
濒危植物永瓣藤叶片功能性状对环境因子的响应
2023-09-28 05:25:52宋帅帅蔡朝晖王世彤肖之强江明喜魏新增
生态学报 2023年17期
宋帅帅,王 建,蔡朝晖,吴 浩,王世彤,肖之强,江明喜,魏新增,*
1 西藏大学青藏高原生态与环境研究中心,拉萨 850000 2 西藏大学理学院,拉萨 850000 3 中国科学院武汉植物园,中国科学院水生植物与流域生态重点实验室,武汉 430074 4 中国科学院核心植物园保护生物学中心,武汉 430074 5 中国科学院大学,北京 100049 6 湖北九宫山国家级自然保护区管理局,通山 437100 7 湖北科技学院核技术与化学生物学院,咸宁 437100
植物与其生存环境之间的相互关系一直是植物生态学研究的重点。植物功能性状是植物在长期进化过程中,为了适应环境变化,减少环境的不良影响,所形成的形态、生理和物候特征[1],能反映植物对不同环境的响应和适应机制[2]。叶片是植物进行光合作用和蒸腾作用的主要器官,是植物与外界进行水、气、热交换的关键门户,对生态系统物质、能量循环具有重要的影响[3—4]。叶片性状与植物对资源的获取和利用关系密切[5],并且有数量多、易于获取、易于观测等优势[6]。
近年来,国内外学者开展了一系列叶功能性状与环境因子之间相关性的研究[7—8]。例如,环境光照增加或者水分减少时,叶片变小变厚、比叶面积降低,以增加主脉密度,提高水分利用效率[9—10]。目前,在区域尺度上叶片性状变异方面的研究,多数使用了物种平均值来研究叶片性状与环境之间的关系[11—12],忽略了种内性状变异性,而越来越多的研究表明种内性状变异与种间性状变异同样十分重要[13—15]。多物种分析表明,某些植物性状的种内变异高达40%[13],而全球荟萃分析将25%的群落内变异和32%的群落间变异归因于种内性状差异[14]。因此,研究不同环境条件下的种内性状变异,对于促进理解环境对性状变异的影响和预测物种对气候变化的响应至关重要[16]。
珍稀濒危植物由于分布范围有限、生境特殊或者数量稀少,其对环境变化更加敏感[17—18]。因此,对珍稀濒危植物叶功能性状的研究将有助于更好地开展濒危植物保护和管理工作。相比于珍稀濒危乔木和灌木物种,珍稀藤本植物的叶片功能性状研究相对较少[7,19—20]。永瓣藤(Monimopetalumchinense)是卫矛科(Celastraceae)永瓣藤属(Monimopetalum)的单种属藤本植物[21],为中国特有种,国家二级重点保护植物[22],被列为中国生物多样性红色名录濒危(EN)种[23]。现有研究主要集中于永瓣藤种群分布、结构与动态、多样性与繁育等方面[24—28],而对叶片功能性状的研究尚未见报道。本研究将以濒危植物永瓣藤(Monimopetalumchinense)为研究对象,对15项叶片形态与生理的功能性状进行测定,揭示永瓣藤叶功能性状的种内变异,并结合地理与环境因子,对永瓣藤叶功能性状指标与环境因子的相关性进行分析,揭示其对于异质环境的适应策略,以期为永瓣藤保护、管理与合理利用等工作提供科学依据。
1 材料与方法
1.1 研究区域
永瓣藤分布于湖北、江西、浙江和安徽4个省份的11个县市[26,29],主要分布区为江西省(7县市)和安徽省(2县市),湖北省唯一分布点为通山县洪港镇沙店村,浙江省唯一分布点为金华市婺城区[29]。分布范围介于28.837°N—29.973°N,114.841°E—119.476°E之间,南北跨度约为126km,东西跨度约为514km,海拔范围137—651m。分布区在我国长江中下游南部中亚热带向北亚热带过渡的区域,主要植被类型为常绿阔叶林和常绿落叶阔叶混交林[21],年均温15.3℃,年降水量1687.64mm[21,25],主要为酸性或中性土壤,土壤pH值介于4.53—7.82。气候数据从WorldClim 网站 (http://www.worldc lim.org/) 获取(表1)。

表1 永瓣藤种群地理位置、气候和土壤因子概况Table 1 Location, climate and soil factors of the 11 wild populations of Monimopetalum chinense
1.2 样品采集
2021 年7—8月,对4个省份的11个永瓣藤种群进行样品采集(图1和表1)。记录各种群的经纬度和海拔。种群间至少相隔 10km,种群内个体间至少相隔 10m,每株采集约30片成熟、无病虫害、完全展开、边缘完整(不带叶柄)的叶片。共采集到了209株、6292份叶片(表1),叶片样本现场称重并扫描后置于信封中带回实验室。同时,随机在每个样方内采集土壤样品,采用五点交叉取样法,钻取5份25cm深的土样,将其混合后装入自封袋带回实验室进行预处理(去除杂质并于阴凉处自然风干),以测定土壤化学指标。使用重铬酸钾容量法测定土壤有机质含量、使用酸度计测量土壤pH值、土壤硝态氮使用紫外分光光度法测定、土壤铵态氮使用纳氏试剂比色法测定、土壤有效磷使用钼锑抗比色法测定。
1.3 叶片性状测定
使用电子天平称取每株个体30片叶片鲜重。每个种群取随机选取3片叶片,用叶绿素仪 (Koni-ca Minolta SPAD-502Plus)测量其叶绿素含量。然后将所有叶片平铺于扫描仪(Epson PerfectionV19,分辨率设为300dpi)进行扫描,扫描的叶片图片用万深 LA-S叶分析软件进行批量处理,计算叶面积、叶周长、叶长、叶宽、叶长宽比、叶形状因子(公式1)[7]。将扫描后的叶片放入80 ℃烘箱,48h后取出,称其干重,然后结合叶干重和叶面积计算比叶面积(公式2),结合叶片干重和鲜重计算叶干物质含量(公式3)。叶碳含量和叶氮含量采用碳氮元素分析仪测定,叶磷含量采用钼锑抗比色法测定,最后计算得到碳氮比、碳磷比、氮磷比(表2)。
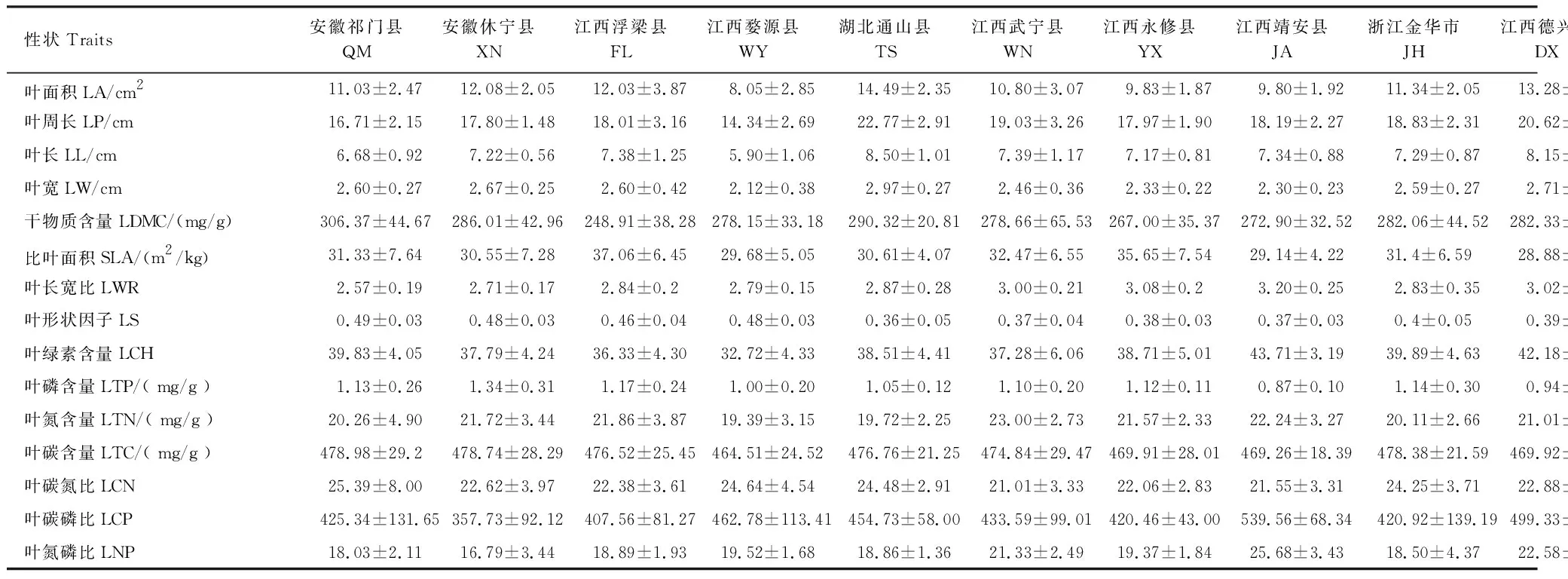
表2 永瓣藤野生种群叶片功能性状Table 2 Leaf functional traits of the 11 wild populations of Monimopetalum chinense

(1)

(2)

(3)
1.4 数据分析
将样点(单一种群)内定义为局域尺度,将整个采样区域(11个样点)定义为区域尺度,计算叶片性状的最大值、最小值、平均值、倍数范围(最大值除以最小值)和变异系数。对环境因子数据进行正态检验,不满足正态分布数据进行转换以满足正态分布和方差齐性。使用Pearson相关系数分析各功能性状之间、叶功能性状与环境因子之间的相关性。采用多元逐步回归分析环境因子对叶片性状的影响,使用向后逐步法选择具有最小赤池信息标准(AIC)值的最优模型[30]。数据处理在 R 4.0.5[31]中完成。
2 结果
2.1 叶片性状变异
叶片功能性状中,变异倍数范围从1.03—1.80倍。在局域尺度上,叶片功能性状变异系数从3.0%—22.5%不等,而在区域尺度上,变异系数从1.1%—15.5%不等。在局域尺度上,变异系数最大的5个性状分别是:叶面积、比叶面积、叶全磷含量、叶碳氮比、叶碳磷比,其中,叶面积是变异最大的叶片性状。在区域尺度上, 叶面积仍然是变异程度最大的,其次是叶氮磷比和叶形状因子,叶碳含量是变异程度最小的性状。大多数性状的局域变异系数都大于区域变异系数,除了叶形状因子以外(表3)。

表3 永瓣藤叶片功能性状的变异系数Table 3 Leaf traits variation of Monimopetalum chinense
2.2 叶片性状之间的相关性
叶功能性状之间的相关分析显示(图2),在叶片形态特征方面,叶面积、叶周长、叶长、叶宽之间相互呈显著正相关,叶片干物质含量与比叶面积显著负相关,叶形状因子与叶周长、叶长、叶长宽比显著负相关。叶生态化学计量特征方面,叶长宽比与叶磷含量、叶碳氮比显著负相关而与叶碳磷比、叶氮磷比显著正相关,叶磷含量与叶碳磷比、叶氮磷比显著负相关,叶氮含量与叶碳氮比显著负相关,叶碳磷比与叶氮磷比显著正相关。
2.3 叶片性状地理格局
叶功能性状与经纬度以及海拔的相关性分析表明,叶长宽比与纬度呈现显著负相关,叶形状因子与纬度呈现显著正相关(图3)。其它功能性状与经纬度以及海拔的关系均不显著。

图3 永瓣藤叶片长宽比和叶形态因子的纬度梯度格局Fig.3 Latitudinal patterns of leaf length-width ratio and leaf shape factor of Monimopetalum chinense
2.4 气候、土壤因子对叶片性状变异的影响
多元逐步回归分析结果表明(表4), 年均温、年降水量、土壤有机质、土壤pH、土壤硝态氮、土壤铵态氮、土壤有效磷对叶片性状都有明显的影响。叶长宽比随着年均温、年降水量以及土壤硝态氮增加和土壤有机质、土壤pH、土壤有效磷的减小而增加(R2= 0.97,P< 0.01)。叶形状因子随着土壤pH和土壤有效磷的增加而变大(R2= 0.85,P<0.001)在单个环境因子中,土壤pH、土壤硝态氮、土壤有效磷、年均温、年降水量、土壤铵态氮和土壤有机质含量分别能解释8个、4个、4个、3个、3个、1个、1个叶片功能性状。叶周长随着土壤pH和土壤硝态氮的增加而减小(R2= 0.60,P<0.05)。叶长随着土壤pH和土壤硝态氮的增加而减小(R2= 0.60,P<0.05)。叶氮含量随着年降水量和土壤硝态氮的增加和年均温、土壤有机质和土壤pH的减小而增加(R2=0.86,P<0.01)。叶氮磷比随着年均温、年降水量的增加与土壤pH、土壤硝态氮、土壤有效磷的降低而变大(R2= 0.84,P<0.05)。

表4 永瓣藤叶片功能性状与气候和土壤因子的多元线性回归模型Table 4 Multiple regression model of leaf functional traits of Monimopetalum chinense varying with climatic and soil factors
3 讨论
3.1 叶片性状的种内变异
永瓣藤叶片功能性状种内变异不均匀,多数性状在区域尺度上的变化值小于局域尺度,区域尺度上这与广泛的环境梯度适应有关[32]。在局域尺度上,永瓣藤15项叶片功能性状的变异系数范围在3.0%—22.5%之间,变异倍数范围在1.03—1.80之间,叶性状变异的不同幅度反映了植物适应不同环境波动的权衡。叶面积、比叶面积、磷含量、碳氮比和碳磷比的变异程度最高,这些性状与植物对光、养分、和水资源的竞争密切相关[33—35]。叶面积的变异程度最大,叶片大小直接影响着植物对光的截取和对碳的获取能力,并且可以通过影响叶片边界层厚度进而影响叶片温度调节和蒸腾速率,进而影响叶片热量和水平衡[10]。在藤本植物中,磷与其它性状之间连接较多,磷可能在调节多种植物功能方面显得更为重要,这些性状上的环境选择通常会影响整个表型[36]。永瓣藤叶片碳、氮含量的变异程度相对较小,磷含量变异系数相对较大,叶片氮(N)、磷(P)和N∶P比值分别为21.15mg/g、1.08mg/g和20.09。与全球陆生植物叶片N、P含量和N∶P(18.74mg/g 、1.21mg/g 和15.55)[37—39]相比,永瓣藤叶片磷含量相对较低。同时,永瓣藤分布区土壤有效磷含量也偏低(2.76mg/kg),这与谢国文等[40]对永瓣藤的研究结果一致,说明,永瓣藤的生长受到了磷限制。
3.2 叶片性状之间的相关性
永瓣藤叶干物质含量与比叶面积显著负相关,即越厚的叶片其干物质含量也越高,这是永瓣藤为了适应炎热的生长环境[41—42]。永瓣藤叶长宽比与叶磷含量、叶碳氮比显著负相关。一般认为,叶的宽窄能影响叶的光和面积,同时通过改变叶的周长-面积比,影响叶边界层的水汽交换[39],叶磷含量调控植物生理过程(特别是生长速率),叶片碳氮比叶片碳含量与生产力之间的关系[43—44]。永瓣藤较长的叶片其叶磷含量较低,叶碳氮比也较低,说明较长叶片的永瓣藤生长速率较低,受到了更严重的磷限制。
3.3 叶片性状对环境因子的响应
植物通过调整叶片边界层厚度来优化叶肉细胞的温度,以提高光合作用酶的活性[38],较宽的叶片其边界层较厚,能减少水汽交换[45],因此,生活在凉爽地区的植物往往具有较小的叶片长宽比(即较宽的叶片),以保持较厚的边界层和较高的叶片温度以进行光合作用,而温暖潮湿的地区,叶片往往具有较大的叶片长宽比(即较长的叶片),以提高植物叶脉的水分输送能力[10,46]。本研究中,叶长宽比随着纬度增加显著减小,叶形态指数随着纬度增加显著增加,即叶片随着纬度增加逐渐变得更宽,更趋近于圆形,这种模式可能反映了植物的热力学调节与水力限制之间的权衡,以响应跨空间的气候变化[47]。这一结果印证了全球变暖会增加植物叶长宽比的结论[48]。
多元逐步回归分析表明(表4),没有任何单个环境因子可以解释所有的叶片性状变异,环境和叶片功能性状变异之间的关联比较复杂。25%—97%的叶片性状变异能由气候和土壤因子组合解释,但不包括叶干物质含量,这可能是由于其分布的遗传适应。在所有的环境因子中,土壤pH、土壤硝态氮、土壤有效磷是解释度最高的三个,年均温和年降水量的解释度次之。这说明永瓣藤叶片性状变异主要由土壤因子驱动,同时受到气候因子的影响。先前研究指出,土壤肥力和土壤pH与叶片功能性状显著相关[14,48—49]。永瓣藤叶周长、叶长都受到土壤pH与土壤硝态氮的显著负效应影响,即土壤pH值越低、土壤硝态氮含量越低,叶周长和叶长越大。在物种或群落水平上, 植物叶片氮和磷含量的分布格局一般呈现随年平均气温和降水量的增加而降低的趋势[50],本研究的结果与此趋势是一致的,永瓣藤叶片氮含量与磷含量分别受到土壤硝态氮与土壤有效磷的正效应影响,而受到年均温与年降水量负效应的影响。在温度和水分充足的情况下,永瓣藤叶片趋向于慢速生长的保守策略。
叶功能性状是植物策略的良好预测指标。虽然永瓣藤受到了环境的限制,但叶片功能性状通过与环境因子间的相互作用,利用种内变异和性状组合,能够良好适应当前环境因子的变异。
4 结论
本文探讨了濒危植物永瓣藤的叶片功能性状变异以及对环境因子的响应与适应策略,有助于了解我国亚热带低山藤本植物叶片功能性状的地理格局及其种内变异的环境驱动力。叶片性状之间的变异程度不同,局域尺度性状变异值甚至高于区域尺度。叶片面积的变异程度最高。叶片的形态受到纬度梯度的影响,随着纬度的上升,永瓣藤叶片通过变得宽且圆来增加叶片边界层厚度,降低叶片水热散失, 这种模式可能反映了植物的热力学调节与水力限制之间的权衡,以响应跨空间的气候变化。永瓣藤叶片磷含量较小,表明永瓣藤的生长受到了磷限制。土壤pH、土壤硝态氮与土壤有效磷是永瓣藤叶片性状变异的主导因子,年均温与年降水量也能影响部分叶片性状变异。本研究在分布区尺度上揭示了永瓣藤叶片性状的变异情况,并分析了叶片性状与环境的关系,为保护濒危植物和藤本植物提供新的见解。
猜你喜欢
湖南农业大学学报(自然科学版)(2021年3期)2021-07-02 01:48:06
趣味(数学)(2020年4期)2020-07-27 01:44:16
世界科学技术-中医药现代化(2020年2期)2020-07-25 02:06:08
支部建设(2020年15期)2020-07-08 12:34:32
新疆农业科学(2020年1期)2020-02-14 03:22:46
临床医药文献杂志(电子版)(2017年11期)2017-05-17 04:47:56
青苹果·教育研究版(2016年7期)2016-12-12 05:19:52
中学生物学(2016年8期)2016-01-18 09:08:21
中国麻业科学(2015年5期)2015-12-28 06:22:11
百科知识(2015年18期)2015-09-10 07:22:44
