
大藤峡大坝蓄水对浔江鱼类群落的影响
2023-09-28 03:41李育森赵会宏王大鹏罗永巨黄世慧余梵冬
生态学报 2023年17期
李育森,赵会宏,王 庆,王大鹏,罗永巨,周 磊,黄世慧,余梵冬
1 华南农业大学海洋学院, 广州 510000 2 广西水产科学研究院, 广西遗传育种与健康养殖重点实验室,南宁 530021 3 中国水产科学研究院珠江水产研究所,广州 510380
鱼类作为水生生态系统中的重要类群,可以通过下行效应影响整个水生态系统的生物与环境,同时鱼类群落也会受到外界自然环境的影响[1]。基于鱼类群落对环境变化的响应也通常被用于生态系统完整性评价工作[2—4]。影响鱼类群落结构与资源量的因素可分为环境因素与人为因素,环境因素又可以归类为生物因素与非生物因素两大类。唐衍力等[5]发现,不同年龄的许氏平鮋由于饵料分布的差异,以及存在着摄食偏好,会分布于不同的环境以减少种内竞争;何雄波[6]研究表明,不同鱼类可通过改变对相同食物的摄食比例来缓解种间竞争。鱼类有特定的水文需求,水温会影响鱼类的生活史,包括其分布、洄游、产卵和生长等[7]。在浊度较高的水域饵料生物的丰度相对更高,鱼类饵料来源丰富,更有利于鱼类种群的增长[8]。部分研究发现,pH与溶解氧是驱动鱼类群落结构变化的主要环境因子[9—11];也有研究发现盐度与底质也会影响鱼类群落结构[12—13]。
人为因素对鱼类群落的影响主要包括水利水电工程建设、过度捕捞、引进外来物种等[14—16]。近年来越来越多的水利水电工程不仅会对原有的鱼类群落产生影响,还会导致栖息地的退化,从而降低水域生态系统的生物多样性[17]。水利水电工程的运行改变了河流原本的水文动力、水质状况、初级生产力等进而影响鱼类群落[18]。大坝蓄水导致水体流速减缓,大量泥沙、营养物质在库区滞留,水体垂直交换受到限制,水温与溶氧在垂直方向上分层[19]。有研究表明大坝降低了水体的搬运作用与水动力,造成水中携带的泥沙与营养盐堆积,虽然降低了水体的浊度与硬度等,但有毒有害物质也随之沉降,从而影响水质[20—21]。河流的连通性是保证鱼类洄游的前提,拦河筑坝有可能会阻碍洄游性鱼类的索饵、生殖等洄游路线。葛洲坝截流后,中华鲟数量锐减,洄游到葛洲坝下产卵场的亲鱼减少,自然产卵年际间由连续变成偶发[22];坝下江段白鲟与长江鲟的捕获数量也急剧减少[23]。除了直接阻断亲鱼洄游路线外,鱼类繁殖还会受到大坝泄洪或者蓄水造成的水流速、水温等变化的影响。李伟等[24]对沅水干流四大家鱼的调查中发现,大坝阻隔导致受精卵所需最小漂流距离不足从而影响了四大家鱼的繁殖,同时水库下泄水温度偏低,鱼类繁殖也会滞后。徐薇等[25]对银盘电站蓄水前后乌江下游繁殖鱼类调查发现,流水生境繁殖类群减少、缓静水生境繁殖类群增加,部分鱼类产卵规模与产卵场减小,漂流性卵鱼类早期资源衰退。Rowell等[26]发现科罗拉河大坝的修建使得当地麦氏托头石首鱼幼鱼生长速率下降、性成熟年龄延缓。广西大藤峡枢纽蓄水后,其坝下的浔江水文状况变化较大,极有可能对浔江鱼类群落结构造成影响。本研究于浔江蓄水前后共计4年对其鱼类资源进行调查,探究鱼类群落受到的影响,为后续浔江的生态保护工作提供理论基础。
1 研究区域概况
浔江位于广西壮族自治区东部,属于珠江流域的西江干流中游,自北向南的黔江与自西向东的郁江在桂平汇合称为浔江,盛行亚热带季风气候。近年来关于浔江生态环境的研究报道越来越多[27],陈晓鸿等[28]研究表明,浔江已属于Ⅴ类水质,水体总氮、总磷超标,鱼体内锌、铬、砷同样存在超标现象,同时,张迎秋等[29]研究发现,长洲水利枢纽改变了浔江桂平段的水文状况,导致该江段鱼类优势种发生变化。广西大藤峡枢纽于2014年11月工程开工,于2019年6月蓄水,由黔江主坝与副坝、南木江副坝组成,总库容34.79亿立方米,防洪限制水位47.60 m,防洪库容15亿立方米。根据全国大江大河实时水情记录(http://xxfb.mwr.cn/sq_djdh.html),大坝蓄水前(2017年7月至2019年2月),浔江大湟江口处流量为5963.64 m3/s,蓄水后(2019年7月至2021年2月)流量有所减少,为5709.63 m3/s;蓄水前平均水位为24.87 m,蓄水后为22.78 m,水位有所降低。
2 材料与方法
2.1 研究区域及样品采集
调查区域为广西壮族自治区大藤峡枢纽坝下的浔江段(桂平至平南,图1),本研究在大藤峡枢纽下游按从近到远设置了5个采样点,S1—S5距离大藤峡枢纽逐渐增大。每个采样点布置定制刺网2张(长度50 m,高1.2 m,网目尺寸3.5—6 cm),虾笼2个(长15 m、高20 cm,宽25 cm),钓钩100个。作业时间为19时至次日7时。采集到的鱼类现场鉴定,并测量和体长(精确到mm)、体重(精确到g)、尾数等数据,随后放归浔江,减少对物种资源的破坏。对于未能及时鉴定的物种则冷藏运送至实验室进一步鉴定并测量。

图1 采样区域Fig.1 Sampling areaS1—S5为距离大藤峡枢纽越来越远的5个采样点
调查时间为2017年7月至2021年2月,调查频率为每月一次。7、8月为夏季,9—11月为秋季,12月至次年2月为冬季,因此,2017.07—2018.02,2018.07—2019.02数据分别归为2017、2018年讨论,后面的采样年度以此类推。大藤峡枢纽蓄水时间为2019年6月,所以2017年、2018年表示蓄水前的情况,2019年、2020年代表蓄水后情况。
2.2 鱼类丰度生物量比较(ABC)曲线与相似性聚类
鱼类ABC曲线最初由Warwick[30]提出,并已广泛应用于评估鱼类群落所受到的干扰水平[31]。从理论上讲,ABC曲线结合了物种丰度(数量)优势和生物量(质量)优势,基于r—选择和K—选择的经典进化理论,两种曲线可以产生3种可能的相对位置,分别代表着受到轻度干扰,中度干扰和强烈干扰的生物群落[30]。鱼类ABC曲线采用PRIMER 5.0 软件绘制;此外,采用相似性分析(SIMPER)进行组间相似性及主要贡献种的筛选[32]。采用非度量多维尺度分析(NMDS)对不同年份各采样点鱼类组成差异进行描述。
2.3 生态网络模型
鱼类营养级数据取自世界鱼类数据库(https://www.fishbase.se/),以鱼类为节点,摄食关系为线,通过营养级关系构建邻接矩阵,最后将矩阵输入R语言转化为生态网络图。不考虑鱼类同类相食、寄生与腐蚀等关系,假设两种鱼类营养级差值大于等于1时,高营养级个体捕食低营养级个体[33]。
采用点度分布[p(k)][34]、网络密度(D)[35]、平均路径长度(APL)[36]和聚类系数(C)[37]来评价生态网络结构的稳健性。采用累积概率分布[P(k)]来替代p(k)以便减少误差[38]。同时采用K-S检验网络的点度分布是否符合幂律分布[P(k)—k-α][39]。P>0.05则点度分布符合幂律分布,此时的网络存在集散节点,属于复杂网络。当α>3时,网络中基本不存在离散节点;当3≥α> 2时,网络有少量集散节点,为无标度网络;2≥α>1时,拥有较多的集散节点。
(1)
(2)
式中,nk为点度值为k的数量,N为种类数,k和k′均表示某个点的点度。
网络密度(D)公式如下:
(3)
式中,d(i,j)为种类i与j之间的关系量,N为种类数。
平均路径长度(APL):
(4)
式中,d(i,j)为种类i与j间的最短路径,N为种类数。
聚类系数(C):
(5)
式中,N为种类数,Ei为种类i的k个邻节点之间实际存在的边数量,ki为种类i邻节点的数量。
生态网络图与相关参数的计算均通过R语言完成,涉及igraph(https://igraph.org/)、reader(https://cran.r-project.org/web/packages/readr)、haven(https://cran.r-project.org/web/packages/haven/)、gglot2[40]安装包。
2.4 相对重要性指数与时间生态位
相对重要性指数[41]:
IRI=(N+W)F×104
(6)
式中,N为某一种类的数量占总尾数的百分比;W为某一种类的重量占总重量的百分比;F为某一种类的出现站数占调查总位点数的百分比。当IRI值大于500时为优势种,500—100的为常见种,100—10的为一般种,10—1的为少见种,小于1的为稀有种。本研究将IRI>10的鱼类作为主要鱼类进行研究。
时间生态位宽度采用 Shannon-Wiener指数计算[42]:
(7)
式中,Bi为生态位宽度,Pij为种i在第j个站位下的个体数占该种所有个体数的比例;R为资源总数。Bi<1.00为窄生态位种,1.00≤Bi≤2.00为中生态位种,Bi>2.00为广生态位种。
空间生态位重叠采用 Pinaka 指数计算[43]:
(8)
式中,Qik为种i与种k的生态位重叠指数,其值越大表示生态位重叠程度最高。其取值范围为0—1;Pij为种i在第j个资源状态下的个体数占该种所有个体数的比例;Pkj为种k在第j个资源状态下的个体数占该种所有个体数的比例;R为资源状态总数。其中,Qik>0.6时,生态位显著重叠[44]。
2.5 总体相关性检验
用Pianka生态重叠指数评估群落构建模式。
(9)
(10)

(11)
式中,Iobs为群落C-score指数实测值,Isim为零模型群落值,Ssim为零模型群落标准离差。当标准化效应尺度值(SES)介于-2和2之间时, 说明群落处于随机分布格局(P>0.05); 当标准化效应尺度值绝对值大于2时(P<0.05),说明群落为非随机分布, 正值说明群落处于分离格局, 而负值则说明群落处于集聚格局[45]。
2.6 相关数据的差异性分析
使用SPSS 19.0软件与GraphPad Prism 9.0.0软件进行数据统计,利用单因素方差分析(ANOVA)对近4年渔获物种类、数量、质量的季节、年度差异进行评价,差异显著性阈值P=0.05。
3 结果与分析
3.1 鱼类群落物种组成和相似性
浔江2017—2020年调查期间共采集到鱼类77种,隶属7目19科63属。渔获重量与尾数如表1所示。单因素方差分析结果显示,蓄水前各季节种类数在21—55种,日均渔获质量(8.20±3.46)kg至(12.03±2.78)kg,日均渔获尾数在(89±42)尾至(326±127)尾之间;蓄水后各季节种类数在17—45种,日均渔获质量(7.54±3.19)kg至(11.01±2.46) kg,日均渔获尾数在(105±74)尾至(229±125)尾。蓄水前浔江鱼类种类数量和日均渔获重量优于蓄水后,但未达到统计学意义上的显著差异;而日均渔获尾数在蓄水后有所上升。

表1 浔江近4年渔获物详情Table 1 Details of catch in Xunjiang river in recent 4 years
采用非度量多维尺度分析(NMDS)对不同年份各采样点鱼类组成差异进行描述,结果显示2017年与2018年各采样点之间相隔较远,而2019年与2020年各采样点之间都相对聚集,表明蓄水后各采样点间鱼类组成差异有所减小(图2)。

图2 浔江近4年鱼类群落结构的NMDS分析 Fig.2 NMDS analysis for fish assemblages structure in Xunjiang river in recent 4 yearsNMDS:非度量多维尺度分析Non-metric multi-dimensional scaling
各采样点鱼类种类差异如图3所示:2017年与2018年鱼类种类数量呈现出上游采样点多于下游采样点的趋势;2019年与2020年有所不同,下游采样点种类数高于上游采样点。于此同时,优势种卷口鱼(Ptychidiojordani)、赤眼鳟(Squaliobarbuscurriculus)、鲤(Cyprinuscarpio)和粗唇鮠(Leiocassiscrassilabris)中,除了鲤随着时间推移在5个站点间始终分布较为均匀外,其他优势种在各站点间均由原来的分布不均变得更加均匀(表2)。

表2 不同年份各站点鱼类丰度变化/%Table 2 Changes in fish abundance at each station in different years

图3 近4年浔江各采样点鱼类种类差异Fig.3 Fish species difference between different stations of Xunjiang river in recent 4 years
基于渔获种类与重量的相似性聚类分析显示,各年度间鱼类群落相似度均在59.73%以上,其中2018年与2019年鱼类群落相似性最高,达75.42%,2017年与2020年鱼类群落相似性最低,仅为59.73%(图4),对相似性贡献最高的种类为卷口鱼、其次为赤眼鳟。

图4 浔江鱼类群落相似性聚类Fig.4 Similarity clustering of fish communities in Xunjiang river卷口鱼 Ptychidio jordani;赤眼鳟 Squaliobarbus curriculus;鲤 Cyprinus carpio;粗唇鮠 Leiocassis crassilabris;斑鱯 Mystus guttatus;鲮Cirrhinus molitorella;罗非鱼sp Tilapia;瓦氏黄颡鱼 Pelteobagrus vachelli;鲇 Silurus asotus;麦瑞加拉鲮Cirrhinus mrigala;草鱼Ctenopharyngodon idellus;大刺鳅Mastacembelus armatus;海南鮊Culter recurviceps;间鱼骨Hemibarbus medius;尖头塘鳢 Eleotris oxycephala;大眼鳜Siniperca kneri;鲫Carassius auratus;银鯝Xenocypris argentea;东方墨头鱼Garra orientalis;短须鱊 Acheilognathus barbatulus
3.2 鱼类群落ABC曲线
浔江近4年鱼类ABC曲线如图5所示:大藤峡大坝蓄水前后,曲线发生了明显的变化,W值在 -0.004到0.092之间。累积生物量曲线起点呈连年上升趋势,部分鱼类优势度有所上升。蓄水前生物量曲线位于丰度曲线上方,鱼类群落内生物量比丰度更显优势,鱼类群落受到外界干扰较小,尤其是2018年鱼类群落最为稳定,此前W值均大于0;蓄水后生物量曲线与丰度曲线交叉重叠现象更为严重,丰度与生物量曲线分布优势难分,鱼类群落受到的干扰较为严重,其中2020年曲线W值为-0.004,受到的干扰加剧,干扰程度在大坝蓄水后随着时间的推移而加剧。
近日,浩鲸科技携手南京市公安局交通管理局打造的智慧交通综合应用平台二期项目正式通过初验。该平台整合一期项目数据和“一标三实”信息采集会战数据,可广泛用于开展路况拥堵态势分析、道路拥堵分析及大队工作监管等,助力南京市公安局交通管理局推进以大数据研判为支撑,信息主导警务的智慧新交管建设,帮助交警在面对日益复杂的道路状况和出行场景时,有效提升交通管理效能。

图5 浔江鱼类ABC曲线Fig.5 Fish ABC curve of Xunjiang riverABC:鱼类丰度-生物量比较曲线Abundance biomass comparison curve
3.3 群落生态网络分析
3.3.1鱼类营养级
参与生态网络构建的鱼类共有77种,其中2017年53种,2018年55种,2019年45种,2020年34种。营养级范围2—4.4。顶级物种(仅为捕食者,营养级>3.4)有红鳍原鲌(Cultrichthyserythropterus)、鲶(Silurusasotus)、淡水石斑鱼、长吻鮠(Leiocassislongirostris)等序号范围在1—20的种类,中间物种(同时作为捕食者与被捕食者,3≤营养级≤3.4)有中间黄颡鱼(Pelteobagrusintermedius)、粗唇鮠、七丝鲚(Coiliagrayii)等序号范围在21—58的种类,带半刺光唇鱼(Acrossocheilushemispinuscinctus)、麦瑞加拉鲮(Cirrhinusmrigala)、鲮(Cirrhinusmolitorella)等序号范围59—77的为基础物种(仅被捕食,营养级<3)(表3)。

表3 浔江鱼类营养级Table 3 Trophic levels of fishes at Xujiang
3.3.2生态网络特性
浔江鱼类生态特性如表4、图6所示:2017年与2018年网络密度水平相当,均为0.11,2019年网络密度明显降低(0.09),2020年网络密度又增至0.13。平均路径长度均为1。幂指数呈现递增趋势,表示网络中的集散节点数在减少,2017—2019年3≥α>2,且p>0.05,生态网络中有仅少量集散节点,为无标度的复杂网络结构,2020年α>3,此时生态网络中基本不存在集散节点,为简单网络,于此同时聚类系数为0。
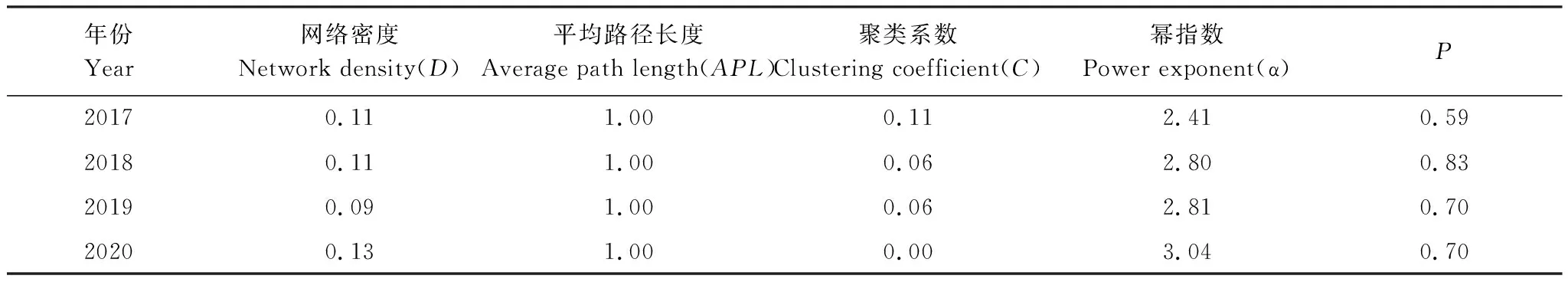
表4 浔江鱼类群落生态网络特性Table 4 Ecological network characteristics of fish community in Xunjiang River

图6 2017—2020年浔江鱼类生态网络Fig.6 Fish ecological network of Xujiang river form 2017 to 2020各序号对应的鱼类种类见表3
3.4 部分经济鱼类时间生态位
3.4.1时间生态位宽度
调查期间,卷口鱼、赤眼鳟、鲤、粗唇鮠为优势种(IRI>500),IRI分别为2347.23、1317.83、1029.10、850.59;罗非鱼sp(Tilapia)、斑鱯(Mystusguttatus)等7种鱼类为常见种,IRI值在104.47—451.23之间;鲶、海南鲌(Culterrecurviceps)、大眼鳜(Sinipercakneri)等9种鱼类为一般种。本文仅讨论IRI>10的经济鱼类,大藤峡蓄水前后,20种主要鱼类时间生态位宽度在0.81—2.47之间,蓄水前时间生态位最宽的是鲤(2.47),其次是赤眼鳟(2.46),最窄的是纹唇鱼(Osteochilussalsburyi)(0.73)。蓄水后时间生态位最宽的是卷口鱼(2.42),其次是鲤(3.35),最窄是鲶(0.81)。蓄水前后始终为广生态位种(Bi>2.00)的有卷口鱼、赤眼鳟、鲤等;始终为中生态位种(1.00≤Bi≤2.00)的有罗非鱼sp、大眼鳜、鲫等;此外,以东方墨头鱼为首的9种鱼类生态位宽度变宽;鲶为首的11种鱼类生态位宽度变窄。大藤峡枢纽蓄水后,草鱼由广生态位种变为中生态位种;鲶由中生态位种变为窄生态位种;大刺鳅与海南鲌由中生态位种变为广生态位种;东方墨头鱼与纹唇鱼由窄生态位种变为中生态位种(表5)。

表5 主要鱼类IRI值及时间生态位宽度变化Table 5 IRI value and temporal niche width of major fishes
3.4.2时间生态位重叠
蓄水前后浔江主要鱼类时间生态位重叠值变化如表6所示:共获得主要鱼类种对有380对,时间生态位重叠值(Qik)范围为0—0.92。Qik>0.6,即显著重叠的种对有83对,占总对数的21.84%; 0.6≥Qik≥ 0.3的种对有170对,占总对数的44.74%;0.3 >Qik的种对有127对,占总对数的33.42%;瓦氏黄颡鱼-七丝鲚(sp8-sp20)的生态位重叠值最高(0.92),其次是鲶-鲫(sp12—sp15)、间鱼骨-七丝鲚(sp9—sp20)重叠值分别为0.91、0.90,均出现在蓄水后。而蓄水前生态位重叠最高的种对是鲤-草鱼(sp3—sp10),为0.85;其次是赤眼鳟-大刺鳅(sp2—sp11)、间鱼骨-草鱼(sp9—sp10),均为0.82,这三个种对在蓄水后除了赤眼鳟-大刺鳅仍为显著重叠外,另外两个种对重叠值均大幅度降低。蓄水前后,显著重叠的种对有所下降,由46对降为37对;0.6≥Qik≥0.3的种对有所上升,76对上升为94对。

表6 主要鱼类时间生态位重叠Table 6 Temporal niches overlap of major fishes
3.5 浔江鱼类群落格局与构建模式
基于浔江鱼类时间生态位重叠构建的零模型结果显示:2017—2020年,Pianka指数实测平均值均大于模拟平均值,表明近4年,浔江鱼类群落的组成取决于环境因素,生境过滤占主导作用。标准化效应尺度均大于2,说明该群落为非随机分布,处于分离格局。Pianka指数和资源利用模式的观测值与模拟值之间具有显著差异(p<0.05),也表明浔江鱼类群落的构建是非随机占主导。但2020年标准化效应尺度降至2.70,实测值更接近模拟值,表明随着时间的推移,浔江鱼类群落趋向于由竞争主导的随机分布发展格局,栖息地的异质性有所降低(表7)。

表7 基于Pianka指数的标准化效应尺度分布Table 7 Distribution of standardized effect scale base on Pianka index
4 讨论
4.1 大藤峡蓄水对浔江鱼类群落稳定性的影响
江河的自然流态通常被认为是水生生物多样性与生态完整性的要素之一[46],大藤峡枢纽蓄水降低了河流连通性,改变了河流原有的水文状况,这也导致了鱼类群落发生了变化。比如,大坝蓄水后浔江大湟江口处平均流量由5963.64 m3/s,减少为5709.63 m3/s;平均水位由24.87 m显著降低为22.78 m。大坝蓄水后,浔江两年度监测到的鱼类种类由蓄水前的69种降为51种,物种数量有所下降,这与前人研究结果相似[17]。大藤峡枢纽蓄水前,鱼类ABC曲线揭示浔江鱼类群落呈现出好转的迹象,仅受到轻微的外界干扰,并且干扰主要来源于人为捕捞。究其原因,可能是禁渔期制度的实施以及人工增殖放流和内陆捕捞准用渔具类型及渔具最小网目(或网囊)尺寸限制对鱼类资源的恢复产生了一定的效果。有研究表明不同类型刺网渔获物结果差异很大,网目尺寸越小对渔获群落干扰越严重[47]。大藤峡枢纽蓄水后,2019年ABC曲线显示,生物量曲线与丰度曲线开始相交,并且到2020年,丰度曲线开始占优,W值已经小于0,表明此时鱼类群落受到较为严重干扰[30,32]。此外,调查发现蓄水前浔江鱼类物种数量和日均渔获重量优于蓄水后,且蓄水后鱼类群落以小规格鱼类为主,这也表明了浔江鱼类群落结构在蓄水后发生了一定的改变。
2017—2019年浔江鱼类所构建的生态网络模型呈现为服从幂律分布的无标度复杂网络。吴康等[48]认为,大多数网络连接集中于少数节点,这是网络结构化和非随机性演化趋势的反映。本研究结果表明,2017—2019年网络结构幂指数呈现递增趋势,表示网络中的集散节点数在减少,网络发生随机性演化,结构趋向于简单化。聚类系数可以用来阐述种间相互作用,聚类系数越低,物种间的联系越相似[49],本研究中聚类系数持续降低,表明浔江鱼类的种间联系越来越相似,网络复杂性降低。2020年的网络结构显示,此时α>3,网络中基本不存在离散节点,并且聚类系数为0,这可能会引起网络崩溃[33]。Dunne等[38]指出,低连接度的食物网总体或者部分会服从幂律分布,这与本文的研究结果类似。
浔江鱼类生态网络平均路径长度均为1,表明生态系统中的能量可以通过摄食关系快速在不同营养级的鱼类间传递[50],远低于海洋食物网路径长度的正常范围(1.3—3.7)[38]。大藤峡蓄水后,随着时间推移,鱼类种类数减少、网络中的节点数降低、网络密度呈减小趋势。鱼类群落结构主要受环境变化的影响[51],浔江位于大藤峡枢纽的下游,其水文状况会受到大坝的直接影响,环境因子的改变会驱使不同类型鱼类的迁入与迁出,从而会慢慢影响着鱼类的群落结构,从而引起生态网络的响应。
4.2 大藤峡蓄水对浔江部分经济鱼类的影响
本研究优势度排名前9的鱼类中,除了罗非鱼sp始终为中生态位种外,其他的均始终为广生态位种,鱼类优势度与时间生态位宽度大小呈一定的正相关性。生态位宽度是物种对环境适应及种间竞争和分布范围评估的重要指标,宽度值越大,该物种对资源的利用和环境适应能力越好[52]。大藤峡蓄水后东方墨头鱼时间生态位宽度增加最多,鲶下降最大,表明东方墨头鱼对改变后的环境适应更强,而鲶对当下环境适应度降低。大刺鳅与海南鲌时间生态位宽度发生了明显上升,由中生态位种变为广生态位种;东方墨头鱼与纹唇鱼由窄生态位种变为中生态位种,这些鱼类的时间生态位宽度增加可能是由于生存环境被压缩导致更容易被捕获所致。鲤为广布性鱼类,对环境的适应性较强[53],蓄水前后鲤的时间生态位宽度均保持在极高的水平,表明在浔江的鱼类中,鲤对环境资源的利用最强,其出现的时间维度最宽,更能适应季节变换引起的环境变化。Olsson等[54]指出,外来入侵鱼类具有更大的生态位,本研究中罗非鱼sp时间生态位宽度可以认为未发生变化,表明它对环境的适应能力极强,环境变化并不能让它的种群发生明显变动。

4.3 大藤峡蓄水对浔江鱼类群落构建机制的影响
Stegen等[60]认为生物群落构建可以分为两个确定过程(同质性选择、异质性选择)和两个随机过程(同质性扩散、具生态漂变的扩散限制)。本研究基于时间生态位重叠指数,构建零模型,发现浔江2017—2020年鱼类群落的构建取决于环境因素,生境过滤占主导作用,为非随机分布的分离格局,处于群落构建的异质性选择过程。牛克昌等[61]提出,在有限的资源条件下,相似物种会通过生态位分离以减少种间竞争,此时竞争排斥所造成的影响会超过生境过滤[62]。而2020年构建的零模型参数结果显示,标准化效应尺度降至2.70,实测值更接近模拟值,表明随着时间的推移,浔江鱼类群落趋向于由竞争主导的随机分布发展格局,栖息地的异质性有所降低;此外,不同年份各采样点鱼类NMDS分析发现蓄水后各采样点鱼类差异降低也支持这一观点。坝下鱼类群落所受的影响可能随着时间的推移而发生变化,环境稳定后物种间的相互作用可能会变得更加重要[63]。本研究发现,大藤峡枢纽蓄水后,浔江鱼类群落格局正在快速发生变化,虽然目前还处于非随机分布的分离格局,但是已经有明显向由竞争主导的随机分布格局发展的趋势。
5 结论
大藤峡蓄水后,浔江鱼类种类数减少,浔江鱼类ABC曲线揭示鱼类群落在蓄水后开始受到严重干扰,并随着时间的推移受到的干扰愈加严重;与此同时浔江鱼类生态网络复杂性降低,发生随机性演化。部分鱼类的时间生态位重叠值上升,某些鱼类种对的种间竞争加强;浔江鱼类群落由生境过滤占主导作用的非随机分布的分离格局向竞争主导的随机分布发展格局转变。因此,大藤峡蓄水已经对浔江鱼类群落造成较大负面影响,需及时进行保护与修复。建议大藤峡枢纽在履行水力发电、防洪泄洪、储水灌溉等功能的同时,也要兼顾保持河流的持续流动,在鱼类繁殖期进行生态流量补偿促进鱼类繁殖,此外,保持过鱼通道的畅通和保持一定的流速,以便洄游习性鱼类进行迁移。
猜你喜欢
水利水电快报(2023年2期)2023-02-21
人民珠江(2022年10期)2022-10-28
海洋信息技术与应用(2022年1期)2022-06-05
水泵技术(2021年3期)2021-01-22
中国新技术新产品(2020年17期)2020-11-30
儿童时代·幸福宝宝(2020年9期)2020-09-08
歌海(2016年6期)2017-01-10
金色少年(奇趣科普)(2016年8期)2016-09-21
中国水利(2015年13期)2015-02-28
城市道桥与防洪(2013年5期)2013-03-11
