生物量与土壤矿化速率的关系
2.5 植被和土壤因子影响土壤氮矿化的结构方程模型
不同生长期和半灌木扩张对土壤氮矿化有着显著交互作用,因此可分析不同生长期内土壤环境及不同扩张强度下群落结构与土壤氮矿化之间的关系。偏最小二乘法结构方程模型(PLS-SEM)表明,半灌木扩张对物种多样性(物种丰富度和植物密度,P<0.05)有显著负影响,对地上生物量有显著正影响(P<0.05);而土壤水分和温度也在不同生长期有显著变化(P<0.05);在整个生长期来看,物种多样性主要对土壤硝化速率和土壤净矿化速率有显著的负影响(P<0.05),地上生物量主要对土壤硝化速率有显著负影响(P<0.05),对土壤铵化速率有显著正影响(P<0.05),土壤水分和温度对硝化速率和净矿化速率有显著负影响(P<0.05),而对铵化速率有显著正影响(P<0.05)(图8)。此外,半灌木扩张通过影响物种多样性和地上生物量间接影响土壤硝化速率和净矿化速率,生长季前、后通过改变土壤含水量和温度间接影响土壤硝化速率、铵化速率和净矿化速率。结构方程模型拟合优度详见表4。
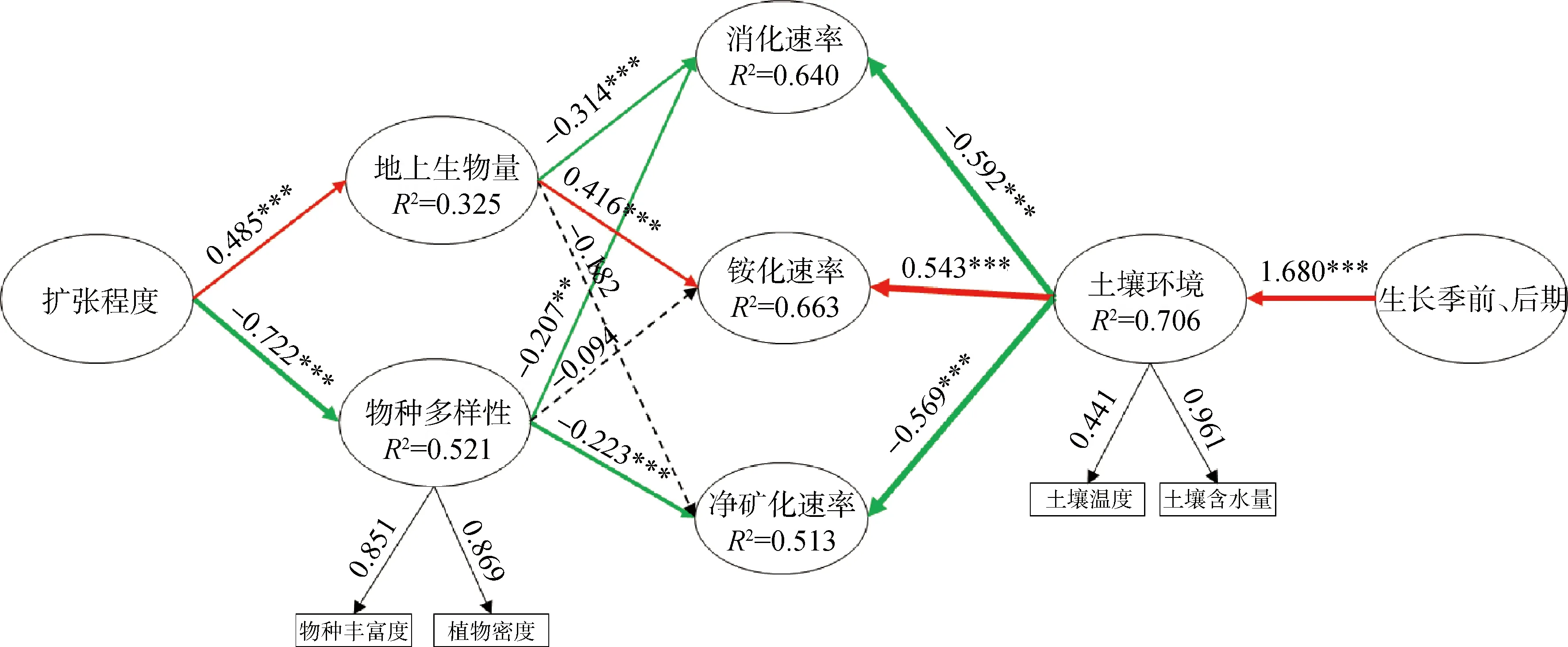
图8 偏最小二乘法结构方程模型(PLS-SEM)分析在不同扩张强度下物种多样性、地上生物量和生长季前后土壤因子对土壤氮矿化速率的影响

表4 结构模型评估
3 讨论
3.1 半灌木扩张草原物种多样性和地上生物量变化及其对土壤氮矿化的影响
半灌木扩张能显著改变草原生态系统结构和功能,影响土壤水分储存、养分转化等生态过程[3]。当草原生态系统优势物种由草本物种转变为半灌木物种后,群落物种组成、植物多样性降低以及生产力会随之增加[21-22]。云雾山典型草原已被半灌木植物白莲蒿入侵,逐渐成为优势物种,形成一种新的稳定状态。物种丰富度和植物密度随扩张增强而显著降低,地上生物量随着扩张增强而显著增大,进而引起土壤氮释放及氮循环过程的改变[8]。此外,群落物种多样性和生产力的变化也是土壤氮矿化改变的重要因素[23,24],在全球草原生境转换和退化过程中,物种多样性持续下降、生态系统生产力对土壤氮素的需求发生转变,从而引起土壤氮转化模式发生改变[25]。受半灌木扩张的影响,云雾山典型草原土壤氮矿化速率发生显著变化,且氮矿化主要以硝化过程为主。这可能是由于草原植物自身的生长对硝态氮的利用强度随时间的推移逐渐增加[26];也可能是由于土壤氮矿化密切相关的功能微生物组成和数量发生改变及其对氮素需求量的变化,从而影响土壤氮矿化速率[27]。物种丰富度变化会引起生态系统土壤氮矿化速率的改变[28]。在本研究中,随着半灌木扩张强度的增大,群落物种多样性显著降低,土壤氮净矿化速率也随着物种多样性的的降低呈现增加的趋势。原因可能是半灌木扩张增大,群落生产力提升,对氮素的可利用性增强所产生的结果,因此,土壤氮素的可利用性对群落物种多样性和生产力的维持同样具有重要作用[29]。
3.2 生长季内土壤含水量和土壤温度的变化对土壤氮矿化的影响
土壤氮矿化季节动态变化主要受到土壤含水量和土壤温度的影响,可直接促进或抑制土壤氮素的转化[30,31]。本研究中,土壤氮矿化速率在生长季前、后期土壤矿化过程发生转变,在生长季前期土壤氮主要表现为矿化,在生长季后期则主要表现为固持。这也可能是生长季内土壤含水量和温度的波动改变了土壤氮素转化模式[32],这种土壤氮素转化模式的改变受到半灌木扩张强度和生长季的双重因素影响。此外,土壤含水量和土壤温度的变化也是影响氮矿化速率的重要因素[33,34],本研究中在生长季前期土壤硝化速率与土壤含水量之间呈显著正相关;在生长季后期,土壤含水量和铵化速率之间有显著正相关。在单独生长期(生长季前期、生长季后期)内土壤硝化速率、铵化速率和净矿化速率和土壤温度之间均无显著相关关系;表明研究区半灌木扩张草原土壤矿化作用对土壤含水量的变化可能更为敏感,这与Wang等[35]的研究结果(土壤净氮矿化速率与温度和水分均为正相关)有所差异。其原因可能是在生长季前后土壤温度变化不显著,从而导致土壤矿化速率不明显[29]。土壤温度和相对干燥度可能改变土壤氮的矿化速率[36],说明土壤矿化速率可能受到土壤水分和温度的共同影响,因此土壤含水量和温度的共同作用对氮素矿化要比单独的温度或水分的影响大;并且在不同生长时期内土壤含水量和土壤温度的变化会使得土壤净氮矿化速率发生显著改变[37,38]。本研究结果表明,不同生长期显著影响土壤硝化速率、铵化速率和净氮矿化速率,线性拟合分析也揭示了土壤含水量和土壤温度显著影响土壤净氮矿化的季节动态,该结果与之前学者在典型草原的研究结果类似[32,39];原因可能是在生长后期大量的降雨增加了土壤氮素淋溶,从而表现出土壤氮矿化受到抑制[39,40]。从整个生长季来看,土壤氮净矿化速率随着土壤含水量的增加呈现显著降低的趋势;而土壤氮净矿化速率随着土壤温度的增加呈现先增加后降低的趋势,其主要原因是土壤氮矿化具有较强的季节性[32,39]。在生长前、后期土壤含水量和土壤温度变化显著,直接导致不同生长期内土壤矿化呈现不同的规律。结构方程模型分析进一步揭示了土壤含水量和土壤温度的季节变化是影响土壤氮的矿化过程的重要因素。未来的研究应结合植物功能性状、土壤微生物等进一步从时间尺度上揭示植物氮利用策略、土壤氮的迁移转化过程以及草地稳态转变(草本态到灌木态)的深层机制,为后续宁夏云雾山天然草地的可持续管理提供理论和实践指导。
4 结论
本研究通过对黄土高原半灌木扩张草原生长季前后土壤氮矿化特征及其影响因素的深入分析,发现云雾山半灌木扩张草原土壤氮矿化主要以硝化作用为主,且具有较强的季节性。半灌木扩张通过减少地上物种多样性和增加生产力,进而改变土壤氮矿化速率;轻度和中度半灌木扩张程度下更有利于土壤氮的矿化(固持)过程。生长季前、后期土壤含水量和温度的变化是导致氮矿化过程发生改变的重要因子,土壤水分和温度增加对氮矿化速率有抑制作用。因此,建议通过人工措施保持草灌的稳定性,促进土壤氮矿化,进而提高土壤氮素的利用性。
猜你喜欢
北京大学学报(自然科学版)(2022年4期)2022-08-18 06:57:34
草业学报(2021年9期)2021-09-22 08:55:16
乡村科技(2018年8期)2018-02-13 12:16:18
辽宁林业科技(2017年4期)2017-06-22 10:23:36
农业环境科学学报(2017年2期)2017-03-20 14:57:30
诗潮(2017年2期)2017-03-16 10:42:30
湖南林业科技(2017年1期)2017-02-06 05:29:17
浙江大学学报(工学版)(2016年2期)2016-06-05 09:20:50
大气科学(2015年5期)2015-12-04 03:04:44
植物营养与肥料学报(2012年1期)2012-10-26 02:49:24
