
桦褐孔菌酚类化合物激活过氧化氢酶的作用及机制
2023-09-13 12:49:02郝瑞林单树花胡晓燕张志怡李荣山李卓玉
山西农业科学 2023年9期
郝瑞林 ,单树花 ,胡晓燕 ,张志怡 ,李荣山 ,李卓玉
(1.山西大学 生物技术研究所,山西 太原 030006;2.忻州师范学院 生物系,山西 忻州 034000;3.山西省人民医院,山西 太原 030006)
桦褐孔菌(Inonotus obliquus)属于多孔菌属,是一种罕见的白腐真菌,主要寄生在桦树上[1-2]。桦褐孔菌主要分布在北纬45°~50°的寒冷地区,如中国的东北部以及北美和北欧的森林带[3]。从桦褐孔菌中发现了丰富的次级代谢产物如多糖、萜类化合物、多酚以及黑色素[4-7]。桦褐孔菌的代谢产物显示出优异的预防肿瘤、改善炎症反应、调节血糖代谢、免疫和改善氧化应激压力的能力[8-13],作为主要代谢物,桦褐孔菌来源的多酚显示出多种生物学活性[14-16]。从桦褐孔菌中提取的3,4-二羟基苯甲醛肟(3,4-Dihydroxybenzaldoxime)对白血病和癌症细胞具有强烈的细胞毒性作用[17]。儿茶酚丙烷类化合物可以通过阻断拓扑异构酶II,抑制癌细胞的生长[18]。此外,酚类化合物可以显著降低IL-6和TNFα,进而改善炎症[19]。桦褐孔菌含有多种酚类化合物,可作为强效抗氧化剂[20]。桦褐孔菌的多酚提取物可防止过氧化氢诱导的氧化应激,而多糖、三萜和类固醇提取物无此作用[21]。目前,已从桦褐孔菌中鉴定出多种酚类化合物,包括在IO中鉴定出许多多酚,包括咖啡酸、3,4-二羟基二苯甲酮、没食子酸、丁香酸、原儿茶醛、3,4-二羟苯甲醛、2,5-二羟基对苯二甲酸、Phelligridin C-I和 methylinoscavin A-B[18,22-23]。在这些酚类化合物中,Phelligridin D、Phelligridin E 和Phelligridin G显示出较强的改善氧化应激压力的能力[23]。
活性氧(ROS)在O2还原过程中产生,包括超氧阴离子(O2·-)、过氧化氢(H2O2)和羟基自由基(OH·)[24-25]。ROS可导致细胞中大分子的氧化损伤,并导致细胞功能障碍[24]。过氧化氢酶是已知的ROS防御酶家族之一,能够催化H2O2的分解,进而缓解由H2O2过量累积导致的氧化损伤[24,26-27]。糖尿病、骨肉瘤、结直肠癌以及膀胱癌等病理导致的过氧化氢酶活性降低是细胞中过氧化氢累积以及氧化应激压力升高的主要原因[28-31]。CAT可以将代谢过程以及歧化反应所产生的副产品过氧化氢转变为氧和水,清除体内的过氧化氢,从而使细胞免于过氧化氢的损害[32]。桦褐孔菌醇提取物和水提取物可以有效提高CAT的活性,进而改善氧化应激所诱发的炎症反应[33-34]。酚类化合物是桦褐孔菌重要的组成成分,通过酚类化合物激活过氧化氢酶可能是桦褐孔菌缓解氧化应激压力并改善相关疾病的重要途径。
本研究应用钼酸铵显色法评估了桦褐孔菌酚类化合物对过氧化氢酶的调节作用,利用分子动力学模拟进一步分析了其具体机制,旨在阐明小分子化合物调控过氧化氢酶活性的可能机制,为抗氧化酶调节剂的开发提供参考。
1 材料和方法
1.1 试验材料
供试桦褐孔菌子实体,从吉林省延边朝鲜族自治州敦化市农贸市场购买,子实体经60 ℃烘干至恒质量备用。
1.2 试剂、仪器与设备
色谱纯甲醇和乙腈(赛默飞世尔科技股份有限公司(马萨诸塞州,美国));来源于牛肝的过氧化氢酶,由碧云天生物技术研究所(上海,中国)提供;过氧化氢酶活性检测试剂盒(南京建成生物工程研究所)。
LC-16P制备型高效液相色谱仪(日本,岛津);LC-20AT高效液相色谱仪(日本,岛津);LC-MS8050三重四级杆高效液相色谱质谱联用仪(日本,岛津);Synergy HTX全波长酶标仪(美国,安捷伦)。
1.3 试验方法
1.3.1 桦褐孔菌多酚的提取与分离纯化 桦褐孔菌粉碎后过0.25 mm筛,提取和纯化过程参考文献[35]进行。纯化后的桦褐孔菌酚类化合物通过高效液相色谱法(HPLC)进一步分离,并使用反相C-18制备柱(10 mm×250 mm,10 μm)。使用20% CH3CN(5%甲酸)作为洗脱剂,以流速10 mL/min洗脱90 min后得到12个馏分。通过HPLC-DAD-MS/MS对馏分进行表征[36],馏分在60 ℃减压浓缩后经过冷冻干燥,将冷冻干燥后的样品溶于蒸馏水后过0.22 μm滤膜。过滤后的样品注入LC-MS8050高效液相色谱质谱联用仪,通过反相柱Poroshell 120 SB-C18(150 mm×4.60 mm,3.5 μm)分离和鉴定。分离采用0.1%甲酸水溶液(A)和0.1%甲酸乙腈(B)作为流动相,洗脱梯度为0~5 min,5% B;5~45 min,5%~45% B;45~47 min,45%~5% B;47~50 min,5% B。流速为1.0 mL/min,进样体积为20 μL。电喷雾电离(ESI)质谱法用于鉴定阳离子和阴离子模式下的酚类化合物。质谱条件为电压3.5 kV,干燥气体为氮气加热至250 ℃,毛细管温度为300 ℃,采集范围为100~1 000 m/z。
1.3.2 过氧化氢酶活性检测 过氧化氢酶分解H2O2的反应可通过加入钼酸铵而迅速中止,剩余的H2O2与钼酸铵作用产生一种淡黄色的络合物,在405 nm 处测定其变化量,可计算出过氧化氢酶的活力。牛肝过氧化氢酶溶于50 mmol/L磷酸钠缓冲液(pH值7.0),得到过氧化氢酶储备液(4.0×10-9mol/L)。制备浓度为1 mmol/L的桦褐孔菌酚类化合物的储备溶液,并将其添加到过氧化氢酶储备溶液中,混合液与反应缓冲液混匀后在37 ℃下孵育5 min。根据过氧化氢酶活性检测试剂盒说明书,继续操作并利用全波长酶标仪,在405 nm处测定吸光值(A)。
其中,A化合物+过氧化氢酶表示化合物与过氧化氢酶混合后反应液的吸光值,A化合物表示只含有桦褐孔菌酚类化合物的反应液的吸光值,A空白表示用蒸馏水等体积代替过氧化氢酶和酚类化合物的反应液的吸光值。
1.3.3 分子对接 使用AutoDock Vina软件进行的计算对接研究桦褐孔菌酚类化合物对过氧化氢酶的影响[37]。从蛋白质结构数据库(PDB,https://www.rcsb.org/)中获得过氧化氢酶(PDB ID:1TGU)的PDB格式文件。在对接模拟之前,使用UCSF Chimera程序将蛋白质的能量最小化[38],用PyMOL软件去除蛋白质结构中的杂原子、水分子和重复链[39]。使用Autodock工具完成蛋白质的脱氢、Gasteiger电荷的计算和非极性氢原子的融合[40]。处理后的蛋白质结构文件保存为pdbqt格式。为了确保过氧化氢酶的活性位点在对接过程中被完全覆盖并且配体可以自由移动,建立了尺寸为60 Å×60 Å×60 Å的网格盒,对接中心的X、Y、Z轴坐标分别为15.781、17.888和13.220。最大对接次数设置为50次。对接结果用PyMOL可视化和分析。
1.3.4 分子动力学模拟 为了收集更详细的结合信息,使用GROMACS 2020.4对Phelligridin E与过氧化氢酶的对接结果进行了分子动力学模拟(MD)。使用acpype程序,GAFF力场用于预处理Phelligridin E。amber14ffSB力场用于处理过氧化氢酶。acpype软件用于创建配体拓扑文件。设置大小为1 nm的盒子,通过添加Na+,使Phelligridin E/过氧化氢酶复合物的表面电荷降低到0后进行时间为50 ns的分子动力学模拟(MD)。使用VMD对模拟过程中Phelligridin E/过氧化氢酶复合物的轨道和结构进行均方根偏差(RMSD)、溶剂可及表面积(SAS)、氢键分析和结合自由能分解。
1.4 数据分析
所有数据结果均以平均值±标准偏差表示,统计分析用Excel和SPSS 26.0软件进行处理。
2 结果与分析
2.1 桦褐孔菌酚类化合物的鉴定
应用HPLC-DAD-MS分析用于鉴定馏分1~12,结果如表1所示。

表1 HPLC-DAD-MS法鉴定桦褐孔菌中的酚类化合物Tab.1 Identification of phenolic compounds from Inonotus obliquus by HPLC-DAD-MS
由表1可知,馏分1~12被鉴定为原儿茶醛、丁香酸、8-去甲基桉树素、2,5-二羟基对苯二甲酸、柚皮素、3-羟基-2-氧代苯-4,6-二羧酸、儿茶素、(E)-3,4-二羟基苯亚甲基丙酮、Phelligridin E、Methylinoscavin B、Interfungin B和Phelligridin I。
2.2 桦褐孔菌酚类化合物对过氧化氢酶活性的影响
过氧化氢酶是生物体抵抗氧化应激的重要屏障,在许多氧化应激相关的疾病中都出现了过氧化氢酶活性降低的现象[41]。通过激活过氧化氢酶等抗氧化酶的活性从而改善氧化应激导致的相关疾病是一种有效的策略。为了进一步探索桦褐孔菌酚类化合物应用于干预氧化应激相关疾病的可能性,本研究利用体外试验分析了桦褐孔菌酚类化合物对过氧化氢酶活性的影响,结果显示(图1),12种桦褐孔菌酚类化合物中仅有原儿茶醛、Phelligridin E 和 Phelligridin I显示出增强过氧化氢酶活性的潜能,特别是Phelligridin E。经终浓度分别为0.5、1.0、2.0、3.0、4.0、5.0 μmol/L的Phelligridin E处理后,过氧化氢酶的活性分别增加了41.90%、44.68%、52.66%、58.10%、62.78%和68.10%。
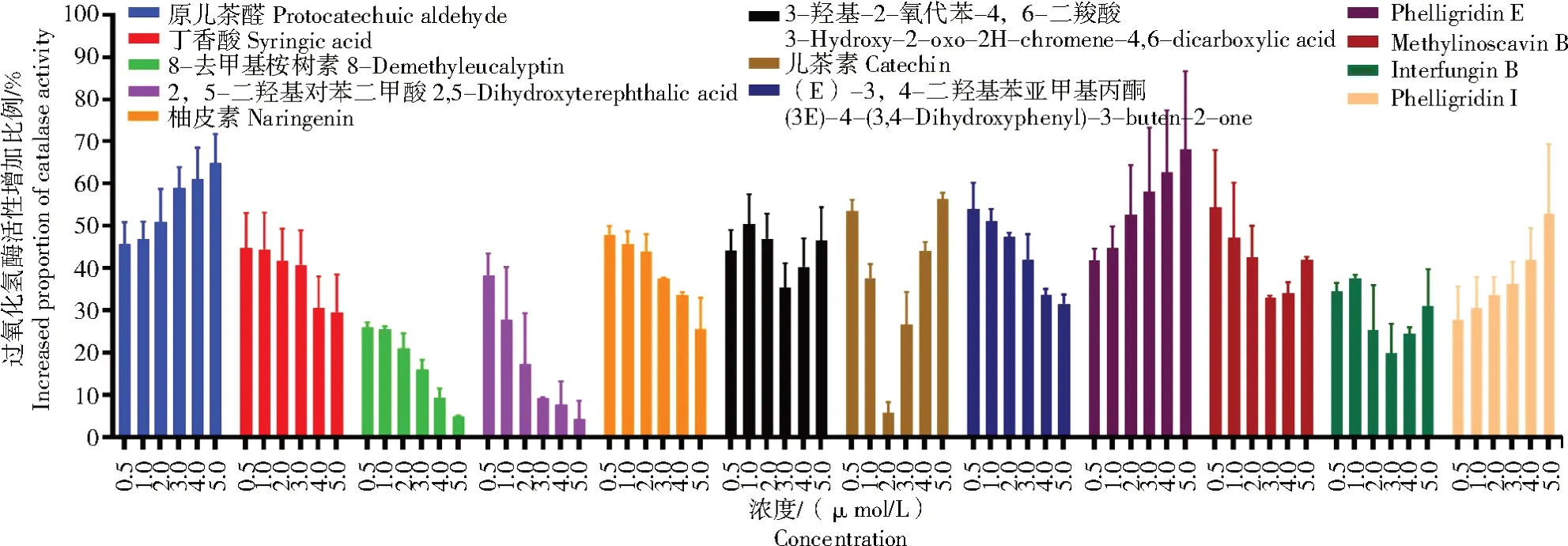
图1 桦褐孔菌中的酚类化合物对过氧化氢酶活性的影响Fig.1 Effect of phenolic compounds from Inonotus obliquus on catalase′s activity
2.3 桦褐孔菌酚类化合物与过氧化氢酶的对接能量
为了分析桦褐孔菌酚类化合物激活过氧化氢酶的可能机制,应用分子对接技术比较了12个馏分与过氧化氢酶的结合自由能。由表2可知,在12种酚类化合物中,苯乙烯基吡喃酮类多酚与过氧化氢酶的结合能低于其他多酚,特别是Phelligridin E。提示Phelligridin E可能通过与过氧化氢酶的结合来增强酶的活性。

表2 过氧化氢酶与桦褐孔菌酚类化合物的结合自由能Tab.2 The binding energy between catalase and phenolic compounds from Inonotus obliquus kcal/mol
2.4 分子动力学模拟
2.4.1 分子动力学模拟稳定性评价 通过检查过氧化氢酶以及过氧化氢酶/Phelligridin E复合物在动力学模拟过程中的势能、温度、压力和密度的变化来评价分子动力学模拟的质量。如图2所示,在模拟过程中,过氧化氢酶和过氧化氢酶/Phelligridin E复合物的体系势能和密度保持了稳定,而体系温度和压力分别保持在300 K和 0 Pa,表明对过氧化氢酶以及过氧化氢酶/Phelligridin E复合物的模拟是高质量的。

图2 过氧化氢酶和过氧化氢酶/Phelligridin E的分子动力学模拟(MD)评估Fig.2 Evaluation of MD(molecular dynamics) simulations of catalase and catalase/Phelligridin E
2.4.2 RMSD、RMSF、SAS和Rg分析 对比分析过氧化氢酶以及过氧化氢酶/Phelligridin E复合物在分子动力学模拟过程中的RMSD、RMSF、Rg和SAS,结果如图3所示。从图3可以看出,在模拟过程中,结合Phelligridin E后,过氧化氢酶的RMSD出现明显波动;在模拟结束时,RMSD稳定保持在0.6 nm。在与Phelligridin E结合的过程中,过氧化氢酶的0—100、400—450位的氨基酸残基出现了较大的RMSF值变化。这些位置的氨基酸残基可能参与了过氧化氢酶与Phelligridin E的结合及配体结合后引起的蛋白质结构的变化过程。与Phelligridin E结合后,过氧化氢酶的Rg轻微增大;ROG常用于表征蛋白质空间结构的紧凑程度,ROG分析结果表明,Phelligridin E可能使过氧化氢酶的空间结构变得松散。与Phelligridin E结合后,过氧化氢酶的SAS呈现出下降趋势,表明Phelligridin E的结合可能使过氧化氢酶的亲水性下降。

图3 过氧化氢酶和过氧化氢酶/Phelligridin E复合物的50 ns分子动力学模拟结果Fig.3 Results of 50 ns molecular dynamics simulations of catalase and catalase/Phelligridin E complex
2.4.3 Phelligeidin E与过氧化氢酶相互作用分析 由图4可知,在结合Phelligridin E之后,过氧化氢酶的20—50位以及360—400位的氨基酸残基的空间构象发生明显变化(白色箭头表明结合Phelligridin E前后氨基酸残基的构象有明显变化)。这一改变与过氧化氢酶的RMSF的波动是一致的,过氧化氢酶空间结构的变化是由于酶与Phelligridin E之间的相互作用导致的。图4-B、C展示了Phelligridin E相对于过氧化氢酶的空间分布,图4-D、E展示了应用Protein_Ligand Interaction Profiler和LigPlot分析MD模拟第50 ns时的过氧化氢酶与Phelligridin E之间的相互作用。Protein_Ligand Interaction Profiler能够对蛋白质和配体做全面分析,包括氢键、疏水作用力和离子键分析[41],而LigPlot能够对蛋白质和配体之间形成氢键的可能做更充分的评估[42]。Protein_Ligand Interaction Profiler分析结果显示,Phelligridin E与过氧化氢酶的ASN369、HIS372、ILE373形成较强的疏水作用力(图4-D)。LigPlot分析结果显示,Phelligridin E与过氧化氢酶的ASP25、ASN369、HIS372、PRO391和CYS393形成氢键(图4-E)。图4-F显示了MD模拟过程中Phelligridin E与过氧化氢酶间氢键的动态变化,在50 ns内,过氧化氢酶与Phelligridin E间的氢键数量在2~6个。本研究利用gmx_MMPBSA对过氧化氢酶模拟过程中的吉布斯自由能进行了分解,结果显示,过氧化氢酶的GLU67、ILE373、PRO391、PHE326和MET392对Phelligridin E与过氧化氢酶的结合贡献较大(图4-G)。其中,ILE373与Phelligridin E形成距离为3.63 Å的疏水作用力,PRO391与Phelligridin E形成距离为2.54 Å的氢键,但过氧化氢酶的GLU67未与Phelligridin E形成4 Å范围内的作用力。GLU67在空间上位于ASP25附近,ASP25与Phelligridin E之间的氢键可能导致GLU67的位置发生改变。以上分析表明,氢键和疏水作用力在促进Phelligridin E与过氧化氢酶的结合中可能同样重要。
2.4.4 二级结构的变化 通过对MD模拟前后蛋白质二级结构组成的对比分析,发现在结合Phelligridin E后,过氧化氢酶的二级结构组成发生明显改变。如图5显示,Phelligridin E与过氧化氢酶之间的相互作用会导致酶分子的33—43、44—53、393—403、433—438位的氨基酸残基所形成的二级结构发生改变。这些变化具体为40—43位以及50—53位氨基酸残基倾向构建B-Sheet,393—400位氨基酸残基倾向构建5-Helix,433—437位氨基酸残基倾向构建3-Helix。

图5 由过氧化氢酶和过氧化氢酶/Phelligridin E复合物的所有氨基酸残基形成的二级结构Fig.5 Secondary structure formed by all amino acid residues of catalase and catalase/Phelligridin E complex
2.4.5 过氧化氢酶的氨基酸残基的二面角的变化 Phelligridin E 与过氧化氢酶间的相互作用力促进酶分子的VAL41、CYS393、ASP396、GLY399、GLY400发生二面角扭转(图6),二面角扭转具体为ASN33-GLN53倾向形成B-Sheet,CYS393-ASN403形成5-Helix,ASN433-ASP438形成3-Helix。

图6 过氧化氢酶和过氧化氢酶/Phelligridin E复合物氨基酸残基的拉氏构象Fig.6 Ramachandran plot of amino acid residues of catalase and catalase/Phelligridin E complex
2.5 Phelligridin E 激活过氧化氢酶的可能机制
图7-A详细显示了H2O2进入过氧化氢酶活性位点的主要通道,H2O2进入过氧化氢酶的通道被不同的氨基酸组包围,包括由ALA333-ASN338形成的左侧的随机折叠片结构、由GLU71-GLY78形成的右侧的双螺旋结构、由TYR325-ILE332形成的上侧的α-螺旋结构和由PRO151-PRO158形成的下侧的无规则卷曲。为了分析Phelligridin E结合后过氧化氢酶活性位点的主要通道的大小,对模拟前后的通道的左右两侧和上下两侧之间的距离进行了分析。图7-B结果显示,结合Phelligridin E后,底物进入通道的左右两侧之间的距离无明显变化,而上下两侧氨基酸基团之间的距离明显增加。因此,推测由于上下氨基酸残基之间的距离增加,底物进入活性位点的途径增加,过氧化氢酶的活性也增加(图7-C)。

图7 Phelligridin E激活过氧化氢酶的机制Fig.7 Mechanism of Pheligridin E activating catalase
3 结论与讨论
过氧化氢酶含有几个到达深埋活性位点血红素基团的通道。高度限制性的进入通道(称为主通道)作为底物(H2O2)到达活性位点的首选途径,并且只能通过该通道(主通道)进入活性位点血红素基团。通道的大小与这4个组的移动有关,其大小决定了进入活性位点的衬底的量[43]。Phelligridin E与过氧化氢酶通过ASN369、HIS372、ILE373形成较强的疏水作用力,通过ASP25、ASN369、HIS372、PRO391和CYS393,形成氢键。Phelligridin E与过氧化氢酶之间的疏水作用力和氢键使过氧化氢酶的VAL41、CYS393、ASP396、GLY399、GLY400发生二面角扭转,这种二面角扭转推动ASN33-GLN53形成B-Sheet,CYS393-ASN403形成5-Helix,ASN433-ASP438形成3-Helix,其中,CYS393-ASN403形成的5-Helix和ASN433-ASP438形成的3-Helix,使活性中心上下两侧的氨基酸残基之间的距离增加,使底物进入活性中心的入口增大,底物更容易进入包含血红素辅基的活性中心,增强了过氧化氢酶消除过氧化氢的能力。
猜你喜欢
中学生理科应试(2024年1期)2024-05-18 13:02:52
特产研究(2024年1期)2024-03-12 05:40:24
生物化学与生物物理进展(2022年6期)2022-07-21 11:52:06
中国食用菌(2021年10期)2021-12-01 11:21:10
中学生数理化(高中版.高考理化)(2021年2期)2021-03-19 08:52:38
池州学院学报(2015年3期)2016-01-05 01:13:04
天津科技大学学报(2015年2期)2015-08-09 01:40:42
食品工业科技(2014年13期)2014-03-11 18:16:49
食品工业科技(2014年13期)2014-03-11 18:16:45
原子与分子物理学报(2014年3期)2014-02-28 22:18:23
