
氮沉降和刈割条件下羊草光响应模型比较及响应特性
2023-09-28 05:25杨斯琪鲍雅静叶佳琦徐梦冉吕晓涛姜良超韩兴国
生态学报 2023年17期
杨斯琪,鲍雅静,*,叶佳琦,2,吴 帅,张 萌,徐梦冉,赵 钰,吕晓涛,姜良超,韩兴国
1 大连民族大学环境与资源学院,大连 116600 2 内蒙古大学生态与环境学院,呼和浩特 010021 3 中国科学院沈阳应用生态研究所额尔古纳森林草原过渡带生态系统研究站,沈阳 110016 4 中国科学院植物研究所植被与环境变化国家重点实验室,北京 100093 5 中国科学院大学,北京 100049
光合作用是植物生命活动的基础,在整个生态系统固碳的过程中发挥重要作用,同时也是衡量环境对植物影响的重要指标之一[1]。光响应曲线能反映植物净光合速率随光照强度的变化趋势,现已成为研究植物光合生理生态学的重要手段之一。通过拟合光响应曲线可获得光合作用的最大光合速率、表观量子效率、光饱和点、光补偿点和暗呼吸速率等参数[2—3]。目前应用最广泛的模型为直角双曲线模型[4]、非直角双曲线模型[5]、指数模型[6]和直角双曲线修正模型[7]。而上述模型的拟合效果和模型适用性因推导机理不同在不同植物中产生差异[8—10]。
羊草(Leymuschinensis)是中国温带半干旱草原上最具优势的物种之一,具有适口性强、产量高、营养物质丰富等特点,并且还具有抗寒、耐盐碱、耐牧、耐刈割等特性[11]。在植被恢复和环境治理方面,羊草是最理想的草种,目前在东北以及内蒙古等地区已经开展了大量对羊草的栽培工作,主要应用于恢复受到盐碱、干旱、滥割、滥牧等原因造成的草原退化[12]。但目前,关于羊草的研究多集中于生物量、生态化学计量、抗逆性等方面展开大量研究[13—16],但关于其光响应特性的研究却鲜有报道,且已有研究多集中于不同氮、磷添加或水分条件下的响应[17—20]。此外,在这些研究中,作者均直接采用一种光响应模型进行拟合分析,并未讨论此模型是否适合羊草的光响应曲线拟合,这就可能造成羊草光合特性研究的偏差。
人类活动使大气氮沉降显著增加,研究指出全球的氮素临界负荷为100 Tg/a,而预计到2050年全球的大气氮沉降总量可能会达到195 Tg/a[21]。土壤理化性质因氮沉降发生改变,植物生长和光合生理特性也会受到影响[22—24]。刈割是一种人为的对草地进行资源索取与利用的技术手段,目前已成为了除放牧以外最重要和最普遍的草场管理方式[25]。近年来,氮沉降与刈割对植物生长的交互影响逐渐受到关注。研究表明,刈割改变了氮富集对土壤微生物[26]、养分利用效率[27]和土壤化学性质[23]等方面的影响,但缺乏不同光响应模型在氮添加和刈割处理下对植物光响应过程的适用性,并且目前关于氮添加和刈割处理对植物光合特性影响的研究相对较少[28—29]。因此,筛选适合氮添加和刈割处理下羊草光响应过程的拟合模型并探讨羊草光合特性对环境变化的响应,对进一步理解和研究羊草的光合作用特性具有重要意义,通过分析其对环境变化的响应机制可制定合理的草原管理措施。
本研究以在内蒙古草原进行为期7年野外氮添加控制试验平台的羊草为研究对象,在不同氮添加浓度和刈割处理下,分别测定其光合作用的光响应曲线,采用4种常用光响应模型(直角双曲线模型、非直角双曲线模型、指数模型和直角双曲线修正模型)进行拟合分析。本研究拟解决以下科学问题:1、筛选出氮添加和刈割下羊草的最适光响应拟合模型;2、探讨羊草光响应特性与氮添加和刈割的关系。研究结果将为深入了解羊草的光合生理生态特征与草原管理和植被恢复提供参考。
1 材料与方法
1.1 实验设计
本研究地点位于中国科学院沈阳应用生态研究所额尔古纳森林草原过渡带生态系统定位研究站(50°12′N, 119°30′E,海拔523 m)。样地长期年平均降水量为375 mm,年平均气温为-3℃[26]。该地区的天然大气氮沉降量为1—2 g N m-2a-1[30]。该生态系统的优势种有羊草(Leymuschinensis)、贝加尔针茅(Stipabaicalensis)和糙隐子草(Cleistogenessquarrosa)等[26],土壤类型为黑钙土。
加氮试验始于2014年5月底,采用随机完全区组设计。所施氮肥为硝酸铵(NH4NO3),有6种氮添加水平(0、2、5、10、20 g N m-2a-1和50 g N m-2a-1)以及刈割(刈割与非刈割)交叉处理,共有12个处理,每个处理重复8次(共96个处理区)。每个样方的面积为10 m×10 m,样方之间设2 m缓冲带。每年5月下旬施用一次氮肥。将肥料与沙子(每块0.5公斤)混合,用手均匀撒播。每年八月下旬在离土壤表面10 cm的高度进行刈割。收获的生物量从样方中移除。其中2021年封育地的羊草相对生物量为66.81%,表层土壤(0—5 cm)的氮含量如下(表1)。
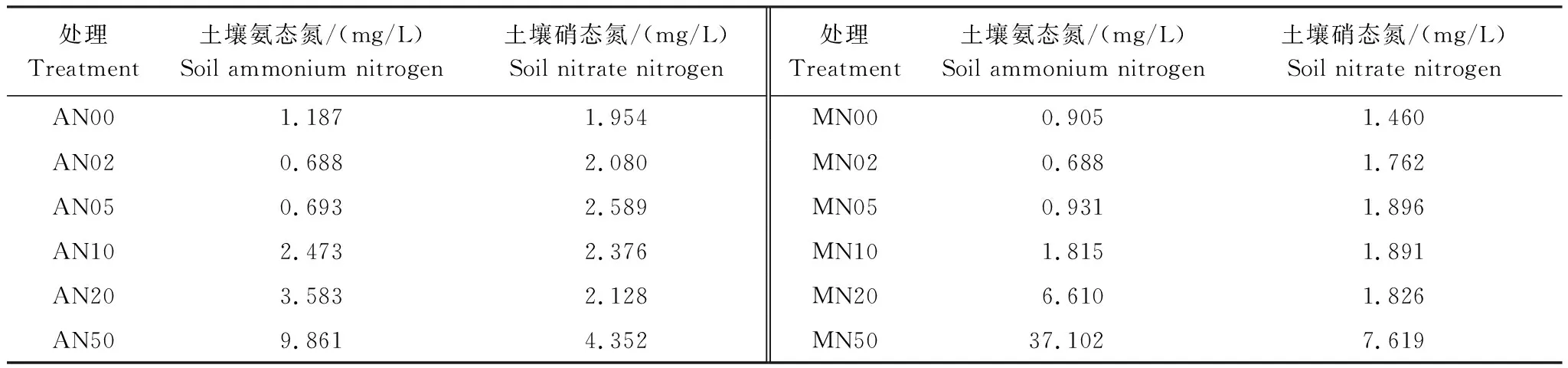
表1 不同氮添加和刈割处理样地的土壤氨态氮和硝态氮含量Table 1 The contents of ammonia nitrogen and nitrate nitrogen in soils with different nitrogen addition and mowing treatments
1.2 光合-光响应曲线测定
于2021年8月采用Li-6400光合作用测定系统(Li-Cor, Inc., 林肯市, 内布拉斯加州,美国)进行光合-光响应曲线测定。每个处理随机选取长势良好且相同叶位的羊草叶片,于晴朗无风天气的9:00—11:30之间测定羊草的光响应过程。光强梯度设置为2000、1800、1600、1400、1200、1000、800、400、300、200、100、50、20 μmol m-2s-1和0 μmol m-2s-1,设置每次改变光强后数据采集时间为120—300 s,气体流速为500 μmol/s,叶室内CO2浓度为390 μmol/mol,空气相对湿度控制在60%左右,温度设置为25℃。净光合速率(Pn)、气孔导度(Gs)、胞间CO2浓度(Ci)、蒸腾速率(Tr)等光合指标由仪器自动记录,并计算水分利用效率(Water use efficiency, WUE)为Pn/Tr。
1.3 模型拟合
1.3.1直角双曲线模型
直角双曲线模型[31]的表达式为:
(1)
式中,Pn为净光合速率;I为光合有效辐射强度;α为初始量子效率;Rd为暗呼吸速率;Pnmax为最大净光合速率。
1.3.2非直角双曲线模型
非直角双曲线模型[32]的表达式为:
(2)
式中,θ为非直角双曲线的曲角,其它参数意义同公式(1)。
1.3.3直角双曲线修正模型
直角双曲线修正模型[33]的表达式为:
(3)
式中,β,γ为修正系数,其它参数意义同公式(1)。
1.3.4指数模型
指数模型[9]的表达式为:
(4)
式中,e为自然对数的底,其它参数意义同公式(1)。
1.4 模型评价
4种模型的拟合优度采用均方根误差(RMSE)、平均绝对误差(MAE)和决定系数(R2)进行评价。计算公式如下[34]:
(5)
(6)
式中,yo,i和ym,i分别为净光合速率的实测值和拟合值;n为样本数。
1.5 数据处理与分析
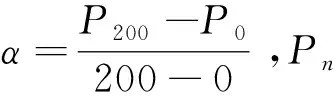
在SPSS 23.0统计软件单因素方差分析和Duncan多重比较,Origin 2021中绘图。
2 结果与分析
2.1 不同光响应模型对光响应曲线的参数拟合效果及拟合优度
不同模型在不同氮添加浓度和刈割处理下对羊草叶片光合-光响应曲线的参数拟合效果上有一定差异(表2、图1)。指数模型的α拟合值与实测值无显著差异(P>0.05),拟合效果最好,而其他三种模型的拟合值均与实测值有显著差异(P<0.05),且均大于实测值(图1)。四种模型的LSP拟合值与实测值均具有显著差异(P<0.05),但指数模型的拟合值与实测值相对接近,直角双曲线模型和非直角双曲线模型的LSP拟合值均显著低于实测值(P<0.05),直角双曲线修正模型的LSP拟合值误差较大(图1)。此外,直角双曲线模型和非直角双曲线模型拟合的Pnmax均高于实测值(图1)。四种模型均能很好的拟合LCP和Rd,与实测值均无显著差异(P>0.05)。因此,4种模型对光响应参数的拟合准确度由高到低为指数模型、直角双曲线修正模型、非直角双曲线模型和直角双曲线模型。
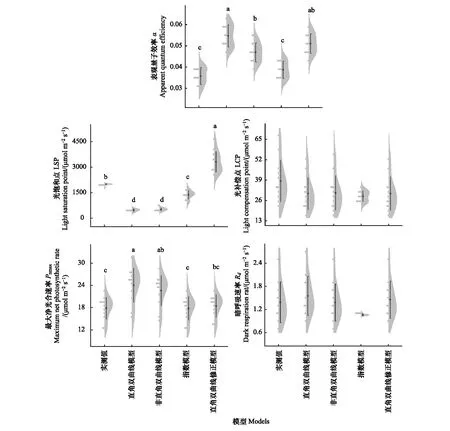
图1 4种光响应模型对羊草的光合参数拟合值与实测值对比Fig.1 Comparison between the fitting values of photosynthetic parameters for four light response models and the measured values of L. chinensis不同小写字母表示处理间差异显著(P<0.05)
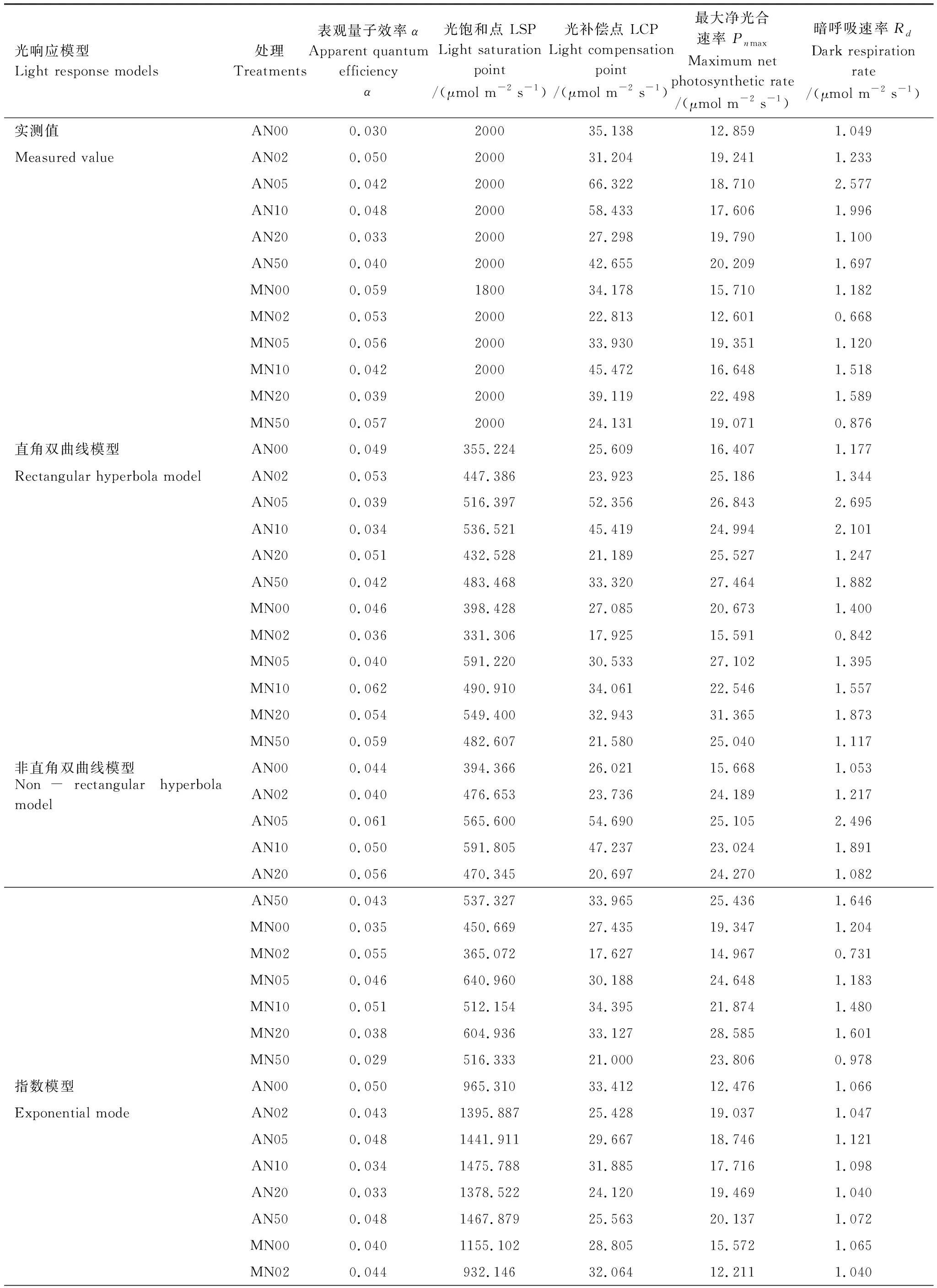
表2 羊草光合-光响应曲线参数实测值与拟合值Table 2 The measured data of photosynthesis-light response curves parameters of L. chinensis and the fitted values by models
图2为4种光响应模型拟合的光响应曲线结果,可知在不同氮添加浓度和刈割处理下,羊草光响应曲线均能由直角双曲线模型、非直角双曲线模型和直角双曲线修正模型较好的拟合,但指数模型的拟合点普遍高于实测点。因此从宏观拟合情况来看,指数模型的拟合效果最差,其余三种模型的拟合效果较好。通过RMSE、MAE和R2进一步评估4种光响应模型的拟合优度。由图3可知,指数模型的RMSE、MAE和R2和其他三种模型均存在显著差异(P<0.05),且RMSE和MAE的值比其他模型高,R2比其他模型低,因此其拟合优度最差。非直角双曲线模型的RMSE和MAE显著低于直角双曲线模型,直角双曲线修正模型的RMSE和MAE与非直角双曲线模型和直角双曲线模型无显著差异,且这三种模型的R2间无显著差异。综合来看,4种模型的拟合优度由高到低分别非直角双曲线模型、直角双曲线修正模型、直角双曲线模型和指数模型。
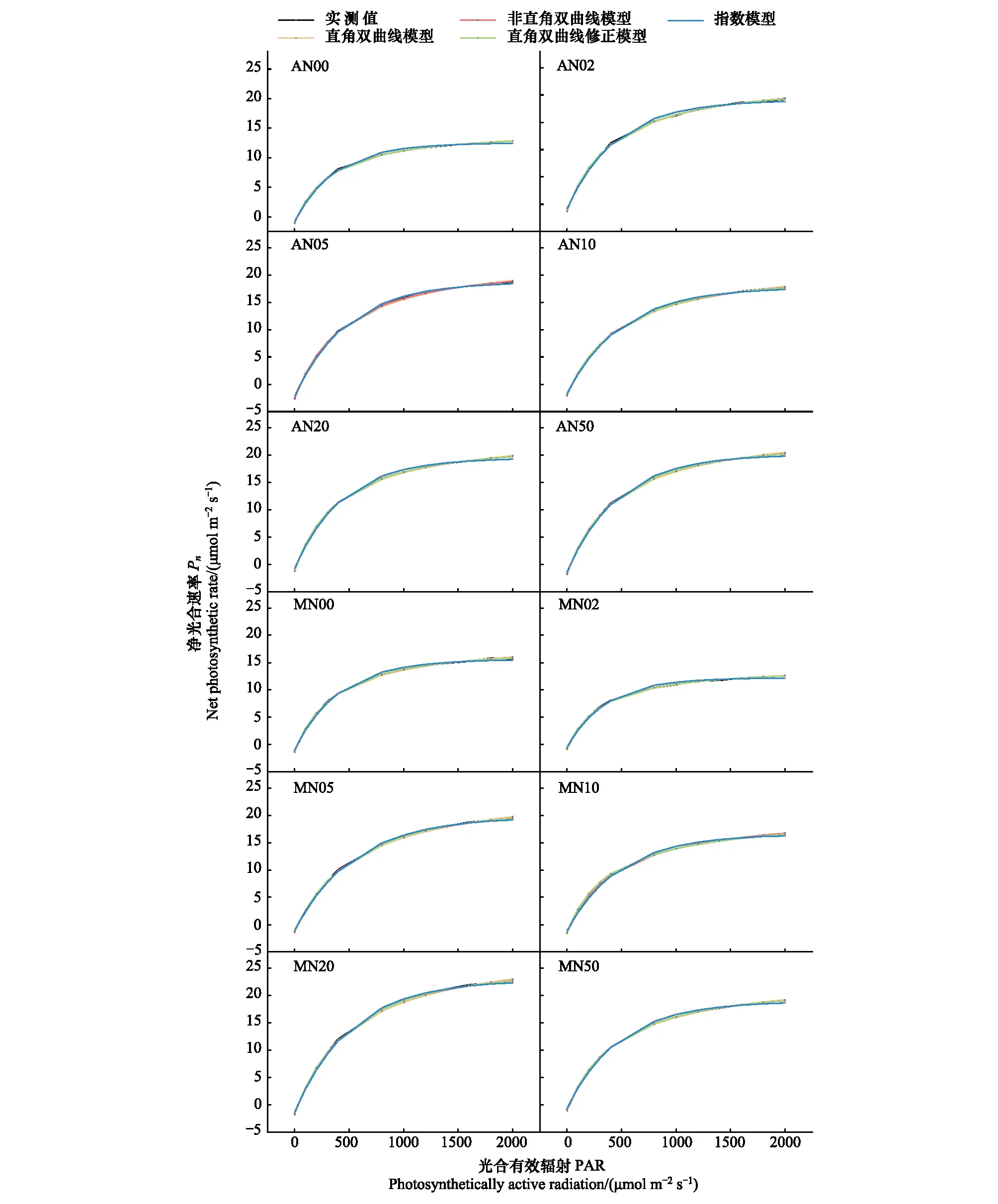
图2 氮添加和刈割处理下不同光响应模型对羊草光合作用光响应曲线的模拟Fig.2 Simulation of photosynthesis-light response curves of L. chinensis by different light response models under nitrogen and mowing treatmentsA:不刈割Unmown; M:刈割Mown; N0: 0 g N m-2 a-1; N2: 2 g N m-2 a-1; N5: 5 g N m-2 a-1; N10: 10 g N m-2 a-1; N20: 20 g N m-2 a-1; N50: 50 g N m-2 a-1
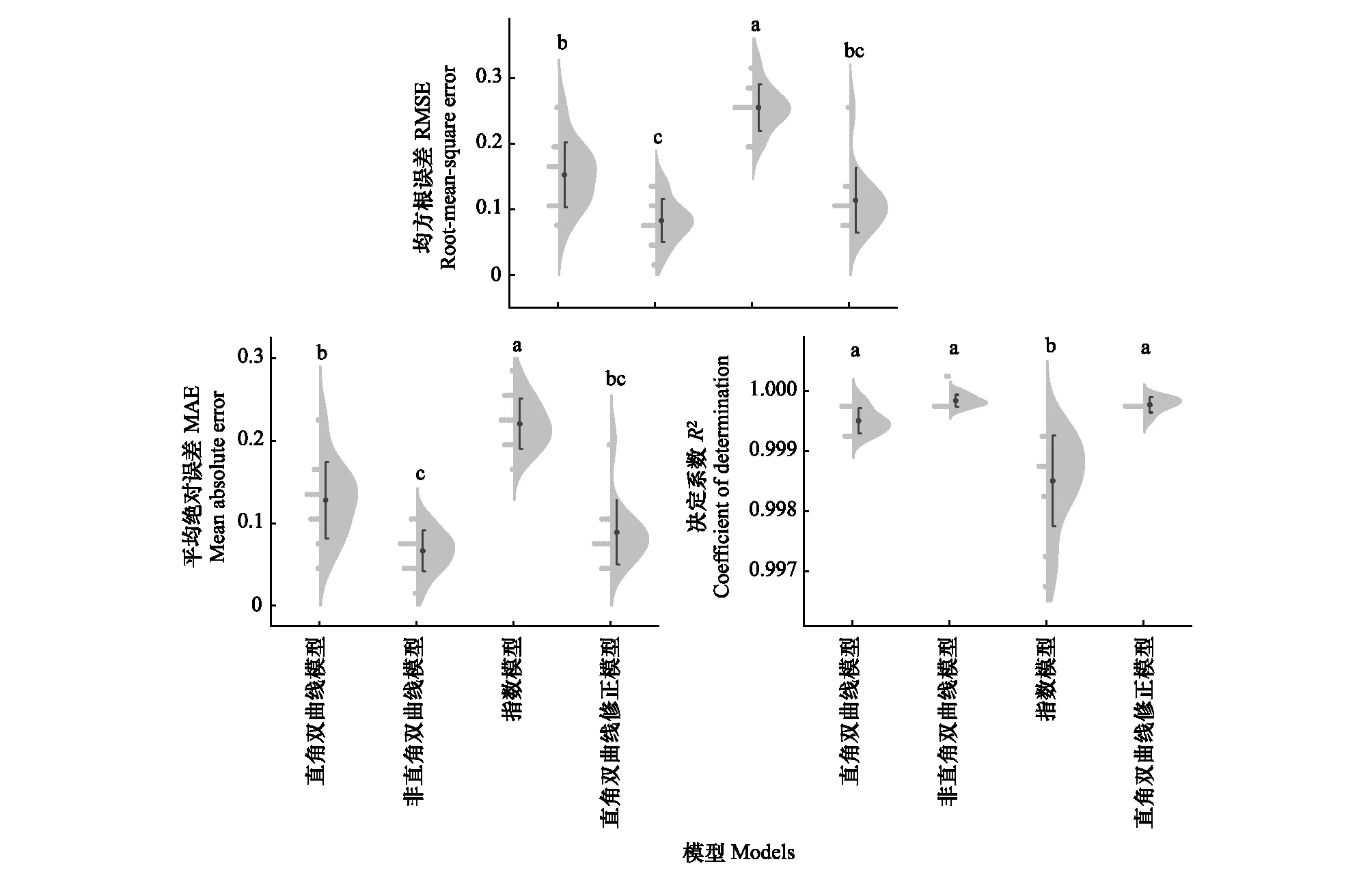
图3 4种光响应模型拟合优度对比Fig.3 Comparison of goodness-of-fit on four light response models
通过比较氮添加和刈割处理下4种模型对羊草光响应拟合值与实测值的差异和模型拟合优度可知,参数拟合程度并不代表模型拟合优度。总体上,指数模型的参数拟合效果最好,非直角双曲线模型的模型拟合优度最高,而直角双曲线模型和直角双曲线修正模型的拟合效果较差。
2.2 氮添加和刈割处理下羊草的光响应变化趋势及特征参数比较
根据上述不同光响应模型拟合效果及拟合优度分析结果,在比较氮添加和刈割处理下羊草光合作用光响应变化趋势时采用非直角双曲线模型的拟合值(图4)。结果显示不同处理的羊草光响应曲线具有明显差异,且波动范围较大(图4)。在光强≤400 μmol m-2s-1时,所有处理下的Pn均表现为随光合有效辐射(PAR)的增加而迅速上升而后趋于平缓的趋势,表现出强光下饱和趋近型光响应曲线的特点。氮浓度升高,羊草的Pn逐渐增加,但各施氮组间相差不大,说明高浓度氮添加对羊草的光合作用影响较小。刈割扩大了羊草Pn随氮添加浓度的波动范围。在所有处理中,羊草均能维持较高的Pn,并未出现明显的光抑制现象。此外,PAR越大,各处理组之间的Pn差值也越大,MN20处理下的羊草表现出最强的光合能力,N20是光能利用率最高的氮素临界值。
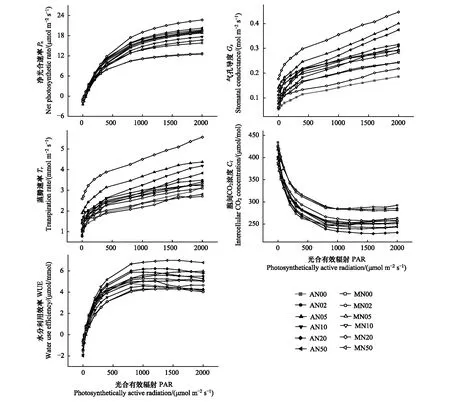
图4 不同氮添加和刈割处理下羊草光合作用光响应曲线Fig.4 Photosynthesis light response curves of L. chinensis under different nitrogen and mowing treatments
图4为氮添加和刈割处理下羊草Gs和Tr的光响应过程。可知在不同处理下,羊草Gs和Tr的光响应均表现为随PAR增加而上升的趋势。MN20处理下的Gs和Tr明显高于其他处理,这与Pn的响应结果一致。说明过量施氮并不利于气孔导度的增加。此外,Tr和Pn的响应规律一致,因为光合作用的进行需要水分的连续供应,借助水分的运载能力不断吸收营养成分和矿质元素[34]。
由图4中氮添加和刈割处理下羊草Ci的光响应过程可见,在光强为0—400 μmol m-2s-1时,所有处理下的Ci均随PAR的增加而迅速下降,在光强为400—1000 μmol m-2s-1时缓慢下降,在光强为1000—2000 μmol m-2s-1时趋于稳定。因为在初期Pn的迅速上升消耗了大量CO2,导致Ci的迅速下降,此时光合效率较高,当光强继续增加,Pn增长缓慢,Ci的下降趋势也减缓,直到趋于稳定。图4为氮添加和刈割处理下羊草WUE的光响应过程。与Pn的响应相似,在低光强范围内(0—400 μmol m-2s-1),WUE迅速上升,而随PAR的增加逐渐趋于稳定。极端环境下,植物能调整自身的光合作用机制,减少过量氮添加的负面影响,维持光合效率,保证正常生长。
通过比较四种模型的参数拟合效果,本文采用指数模型的拟合值进行光合参数分析(表1、表2)。表观量子效率(α)和光补偿点(LCP)能够反映植物利用弱光的能力[32]。非刈割和刈割处理下α的浮动范围分别为:0.033—0.044、0.034—0.044,LCP的浮动范围分别为:24.120—33.412 μmol m-2s-1、24.527—32.064 μmol m-2s-1。在所有加氮处理中,N20的α显著高于不加氮处理(N00)、LCP显著低于不加氮处理(P<0.05)(表3),说明N20处理的羊草在弱光下的光合能力较强,而N00处理的羊草在弱光下的光合能力较弱。
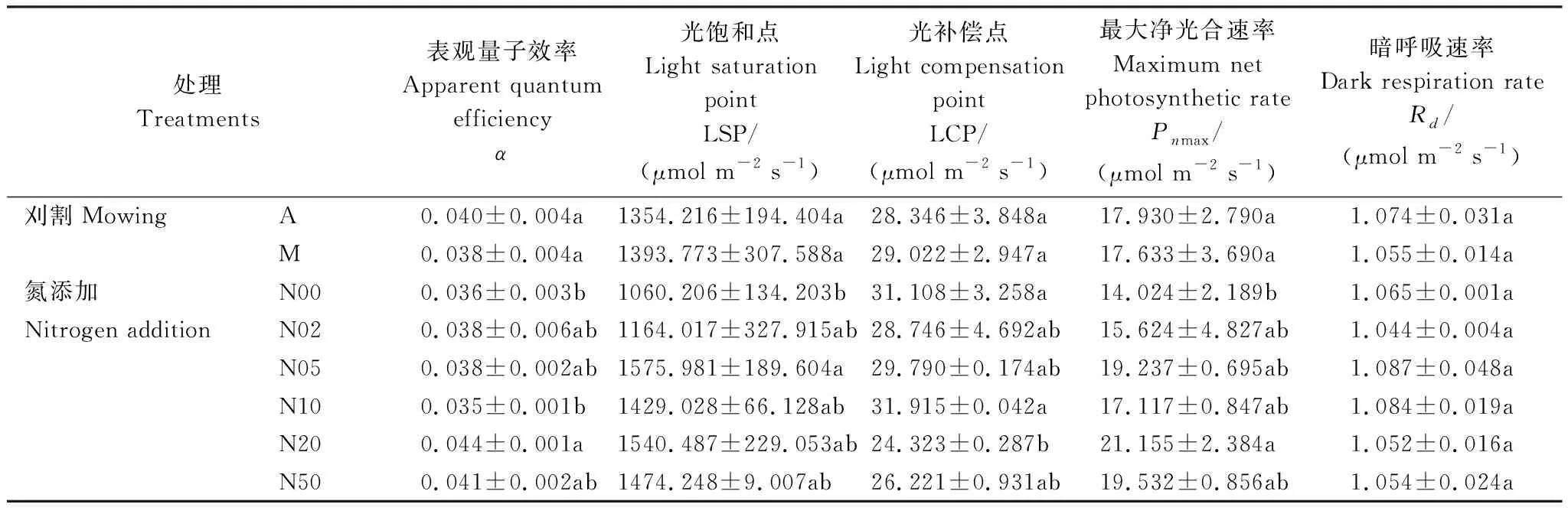
表3 氮添加和刈割下羊草的光响应曲线参数比较Table 3 Comparison of light response curve parameters of L. chinensis under nitrogen addition and mowing
最大净光合速率(Pnmax)代表了植物在最适光强下的光合作用能力,反映了植物的最大光合潜力[35]。不刈割处理的Pnmax范围为:12.476—20.137 μmol m-2s-1,刈割处理扩大了Pnmax的浮动范围,为12.211—22.840 μmol m-2s-1。此外,只有N20处理的羊草Pnmax显著高于不加氮处理(P<0.05)(表3)。
光饱和点(LSP)是衡量植物利用强光能力的一个指标[36]。刈割和不刈割处理的羊草LSP,均表现为随氮添加浓度增加而增加的趋势,且刈割处理(932.146—1710.051 μmol m-2s-1)的LSP浮动范围比非刈割处理(965.310—1475.788 μmol m-2s-1)的宽。N20处理的羊草LSP相比N0有显著提高(P<0.05)(表3)。
暗呼吸速率(Rd)表示植物正常生理活动中提供的必须能量[36]。非刈割处理下Rd的浮动范围为:1.040—1.121 μmol m-2s-1,刈割处理下Rd的浮动范围为:1.037—1.071 μmol m-2s-1,可见刈割处理的羊草通过呼吸消耗的光合产物更少,能够积累更多的有机物。
3 讨论
植物光合作用的光响应曲线是描述光量子通量密度与植物净光合速率之间关系的曲线[37],利用光响应模型拟合参数是评价光合效率和探究植物光合作用响应机制的重要手段[5]。在众多模型中,直角双曲线模型、指数模型和非直角双曲线模型是一条没有极值的渐进线,无法很好地模拟强光下的光抑制现象[33],此时进行光响应参数拟合时,常出现Pnmax拟合值大于实测值、LSP拟合值小于实测值的情况[38—40]。而叶子飘等为更好的拟合光抑制阶段的响应趋势,提出了直角双曲线修正模型,并且此模型可以直接算出LSP,目前已经证明直角双曲线修正模型在达乌里胡枝子[41]、杠柳[42]、胡杨[43]、苦槠[44]等植物的光响应曲线拟合效果上好于其他模型。但在本研究中,4种模型对光响应参数的拟合效果为:指数模型>直角双曲线修正模型>非直角双曲线模型>直角双曲线模型。此外,利用RMSE、MAE和R2对4种模型进行拟合优度比较后发现,非直角双曲线模型对氮添加和刈割处理下的羊草光响应曲线模拟效果最好。这是因为在所有处理下,羊草均没有表现出明显的光抑制现象。俞芹等在对景宁木兰(Magnoliasinostellata)的研究中发现,指数模型对Pnmax、LSP求解较准确,LCP采用直角双曲线修正模型的拟合结果较好,而直角双曲线模型拟合Rd的效果最佳[45]。李雪琴等对蕨类植物光响应模型的拟合比较中发现,直角双曲线修正模型拟合的LSP效果较好,但非直角双曲线模型的拟合精度最高[34]。因此,在对氮添加和刈割处理下的羊草进行光响应曲线拟合时,可以结合两种模型的优势。
氮素是组成植物叶绿素、Rubisco酶及光合器官的重要元素,是影响植物光合生理过程的重要因子[46]。本研究结果显示,在光强为0—400 μmol m-2s-1时,随PAR增加Pn增加,这有利于CO2同化,产生较多的干物质,且Gs、Tr和WUE与Pn的响应类似,这是因为植物能根据植株自身状况在保持正常生理活动前提下,通过调节气孔的开闭程度,合理协调Pn与Gs之间的关系,从而调节WUE的变化,达到最多的碳固定量与最少的水分散失量,以适应外界环境条件变化[32,47]。同时,Ci的响应趋势与Gs相反,说明Pn的增加主要受非气孔因素的影响,这与前人的研究结果一致[46]。随光强的进一步增加,各指标的响应过程因施氮浓度而异。随氮浓度升高,Pn增加幅度变小,基本稳定在同一水平,说明在不刈割处理下,可能由于多年的连续氮素添加产生的累积效应,使氮对羊草光合的促进作用达到饱和,无法进一步提升光合能力。刈割扩大了羊草Pn的波动范围,尤其是在MN20处理下,羊草Pn、Gs和Tr都比不刈割处理的高,可能是在较高氮素添加下,刈割去除了地上部分,促进新生植株对养分的吸收,减缓了高氮的抑制作用,从而提高光能利用效率。
表观量子效率(α)反映了植物在弱光下吸收、转换和利用光能能力[48]。研究表明,在适宜条件下一般植物α在0.03—0.05之间[48]。前人研究表明,水稻、玉米和杂交象草的α在不同氮添加浓度下分别为0.020—0.045、0.028—0.043和0.049—0.069[29,37,49]。本研究发现羊草的α在0.033—0.044之间,说明在弱光下羊草的光能利用效率相对较高。α随施氮量的增加而增加,因为氮添加缓解了氮素限制,使羊草光合作用受抑制的程度有所降低,导致植物在弱光下吸收、转换和利用光能的能力得到提升。羊草Pnmax随着氮素含量的增加呈现先增后趋于平缓的趋势,这说明适量氮添加有利于提升羊草的光合潜能。刈割进一步提高了氮添加处理下羊草的光合潜能,在MN20处理时,氮添加和刈割的交互作用加强了羊草利用强光的能力,制造有机物的能力有所提升。LSP和LCP代表了植物的光合有效范围[50]。研究发现,随氮添加浓度增加,羊草的耐阴能力和耐强光能力逐渐提升,由N0处理的33.412—965.310 μmol m-2s-1增长到N50处理的25.563—1467.879 μmol m-2s-1。刈割普遍提升了氮添加处理下的光合有效范围,其中MN20的光合有效范围为24.527—1702.453 μmol m-2s-1。Rd随氮浓度升高呈现“Λ”变化,表明羊草在养分贫瘠和高氮胁迫环境下通过减少呼吸的养分消耗提高生存能力。羊草光合机构对环境变化的适应能力较强,光合潜力较大。
光合作用的各个参数并不是独立存在的,植物利用各个参数间的协同互作应对环境变化,以保证正常生长。在合理的施氮范围内并刈割使羊草叶片的光合性能得到提升。MN20处理虽然蒸腾速率较高和水分利用效率较低造成了水分耗散,但通过提高气孔导度、光能利用潜力并扩大光强适应范围等途径使羊草叶片的光合作用在所有处理中达到最优。
4 结论
综上所述,非直角双曲线模型对氮添加和刈割处理下羊草叶片光响应曲线的拟合优度最高,指数模型的参数拟合效果最好。通过对羊草的光响应曲线和拟合参数分析可知,适度氮添加能促进羊草光合能力,但过度氮添加并没有进一步提高光合能力。刈割能拓宽光照生态幅,提升光能利用率和减少呼吸消耗光合产物,从而使氮添加下尤其是高氮(N20)添加下羊草的光合能力进一步提升。因此,在未来全球大气氮沉降逐渐增加的情况下,将有利于内蒙古草原羊草的生长发育,并且氮添加对羊草光合作用的影响受到刈割的影响。研究结果不仅有助于加强关于氮沉降对内蒙古草原生态系统碳固定的认识,还有助于未来全球变化情景下陆地生态系统管理措施的制定。
猜你喜欢
湖南林业科技(2021年3期)2021-12-02
——拟合优度检验与SAS实现
四川精神卫生(2021年5期)2021-11-04
现代畜牧科技(2021年7期)2021-07-28
现代畜牧科技(2021年4期)2021-07-21
小学生学习指导(低年级)(2019年9期)2019-09-25
中学生数理化·中考版(2019年8期)2019-07-13
中学生数理化·七年级数学人教版(2019年4期)2019-05-20
中学生数理化·七年级数学人教版(2017年9期)2017-08-15
当代化工研究(2016年6期)2016-03-20
现代畜牧科技(2016年9期)2016-02-21
