
赤水乌骨鸡PPP3CA基因多态性与生长性状关联分析
2023-09-12 00:57蒋桂荣王昌毅
西南农业学报 2023年7期
周 迪,蒋桂荣,欧 仁,谢 艳,李 波,王 燕,杨 蓉,李 俊,沈 银,敖 叶,王昌毅
(1.贵州省种畜禽种质测定中心,贵阳 550018,2.贵州省畜禽遗传资源管理站,贵阳 550000)
【研究意义】赤水乌骨鸡是贵州省地方优良品种资源之一,主产于贵州省赤水市,是赤水市独有的地方特色家禽,贵州省极具特色的小型肉蛋兼用型鸡种,其具有体型大,单冠、平头,胸宽、体深、腹部大、丰满,脚长短适中,羽毛疏松均匀,黑色为主,耳垂紫黑色,喙、腿、爪为黑色的外貌特点[1]。赤水乌骨鸡肉质鲜嫩,富含人体所需氨基酸、微量元素、矿物质和黑色素,具有耐粗饲、耗料少、适应性强、肉质优良、耐热性好、抗潮湿、抗疲劳及觅食能力强等优良特性。2010年,农业部批准对赤水乌骨鸡实施农产品地理标志登记保护,加大了对赤水乌骨鸡的保种,至此展开了赤水乌骨鸡分子遗传的相关研究。但目前,关于赤水乌骨鸡主要经济性状的主效分子遗传机制尚不清楚,通过候选基因研究赤水乌骨鸡生长性状,揭示相关性状基因的遗传效应,对赤水乌骨鸡遗传改良、分子育种及保种等具有重要意义。【前人研究进展】蛋白磷酸酶3(Protein phosphatase 3,PPP3)又称钙调磷酸酶(CalcineurinCaN)是受Ca2+/钙调素(Calmodulin,CaM)调节的丝氨酸-苏氨酸蛋白磷酸酶,共有3种催化亚基,分别为PPP3CA、PPP3CB和PPP3CC[2],其中PPP3CA是蛋白磷酸酶3(PPP3)的催化亚基A,广泛存在于真核生物的多种细胞中[3-4]。在骨骼肌生长发育中,PPP3主要参与成肌细胞分化、成熟肌纤维肥大、钙的信号传导以及细胞内钙浓度变化的调节[5]。然而PPP3CA又是一种去磷酸化的丝氨酸-苏氨酸磷酸酶,在动物发育过程中至关重要,尤其是骨骼肌的生长发育[6]。研究表明,PPP3CA基因还在调控肌纤维中起关键作用,在不同肌肉组织中均有表达,并能够抑制成纤维细胞生长因子23(FGF23)的表达,与IGFⅠ共同参与骨骼肌生长发育的调节[7-9]。Shao等[10]研究发现,侏儒症白旗鱼的脊椎较短且骨骼发育不良,其PPP3CA基因的表达量显著低于骨骼发育正常的白旗鱼,表明PPP3CA基因低表达能引起骨骼发育不良。同时,Chen等[11]研究发现,PPP3CA基因不仅与肌肉生长有关,还与脂肪沉积有关,主要参与肌肉细胞增殖、脂质代谢和脂肪酸降解等过程。此外,PPP3CA基因还参与了动物繁殖调控,PPP3CA基因对性早熟、雌激素信号转导通路和卵母细胞减数分裂具有重要影响,并在精原干细胞减数分裂和精子发生过程中具有去磷酸化作用[12-13]。在卵母细胞发育过程中PPP3CA亚基表达量增加,而在卵母细胞受精时PPP3CB亚基的表达量显著增加,表明PPP3的Aβ亚基可作为辅助生殖技术程序中卵母细胞选择的标志基因[14]。另有研究发现,PPP3CA基因能调控鸡的横纹肌细胞及成肌细胞的分化,并参与骨骼肌纤维成分组成,涉及能量代谢和肌肉生长发育[15-16]。【本研究切入点】有关PPP3CA基因在家禽(如赤水乌骨鸡)及其生长相关性状方面的研究鲜见报道。【拟解决的关键问题】以赤水乌骨鸡为研究对象,PPP3CA基因为候选基因,采用PCR扩增和直接测序技术对其PPP3CA基因全部外显子进行多态位点筛选,运用RNAfold WebServer和Structural Bioinformatics Group在线软件预测PPP3CA蛋白质的二级结构和三级结构,计算其基因频率、基因型频率、纯合度、杂合度、有效等位基因数和多态信息含量,并对Hardy-Weinberg平衡状态进行分析;采用SPSS探讨PPP3CA基因多态性与鸡生长性状的相关性,明确其SNP位点的具体遗传效应,以期为赤水乌骨鸡开发利用提供分子理论基础,并对进一步加快其遗传改良和分子育种提供科学数据。
1 材料与方法
1.1 材料
1.1.1 赤水乌骨鸡 试验用赤水乌骨鸡由贵州绿源禽业有限公司提供。在相同饲养管理水平下进行分笼饲养,自由采食和饮水,选取具有生长记录、体况健康、200日龄的赤水乌骨鸡159羽。翅静脉采集血液,EDTA抗凝,-40 ℃保存备用。
1.1.2 试剂 E.Z.N.A.TMNRBC Blood DNA Kit试剂盒由Omega有限公司生产;2×Taq Master Mix(Dye Plus)由Vazyme有限公司生产;DL2 000 Marker由北京Solarbio(索莱宝)科技有限公司生产;琼脂糖由Hispanagar S A有限公司生产;无水乙醇由贵州塞兰博科技有限公司提供。
1.1.3 引物设计 根据GenBank已公布的鸡PPP3CA基因(登录号:NC_052535.1)序列,采用Primer Premier 5.0软件设计引物(表1),引物由安徽通用生物系统有限公司合成。
1.2 方法
1.2.1 血液DNA提取 根据E.Z.N.A.TMNRBC Blood DNA Kit试剂盒提取血液中DNA,取1 μL于紫外分光光度仪上检测DNA浓度和纯度,将合格的DNA于-20 ℃保存。
1.2.2 PCR反应及基因分型检测 每个样本取1 μL DNA进行混池,PCR体系(30 μL):2×TaqMaster Mix 15 μL;Forward Primer 2 μL;Reverse Primer 2 μL;混池DNA/DNA 2 μL;ddH2O 9 μL。PCR程序:95 ℃预变性3 min;95 ℃变性30 s,退火30 s(表1),72 ℃延伸1 min,35个循环;72 ℃终延伸7 min,4 ℃保存;将PCR扩增产物用1.2%琼脂糖检测,符合预期片段的PCR产物送安徽通用生物系统有限公司进行测序,测序结果采用DNAStar的Seq Man进行比对分析,筛选SNP位点。

表1 PPP3CA基因各外显子引物
1.2.3PPP3CA基因二级和三级结构预测 采用RNAfold WebServer (http://rna.tbi.univie.ac.at/ cgi-bin/RNAWebSuite/RNAfold.cgi)预测PPP3CA蛋白突变前后的二级结构;通过Structural Bioinformatics Group(http://www.sbg.bio.ic.ac.uk/~phyre2/ html/page.cgi?id=index)对PPP3CA蛋白在外显子7 处C29T、T50C和T63G位点的CC、TT、AA型和TT、CC、GG型序列,以及外显子16处 G720A位点的GG和AA型序列的三级结构进行预测。
1.2.4PPP3CA基因型频率、等位基因频率及遗传多样性 采用POPGEN(Version 1.31)计算基因型频率、等位基因频率、卡方值(χ2)、杂合度(He)、群体纯合度(Ho)、有效等位基因数(Ne)和多态信息量(PIC)。
基因型频率=该基因个体数/该群体总数
等位基因频率=群体中该基因总数/群体中该对等位基因总数

He=2×该对等位基因频率的乘积
Ho=1-He
式中,Oi为i水平观测频数,Ei为i水平期望频数,n为总频数,pi为i水平期望频率,k为单元格数,∑为总和;Pi、Pj表示群体中第i个和第j个等位基因频率,m为等位基因数。
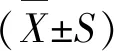
2 结果与分析
2.1 PCR扩增结果
从图1看出,PPP3CA基因外显子扩增产物经1.2%琼脂糖凝胶电泳检测,采用标准型DNA marker 2000条带,分别在368、305、359、302、448及475 bp处均有清晰条带,表明扩增效果较好,结果与预期片段的大小一致,且无引物二聚体。

M,DNA marker 2000;1~3,PPP3CA-Exon7-C29T/T50C/T63G;4~6,PPP3CA-Exon16-G720A;7~9,PPP3CA-Exon8;10~12,PPP3CA-Exon11;13~15,PPP3CA-Exon13;16~18,PPP3CA-Exon15。
2.2 测序结果
从图2看出,在PPP3CA基因外显子7处发现3个SNP位点,外显子16处发现1个SNP位点,分别命名(均以各外显子第1个碱基计数)为C29T、T50C、T63G和G720A,均为错义突变。其中,C29T位点碱基突变,导致第10位氨基酸由苏氨酸突变为蛋氨酸,有3种基因型,分别为CC、CT和TT;T50C位点碱基突变,第17位氨基酸由苯丙氨酸突变为丝氨酸,有2种基因型,分别为TT和CC;T63G位点碱基突变,导致第21位氨基酸由组氨酸突变为谷氨酰胺,有2种基因型,分别为TT和GG;G720A位点碱基突变,第231位氨基酸由甘氨酸突变为丝氨酸,有3种基因型,分别为GG、GA和AA。
2.3 PPP3CA基因的二级和三级结构预测
2.3.1 二级结构 对比PPP3CA蛋白突变前后的二级结构(图3)可以看出,突变前PPP3CA蛋白在外显子7处C29T、T50C、T63G位点的CC、TT、TT型自由能为-29.75 kcal/mol,蛋白质二级结构发生突变表现为C29T、T50C、T63G位点碱基突变,突变后TT、CC、GG型的自由能为-33.33 kcal/mol,自由能降低,稳定性升高;而在外显子16处G720A位点的GG型自由能为-200.52 kcal/mol、AA型自由能为-198.85 kcal/mol,自由能升高,其蛋白质二级结构稳定性降低。

图3 PPP3CA蛋白二级结构的预测图谱

图4 PPP3CA蛋白三级结构的预测图谱

表2 PPP3CA基因外显子7和外显子16多态位点的等位基因频率及基因型频率
2.3.2 三级结构 从图4看出,PPP3CA蛋白在外显子7处C29T、T50C和T63G位点突变,编码的氨基酸发生突变,导致其三级结构中折叠、转折、螺旋等方式发生改变,因此其三级结构也发生改变;而外显子16处G720A位点突变,其231位甘氨酸突变为丝氨酸,但其三级结构未发生改变,表明甘氨酸和丝氨酸在该段三级结构中的作用相似。
2.4 PPP3CA基因的基因型频率、等位基因频率及遗传多样性
测序结果对比发现,仅在赤水乌骨鸡PPP3CA基因外显子7和外显子16发现4个SNPs,分别为C29T、T50C、T63G和G720A,其余外显子均未发现多态位点,将4个SNPs对应的纯合子和杂合子基因型分别定义为CC、CT、TT、TC、TG、GG、GA和AA,分别对几个位点对应的基因型频率、等位基因频率等分析(表2)可知,在C29T突变位点,CC型频率为0.0377、CT型频率为0.1272、TT型频率为0.8050,其中等位基因C在群体中频率为0.1163、T在群体中频率为0.8837,群体纯合度(Ho)为0.7945,杂合度(He)为0.2055,有效等位基因数(Ne)为1.26,多态信息含量(PIC)为0.18(PIC<0.25),属于低度多态;在G720A突变位点,GG型频率为0.1950、GA型频率为0.2013、AA型频率为0.6037,其中等位基因G在群体中频率为0.2957、A在群体中频率为0.7043,群体纯合度(Ho)为0.5835、杂合度(He)为0.4165,有效等位基因数(Ne)为1.71,多态性信息含量(PIC)为0.3298(0.25<0.3298<0.50),属于中度多态;而在T50C、T63G突变位点中出现2种基因型,为T50C位点TT、CC型,T63G位点TT、GG型,其基因型频率、等位基因T、C、T、G频率分别为0.3082、0.6918、0.4591、0.5409;χ2检验结果表明,CC、CT、TT和GG基因座均偏离Hardy-Weinberg平衡状态。
2.5 PPP3CA基因多态性与生长性状的相关性
由表3可知,赤水乌骨鸡PPP3CA基因在外显子7和外显子16处发生碱基突变,与生长性状关联分析发现,外显子7的C29T位点突变,分别有CC、CT和TT共3种基因型,CC型个体的体重和骨盆宽显著高于CT和TT型个体,CC型个体的胸围显著高于TT型个体;外显子7的T50C位点突变,分别有TT和CC 2种基因型,CC型个体的体重、胫长和龙骨长显著高于TT型个体;外显子7的T63G位点突变,分别有TT和GG 2种基因型,TT型个体的胫长和龙骨长显著高于GG型个体;外显子16的G720A位点突变,分别有GG、GA和AA基因型,AA型个体的胫长和龙骨长极显著高于GA型个体,AA型个体的体斜长显著高于GA型个体。表明,外显子7的C29T位点的TT型、T50C位点的CC型、T63G位点的TT型和外显子16 G720A位点的AA型均为优势基因型,能够提高赤水乌骨鸡的生长性状。

表3 PPP3CA基因多态位点与赤水乌骨鸡生长性状关联分析
3 讨 论
骨骼肌在动物生长中有着不可替代的作用。PPP3CA基因能够抑制成纤维细胞生长因子23的表达,与IGFⅠ共同参与骨骼肌生长发育的调节,骨骼肌的层次从上到下依次为肌肉、纤维束、纤维、肌原纤维和肌节[17]。骨骼肌是横纹肌中附着在骨骼上的肌肉,而肌节是含有肌球蛋白II细丝(粗)和肌动蛋白细丝(细),其均参与调节骨骼肌的收缩[8-9]。同时,骨骼肌在生理过程中也发挥重要作用,骨骼肌干细胞具有再生功能,对骨骼肌的发育、生长、维持和修复至关重要[20]。PPP3CA基因在控制骨骼肌纤维类型中起关键作用,其调节骨骼肌的生长发育,与生长因子VEGF和FGF2共同调节细胞的增殖和信号传导[9];在成肌细胞中,PPP3CA基因与PPARGC1A基因共同正向调节成肌细胞生长发育,当抑制PPARGC1A基因的表达时,PPP3CA基因的表达受到显著抑制,将抑制肌肉的生长发育,表明PPP3CA和PPARGC1A基因对成肌细胞生长发育具有协同作用[21]。单艳菊等[22]研究发现,在清远麻鸡1周龄时趾长伸肌和3周龄时腓肠肌外侧头、比目鱼肌出现急速上升,且清远麻鸡3种类型骨骼肌中PPP3CA基因mRNA的表达量均高于隐性白羽肉鸡,在3周龄时呈极显著差异。近年来的文献报道表明,PPP3CA基因对动物的生长发育和肉质具有重要影响,如PPP3CA基因已被证实参与牛肌肉前脂肪细胞的分化[23]。王兴群等[24]发现,江口萝卜猪PPP3CA基因G316451A位点突变,且GG和GA型的大理石纹评分显著高于AA型,表明PPP3CA基因对江口萝卜猪肉质存在明显影响。Chen等[11]研究表明,PPP3CA基因对猪肌肉的生长有重要影响。包艳波[25]研究发现,PPP3CA基因的AA型对大骨鸡母鸡肌肉的锁水率、腿肌熟肉率、胸腿肌pH及胸肌灰分含量影响显著,提示该基因型有可能作为筛选优良肉质性状的候选型。本试验通过对赤水乌骨鸡PPP3CA基因外显子多态性筛选,在外显子7处发现的3个错义突变位点C29T、T50C和T63G,在外显子16处发现1个错义突变位点G720A。基因分型并与生长性状关联分析发现,外显子7 处C29T位点突变的CC型显著提高个体的体重、骨盆宽和胸围;T50C位点突变的CC型显著提高个体的体重、胫长和龙骨长;T63G位点突变的TT型显著提高个体的胫长和龙骨长;外显子16处G720A位点突变的AA型极显著提高个体的胫长和龙骨长,GG型显著提高个体的体斜长。由此看出,C29T的CC型、T50C的CC型、T63G的TT型和G720A的AA型均为优势基因型,但C29T和T50C位点碱基突变为不利突变,而T63G和G720A位点的碱基突变为有利突变,且G720A位点更有利于赤水乌骨鸡生长。说明,PPP3CA基因对赤水乌骨鸡的生长发育具有一定的影响。
4 结 论
在PPP3CA基因外显于7和外显子16处其发现4个错义突变位点,其中,T63G和G720A位点的碱基突变为有利突变,且G720A位点更有利于赤水乌骨鸡生长,可作为赤水乌骨鸡分子标记辅助选育参考位点。本研究首次探讨PPP3CA基因与鸡生长性状的相关性,并在家鸡的PPP3CA基因外显子中发现对赤水乌骨鸡生长性状具有一定影响的多态位点,可作为赤水乌骨鸡分子标记辅助选育参考位点。研究结果可为今后赤水乌骨鸡的育种目标提供参考依据,并为进一步研究鸡PPP3CA基因奠定基础。
猜你喜欢
电子科技大学学报(2022年5期)2022-10-29
中国生殖健康(2020年4期)2021-01-18
中国畜禽种业(2020年7期)2020-12-16
乡村地理(2019年2期)2019-11-16
江苏农业科学(2018年21期)2018-12-06
中国生殖健康(2018年4期)2018-11-06
乡村地理(2018年3期)2018-11-06
乡村地理(2018年4期)2018-03-23
艺术评鉴(2016年6期)2016-05-14
中国畜牧兽医文摘(2016年7期)2016-01-30
