
血叶兰组培苗与内生真菌共生培养的生长效应研究
2023-09-30 07:35杨泽秀陈英转吴文碟钟云芳陈耀丽
热带林业 2023年2期
杨泽秀,陈英转,吴文碟,钟云芳,陈耀丽,3
1.海南珠峰园林科技有限公司,海南 海口 570220;
2.热带特色林木遗传与种质创新教育部重点实验室/海南大学林学院,海南 海口 570228;
3.福建江南春城市建设集团有限公司,福建 漳州 363005
兰科是高级进化类群,是集药用、观赏和科研作用于一体的植物[1],同时也是十分典型和极具代表性的菌根植物,其菌根的共生真菌是公认人工栽培、引种驯化、资源再生和保护的重要决定性因子[2]。兰花的菌根是兰科植物根系与真菌形成的菌根共生体,菌根的营养来源对宿主植物的生活和营养造成一定的影响[3]。因此,兰花内生真菌和组培苗共生的研究对兰科的保育、移栽炼苗管理及药用药理开发具有重要的意义。
血叶兰(Ludisia discolor)是兰科血叶兰属的多年生草本植物,植株的根茎匍匐生长,茎节形态较为明显,像蚕一样趴在坚硬的石头上,故也被称为“石上藕”。血叶兰干燥的植株有安神健脾、治疗食欲不振等效果。除药用外,还可食用,有益气健身等作用[4-5]。近些年研究发现,菌根真菌在带叶兜兰、铁皮石斛和霍山石斛等兰科植物的研究中均有相关的报道,进一步解释了兰科植物和菌根真菌之间关系的重要意义[6-7]。目前,因栖息地和生境的破坏,野生血叶兰资源正在面临威胁,有些地区甚至面临枯竭,对野生血叶兰资源保存的研究刻不容缓。组织培养、菌根技术及建立菌苗共生体系是实现血叶兰资源保存的良好途径。血叶兰近年的研究主要集中在营养价值、开花物候、内生真菌分离与鉴定和组织培养[8-11],而对血叶兰与内生真菌共生培养的研究少见报道。因此,该研究在无菌血叶兰组培苗上,将经过分离纯化的菌株接种于血叶兰组培苗中,培养3~4 个月,定时记录实验组培苗的生长指标,并与对照组进行比较,筛选菌苗共生的有效培养基,以期建立血叶兰菌苗共生体系,为血叶兰保育、菌根技术的研究提供一定的有效途径。
1 材料与方法
1.1 实验材料
选择大小和长势一致的无根无菌血叶兰组培苗和经分离纯化的内生真菌菌株保存备用。
1.2 实验培养基
内生真菌在PDA 培养基中纯培养,内生真菌和组培苗共生则在改良的DE 培养基中培养。
1.3 内生真菌与血叶兰组培苗的初步筛选
长势一致的无菌苗,洗净苗根部并接种于DE 培养基中,每处理4 株无菌苗,经过1 周培养观察,选择无污染的组培苗和直径为0.5cm×0.5cm 的供试真菌菌块,两者同时接种进行菌苗共生,以不接内生真菌的处理作为对照组,每处理重复10 次。实验过程中细心观察,1 个月后统计实验结果,去除导致苗畸形的内生真菌,保留有益的菌株用于菌苗共生的培养实验。
1.4 内生真菌与血叶兰组培苗共生的筛选
将1.3 保留完好的菌株活化备用,选择长势一致的血叶兰无菌苗,清除根部残留培养基后置于精度为千分之一的电子天平上称重,将处理过无菌苗有序接种于经过改良的DE 培养基上,每瓶接种4 株血叶兰组培苗。
选取内生真菌接种于4 株组培苗的中间部位,每个处理重复10 次,以无菌PDA 块作为对照组。培养3 个月后记录各处理的无菌苗生长发育的效果,实验期间去掉致死或发生畸形的菌株。再培养1 个月后进行数据的记录,每个处理以随机选择10 袋组培苗作为一组,测定血叶兰的生根数量、根系长度、植株高度等植物生长发育指标,测定不同内生真菌实验组和对照组的生长发育指标可作为判断内生真菌对血叶兰组培苗效果的重要理论支撑。
1.5 炼苗培养基筛选
挑选经过菌苗共生,长势一致的健壮组培苗进行炼苗实验,温室瓶内组培苗置放约2d 后拧松瓶盖再放2d,为更好适应外部环境,再将瓶盖完全打开2d。经过消毒的无菌镊子小心取出血叶兰无菌苗,洗净组培苗再用浓度为50%的多菌灵溶液浸泡10min后微晾干种植在已配好的基质中。基质搭配的比例如表2,设置4 种不同基质处理,种植前搅拌均匀,并用水拌透,每个处理3 盘,每盘40 株。经过1 个月的移栽炼苗后,详细记录血叶兰组培苗的存活率以及生长指标的状态。
1.6 培养条件
恒温25℃条件下培养,光照时间为12h(黑暗时间为12h),光照强度控制在1800lx~2300lx 之间。
1.7 数据处理
实验采用Excel 2010 和SPSS 26.0 数据分析软件对该实验记录的数据进行汇总以及差异性分析。
2 结果与分析
2.1 内生真菌对血叶兰无菌苗生长指标的影响
实验在野生血叶兰植株中筛选出的66 株内生真菌中,发现大部分的内生真菌能够导致血叶兰组培苗死亡或致病,与血叶兰不能共生培养(图1)。对剩余的6 株菌株进一步筛选,培养3 个月后发现6株菌株在与血叶兰组培苗共生培养中对组培苗的生长发育有一定的促进作用。共生培养G6 号菌株1 个月后,其内生真菌的菌丝在培养基上匍匐生长,基本无菌丝的缠绕(图2),结果如表1。在组培苗鲜重方面,血叶兰与G9(55.01%)和Y6 菌株(55.13%)共生培养的鲜重增长率与对照组(48.24%)相比有显著差异,即(P<0.05),血叶兰在G6 号菌株中共生培养和血叶兰在对照组中培养的鲜重增长率无显著差异;在血叶兰与G11 号、Y20 号2 个菌株共生培养中发现其鲜重增长率均显著低于实验对照组。在血叶兰株高的测量中发现,血叶兰经过与G6、G9 和G11 菌株进行共生培养的株高显著高于实验对照组;血叶兰与G11 菌株共生培养的茎粗(3.59cm)显著低于对照组(4.62cm),血叶兰与其余的内生真菌共生培养的茎粗测量结果均与实验对照组无显著的差异。在血叶兰根数方面,血叶兰与G6 菌株共生培养促使血叶兰生根效果较为明显,其生根数(4.40 条)与实验对照组(2.20 条)的效果最为突出,血叶兰与Y21 号菌株共生培养的生根数低于对照组。在血叶兰根长方面,血叶兰与G11 (3.94cm) 和Y21 号菌株(3.39cm) 共生培养的根长均显著高于对照组(2.84cm)。通过血叶兰组培苗生长发育指标的考量,发现经过G9 号内生真菌处理的血叶兰组培苗的生长发育效果优于其他处理组;其中G6 号内生真菌属于链格孢属,G9、Y21 内生真菌属于炭疽病属,G11、Y20 内生真菌属于轮层炭菌属,Y6 号内生真菌属于木霉属,说明同属不同种类的内生真菌对血叶兰组培苗的生长发育效果也有所差异。

表1 血叶兰组培苗生长发育指标测定Tab.1 Determination of Growth and Development Index of Tissue Culture Seedlings of Ludisia discolor
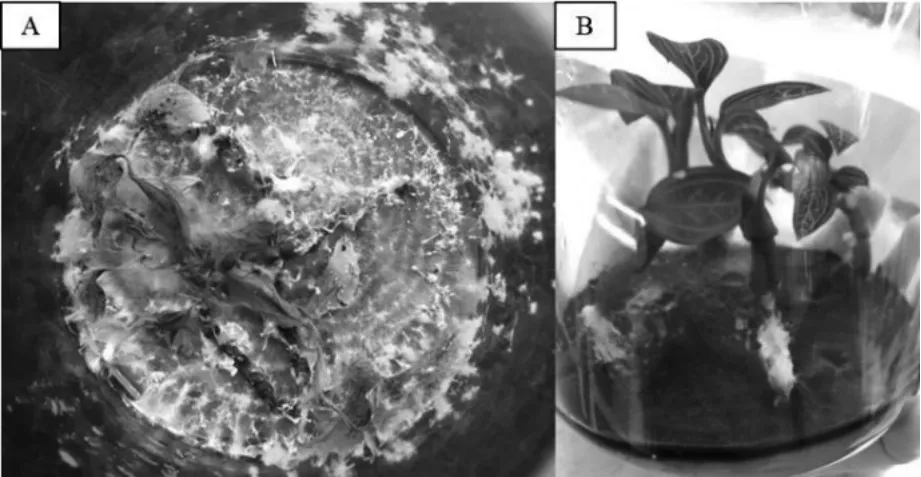
图1 A:共生致死的组培苗,B:正常共生的组培苗Fig.1 A:Ludisia discolor Seedlings by Lethal Symbiotic Culture,B:Normal Symbiotic Tissue Culture Seedlings

图2 血叶兰组培苗生长指标的测定Fig.2 Determination of Growth Index of Tissue Culture Seedling of Ludisia discolor
2.2 基质类型对菌苗共生种植炼苗的影响
植物组织培养炼苗移栽成功的关键在于成活率的占比。该实验将血叶兰移栽种植在4 种不同基质中一个半月后,发现血叶兰组培苗移栽种植在不同基质类型中的成活率均在85%之上(见表2)。其中纯水苔为基质的组培苗成活率为100%,血叶兰无菌苗根系较为粗壮,植株长势良好(图3);血叶兰在松树皮+泥炭土基质(95.5%)中种植,苗生长整齐,生长发育良好。综上,血叶兰以水苔作为种植基质的移栽效果最好。
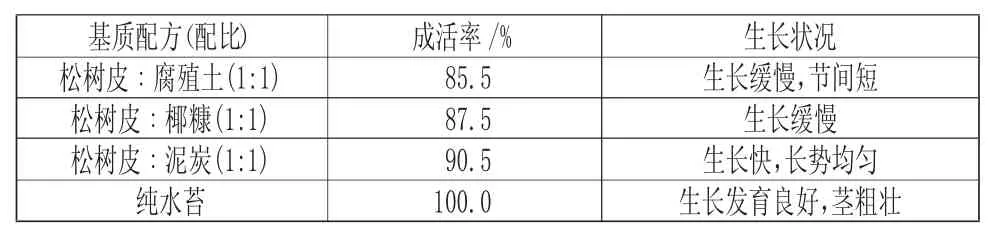
表2 不同基质对血叶兰炼苗成活率的影响Tab.2 Effects of Different Substrates on the Survival Rate of Ludisia discolor Seedling
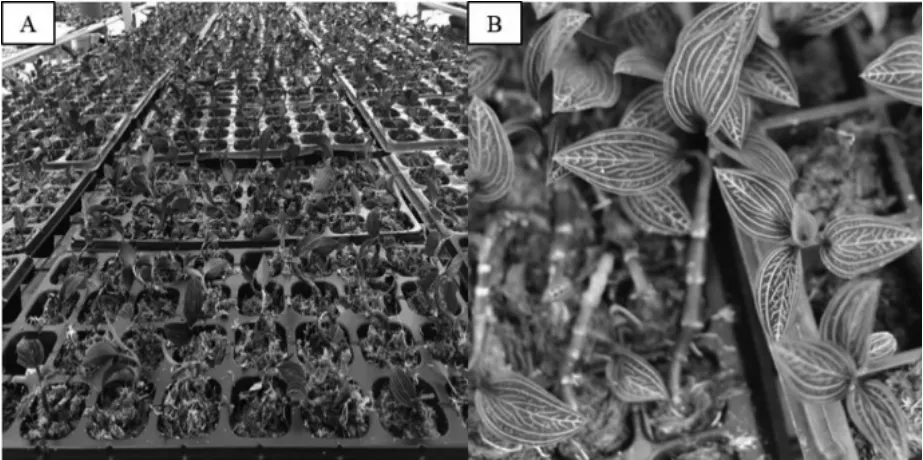
图3 血叶兰的炼苗移栽(以水苔基质为例)Fig.3 Transplantation of Ludisia discolor Seedlings (Sphagnum Substrate as an Example)
3 结论与讨论
3.1 结论
该研究从野生的血叶兰实生苗中分离获得66种内生真菌,有60 株内生真菌与植物存在着极为不利的关系,有6 株的内生真菌均对血叶兰存在有利的关系,对苗具有促进效果。有益菌株分别为G6(链格孢属)、G9 和Y21(炭疽病属)、G11 和Y20(轮层炭菌属)、Y6(木霉属)等6 个菌株。血叶兰与G9 号内生真菌进行共生培养时,其鲜重显著增加(55.01%);经过G6 和G9 菌株共生培养的血叶兰无菌苗的株高、茎粗和生根数等生长发育指标均有显著促进效果;经过G11 号菌株共生培养的血叶兰无菌苗的根长有促进效果。综上所述,血叶兰与G9 号内生真菌进行共生培养的生长发育指标最好。血叶兰以纯水苔作为种植基质为宜,成活率达100%,植株茁壮成长。
3.2 讨论
3.2.1 不同栽植基质对内生真菌与血叶兰组培苗共生培养的影响
血叶兰无菌苗移栽的成活与否跟种植的基质紧密相关。植物需要在组培室内异养条件或者半自养条件转化为全自养条件,其湿度、光照强度和温度对组培苗的成活起关键作用,基质类型也不例外[12]。血叶兰在基质为V火山石:V泥炭土=3:1 中的成活率高达91.7%,在V泥炭土:V松树皮:V珍珠岩=3:1:1 的种植基质中,血叶兰组培苗的成活率达87.0%[13-14]。该实验将水苔作为血叶兰移栽炼苗的种植基质,经过有益菌株共生培养的血叶兰的成活率高达100%,几乎全部成活,植株健壮且生长发育状态良好。原因可能是炼苗基地有遮阴网,空气流畅,再加上水苔自身的保湿效果,使血叶兰茁壮成长。该实验以保水保肥能力强的纯水苔作为种植基质,植株成活率高,这一研究结果与兰科植物中的铁皮石斛和泼墨石斛的移栽炼苗研究结果基本一致[15-16]。因此,血叶兰的后期炼苗,采用水苔作为基质为宜。
3.2.2 血叶兰共生真菌的筛选
内生真菌资源在兰科植物中具有多样性,植物中分离出来的菌株也有异同。不同的内生真菌在植物生长的各阶段发挥的作用也不同,这与植物所处的环境和必需的营养物质等因素有关联[1]。纵使兰科内生真菌资源丰富,并非全部的内生真菌都能够促进兰科植物的生长,少量的内生真菌对兰科植物组培苗具有致病性,出现这种现象的原因可能与接种内生真菌时的数量、菌苗共生的培养基和植物生长发育各阶段均有关系[17-18],植物所需营养超值和内生真菌菌丝的生长过于旺盛都会促使内生真菌和组培苗共生的关系难以达到和谐[19]。五唇兰、海南钻喙兰、大花蕙兰和铁皮石斛等兰科植物的菌苗共生培养都是选择DE 的培养基作为内生真菌和组培苗共生的培养基[20-22]。
以改良的DE 培养基为实验培养基,将从野生血叶兰中分离出来的66 株内生真菌与血叶兰无菌苗进行菌苗共生培养3 个月。结果显示,在所有内生真菌中仅有6 株内生真菌在菌苗共生培养实验中发挥一定的促进作用,占总资源比率的11.00%;有大量的内生真菌资源都会限制血叶兰无菌苗的生长发育,使其生长状态不佳,植株致死率高达89.00%。该实验初筛的有益内生真菌6 株,其对血叶兰组培苗的促生作用均有所不同。血叶兰无菌苗的生根效果在G11 号内生真菌共生培养中较为明显,组培苗根系最长;血叶兰组培苗的鲜重在G9 号内生真菌共生培养中有所增加(55.01%),其株高(9.14cm)、茎粗(4.7mm)、根数(3.60 条)等生长发育指标效果均显著优于其他菌株,原因可能是内生真菌种类的多样对血叶兰无菌苗的促生机制也多样。内生真菌和金线莲组培苗共生实验中发现有5 株内生真菌对金线莲组培苗的生长发育具有促进作用,其中3 株内生真菌分别为镰刀菌属、弯劲霉属和曲霉属[23];而镰刀菌属、刺盘孢属和链格孢属等多是致病的内生真菌。研究发现从野生兰科植物中获得的镰刀菌对同科植物铁皮石斛的种子萌发具有促进效果[24],说明并不是所有的镰刀菌都会促使植物发病。镰刀菌有致病镰刀菌和非致病镰刀菌之分[25],而镰刀菌属具有一定刺激兰科植物种子萌发的作用,也能控制作物的枯萎病[26-27]。综上,说明兰科植物的生长与镰刀菌之间有着密切的关系。内生真菌的有利条件与植物宿主之间存在着协同的作用。植物内生真菌的丰富资源、维持机制和真菌与生态功能之间的关系需进一步探索。
猜你喜欢
——扁根带叶兰
福建农林大学学报(自然科学版)(2021年3期)2021-05-26
辽河(2021年2期)2021-03-24
婚姻与家庭·婚姻情感版(2019年5期)2019-06-03
畜牧兽医科技信息(2018年8期)2018-02-13
创新作文(小学版)(2016年23期)2016-12-01
现代畜牧科技(2016年5期)2016-10-21
河池学院学报(2015年2期)2015-02-27
中国药业(2014年21期)2014-05-26
山东畜牧兽医(2012年1期)2012-04-13
河北林业科技(2011年5期)2011-07-10
