
血管内皮生长因子-A165对形觉剥夺性近视豚鼠巩膜重塑的影响
2023-09-12 02:36彭庆生高洪莲孙瑞婷张凤一
国际眼科杂志 2023年9期
彭庆生,高洪莲,孙瑞婷,张凤一,王 磊,李 童,张 磊
0 引言
近视是全世界主要的公共卫生问题之一[1]。全球约有26.20亿人患有近视,3.99亿人患有高度近视,据估计到2050年,全球将有近47.58亿近视患者,其中高度近视患者达到9.38亿[2]。近视,尤其是高度近视不仅会导致模糊的视觉,还将极大地增加眼部病变的风险,引发脉络膜新生血管、视网膜萎缩、视网膜脱离和视神经病变等疾病[3-4]。目前高度近视已成为全球人口视力损伤的主要原因之一[5]。因此,采取有效的防控措施对于预防近视发生和减少向高度近视进展至关重要。
目前研究表明,眼轴增长是引发近视的主要原因[6],而巩膜重塑变薄与眼轴延长显著相关,其不仅是眼轴增长的主要机制,还是近视发生时的重要改变[7-8],因此巩膜重塑可能是控制近视的关键靶点。而巩膜重塑主要依赖于其细胞外基质成分的改变,受多种因子调控[9],主要包括基质金属蛋白酶-2(matrix metalloproteinase-2,MMP-2)、基质金属蛋白酶抑制剂-2(tissue inhibitor of matrix metalloproteinase-2,TIMP-2)、转化生长因子(transforming growth factor,TGF)-β及α-平滑肌肌动蛋白(α-smooth muscle actin,α-SMA)等重要因子。视网膜中的多巴胺(dopamine,DA)是一种重要的神经递质,在视觉信息传递和屈光发育调节中发挥关键作用[10-11],其活性改变可以通过视网膜-巩膜级联机制影响巩膜重塑,进而调节眼球的生长发育,被认为是近视的终止信号[12-13]。而血管内皮生长因子(vascular endothelial growth factor,VEGF)-A165具有直接和间接的神经营养和神经保护作用[14],可以保护视网膜神经元[15],体内外均能促进DA神经元的存活和功能恢复[16-17],故推测其在玻璃体腔内注射后可以作用于视网膜神经元,调节视网膜中DA的活性,进而影响巩膜重塑。本研究通过遮盖豚鼠右眼建立形觉剥夺性近视(form-deprivation myopia,FDM)模型,探讨玻璃体腔内注射VEGF-A165对FDM豚鼠巩膜重塑的影响及其对近视的干预作用,从而为近视防控提供新的思路。
1 材料和方法
1.1材料
1.1.1实验动物及分组选取健康3周龄英国短毛三色豚鼠120只(济南金丰实验动物有限公司),雌雄不限,体质量150~200g。采用随机数字表法将豚鼠分为空白组、FDM组、PBS组、1ng组、5ng组、10ng组,每组20只。自然照明环境下12h/12h昼夜循环,自由摄水摄食,维持温度为22℃~25℃,湿度为50%~70%。本研究通过滨州医学院附属医院伦理委员会审批(批准号:20220128-77),实验动物的使用严格遵循滨州医学院动物管理委员会的相关规定。
1.1.2主要试剂与仪器主要试剂:重组人蛋白VEGF-A165(美国R&D Systems公司);复方托吡卡胺滴眼液(参天制药中国有限公司);组织RNA快速提取试剂盒(艾科瑞生物工程有限公司);兔抗鼠MMP-2(A6247)、TIMP-2(A16439)、TGF-β1(A16640)、TGF-β2(A3640)、α-SMA(A1011)多克隆抗体(中国ABclonal股份有限公司);BCA蛋白浓度测定试剂盒(北京碧云天生物科技有限公司)。主要仪器:带状光检影镜(苏州六六视觉科技有限公司);A型超声(法国Quantel公司);高效液相色谱仪(美国Thermo Fisher公司)。
1.2方法
1.2.1动物模型建立空白组豚鼠不做任何干预;FDM组、PBS组、1ng组、5ng组、10ng组豚鼠均用修剪出左眼、双耳和口鼻的半透明无毒气球遮盖右眼处理,分笼饲养,每天光照与黑暗周期为12h/12h,连续遮盖14d建立形觉剥夺性近视豚鼠模型。
1.2.2药物干预遮盖前根据分组在PBS组、1ng组、5ng组、10ng组豚鼠右眼玻璃体腔内分别注射PBS缓冲液2.5μL、重组人蛋白VEGF-A1651、5、10ng[16-17],其中VEGF-A165均溶于2.5μL的PBS缓冲液中。玻璃体腔注射前1d使用妥布霉素滴眼液滴右眼。舒泰肌肉注射麻醉,聚维酮碘消毒豚鼠结膜囊及眼表后用生理盐水冲洗干净,显微镜下用眼科镊固定眼球,注射器垂直巩膜进针,避开晶状体推进约2~3mm后注射,停留10s后用眼科镊轻轻夹住针孔辅助拔出针头,观察有无液体流出、结膜有无红肿,氧氟沙星眼膏涂眼,半透明无毒气球遮眼。术后3d内观察右眼有无炎症,氧氟沙星眼膏涂眼(每日3次)。
1.2.3眼部生物学参数测量造模前后分别测量各组豚鼠右眼的屈光度和眼轴长度。测量屈光度前,用复方托吡卡胺滴眼液滴右眼散瞳,待瞳孔充分散大后使用带状光检影镜测量屈光度,由经验丰富的检影师验光,根据工作距离以等效球镜度(球镜度+1/2柱镜度)进行数据分析,连续测量5次计算平均值作为该眼的屈光度。测量眼轴长度前,用盐酸丙美卡因滴眼液行角膜表面麻醉,待药物生效后应用眼部A超测量眼轴长度,选择手动模式连续测量5次计算平均值,精确到0.01mm。
1.2.4标本收集造模结束后,注射凝血剂处死豚鼠,摘取右眼眼球,于冰上快速取其视网膜和后极部巩膜,保存于-80℃冰箱。
1.2.5高效液相色谱法检测视网膜中DA含量取各组豚鼠视网膜称重,每mg组织样本加入新鲜配制的匀浆液20μL(H3ClO40.1mol/L,EDTA Na20.1mmo/L,内标物DHBA),-40℃冷冻匀浆(2mm氧化锆珠,60Hz,30s,4次);低温离心(20000r/min,30min,4℃)后取上清保存于-80℃冰箱。测样前取出再次离心(20000r/min,30min,4℃)后取上清液上机检测。色谱柱:Thermo Acclaim Rapid Separation Liquid Chromatography (RSLC) 2.1*100mm C182.2μm,柱温40℃;流动相:含NaH2PO490mmo/L,柠檬酸50mmo/L,OSA 1.7mmo/L,EDTA 50μmol/L和乙腈4.5%。所有数据均由Chromeleon 6.9色谱工作站采集与分析,以内标法计算目标物浓度。
1.2.6RT-PCR法检测巩膜中MMP-2、TIMP-2、TGF-β2、TGF-β1、α-SMAmRNA的表达取各组豚鼠巩膜,在低温研磨仪中充分研磨后,根据试剂盒说明书中的纯化步骤提取巩膜总RNA,随后将RNA反转录成cDNA,反应程序:42℃孵育2min,37℃孵育15min,85℃加热5s,以4℃降温结束。以GAPDH为内参(引物序列见表1),进行PCR扩增,反应程序:95℃预变性1min,95℃变性10s,60℃延伸30s,循环40次,记录Ct值,使用2-ΔΔCt法计算各目的基因相对表达量。

表1 引物序列
1.2.7Westernblot法检测巩膜中MMP-2、TIMP-2、TGF-β2、TGF-β1、α-SMA蛋白的表达根据分组将豚鼠巩膜置于不同的研磨管中,加入裂解液和蛋白酶抑制剂后于低温研磨仪中研磨5min,转移至EP管,置于4℃中静置30min,低温离心30min(4℃,12000r/min),取上清液于新的EP管中,采用BCA浓度测定试剂盒测定蛋白浓度,剩余上清液中加入5×loading buffer,恒温金属浴中98℃煮5~10min,蛋白样品经电泳、转膜后,室温条件下加入5% BSA封闭2h,加入对应的一抗稀释液(抗体的稀释方法参照其说明书选取稀释溶液与最佳浓度),于4℃冰室摇床上孵育过夜,TBST冲洗3次×15min,室温条件下加入二抗孵育1h,TBST冲洗4次×15min,化学发光法显影,Image J软件分析灰度值。
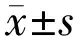
2 结果
2.1眼部生物学参数造模前,各组豚鼠右眼均呈现远视状态,各组豚鼠右眼屈光度、眼轴长度差异均无统计学意义(P>0.05)。造模14d后,与空白组相比,FDM组豚鼠右眼屈光度变为负值,眼轴显著延长,差异均有统计学意义(P<0.01);与FDM组相比,PBS组豚鼠右眼屈光度、眼轴长度未发生明显变化(均P>0.05),而1ng组、5ng组、10ng组豚鼠右眼向近视偏移程度均减轻,眼轴长度均较短(均P<0.01),其中1ng组豚鼠右眼近视偏移程度和眼轴长度均低于5ng组和10ng组,5ng组豚鼠右眼屈光度和眼轴长度均低于10ng组,三组间两两比较差异均有统计学意义(P<0.05),见表2。

表2 各组豚鼠造模前后右眼屈光度和眼轴长度比较
2.2高效液相色谱法检测视网膜中DA含量造模14d后,空白组、FDM组、PBS组、1ng组、5ng组、10ng组豚鼠右眼视网膜中DA含量分别为108.737±8.297、58.623±6.547、61.383±7.900、98.202±6.170、84.535±11.470、72.275±6.863ng/g,差异有统计学意义(F=37.555,P<0.01)。与空白组相比,FDM组豚鼠右眼视网膜中DA含量明显降低,差异有统计学意义(P<0.01);与FDM组相比,PBS组豚鼠右眼视网膜中DA含量未发生明显变化(P>0.05),而1ng组、5ng组、10ng组豚鼠右眼视网膜中DA含量均增加(均P<0.01),其中1ng组增加较5ng组明显,5ng组增加较10ng组明显,三组间两两比较差异均有统计学意义(P<0.05)。
2.3RT-PCR法检测巩膜中相关基因的表达造模14d后,与空白组相比,FDM组豚鼠右眼巩膜中MMP-2、TGF-β2、α-SMA mRNA相对表达量均增加,TGF-β1、TIMP-2 mRNA相对表达量均减少,差异均有统计学意义(P<0.01);与FDM组相比,PBS组豚鼠右眼巩膜中各目的基因的相对表达量未发生明显变化(均P>0.05),而1ng组、5ng组、10ng组豚鼠右眼巩膜中MMP-2、TGF-β2、α-SMA mRNA相对表达量均减少,TGF-β1、TIMP-2 mRNA相对表达量均增加(均P<0.01),其中1ng组豚鼠右眼巩膜中MMP-2、TGF-β2、α-SMA mRNA相对表达量均低于5ng组和10ng组,TGF-β1、TIMP-2 mRNA相对表达量均高于5ng组和10ng组(均P<0.01),5ng组豚鼠右眼巩膜中TGF-β2、α-SMA mRNA相对表达量均低于10ng组,TGF-β1、TIMP-2 mRNA相对表达量均高于10ng组(均P<0.01), 但5ng组和10ng组豚鼠右眼巩膜中MMP-2 mRNA相对表达量差异无统计学意义(P=0.353),见表3。
表3 造模后各组豚鼠巩膜中相关基因的表达
2.4Westernblot法检测巩膜中相关蛋白的表达造模14d后,与空白组相比,FDM组豚鼠右眼巩膜中MMP-2、TGF-β2、α-SMA蛋白相对表达量均增加,TIMP-2、TGF-β1蛋白相对表达量均减少,差异均有统计学意义(P<0.01);与FDM组相比,PBS组豚鼠右眼巩膜中各蛋白相对表达量未发生明显变化(均P>0.05),而1ng组、5ng组、10ng组豚鼠右眼巩膜中MMP-2、TGF-β2、α-SMA蛋白相对表达量均减少,TIMP-2、TGF-β1蛋白相对表达量均增加(均P<0.01),其中1ng组豚鼠右眼巩膜中MMP-2、TGF-β2、α-SMA蛋白相对表达量均低于5ng组和10ng组,TGF-β1、TIMP-2蛋白相对表达量均高于5ng组和10ng组(均P<0.01),5ng组豚鼠右眼巩膜中TGF-β2、α-SMA蛋白相对表达量均低于10ng组,TGF-β1、TIMP-2蛋白相对表达量均高于10ng组(均P<0.05), 但5ng组和10ng组豚鼠右眼巩膜中MMP-2蛋白相对表达量差异无统计学意义(P=0.610),见图1。
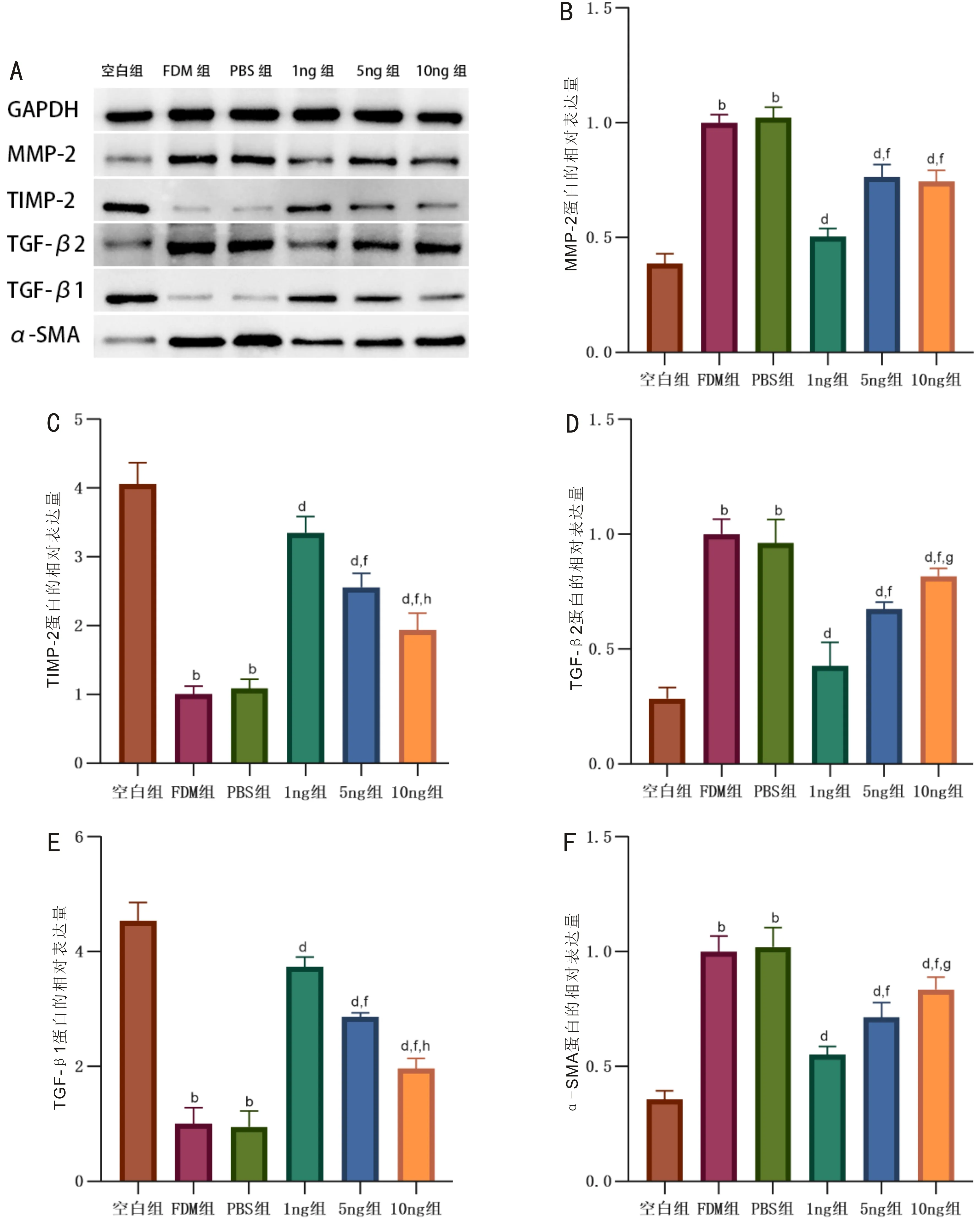
图1 造模后各组豚鼠巩膜中相关蛋白的表达 A:Western Blot电泳图;B:MMP-2蛋白相对表达量(F=99.230,P<0.01);C:TIMP-2蛋白相对表达量(F=94.752,P<0.01);D:TGF-β2蛋白相对表达量(F=49.873,P<0.01);E:TGF-β1蛋白相对表达量(F=121.672,P<0.01);F:α-SMA蛋白相对表达量(F=56.915,P<0.01)。bP<0.01 vs 空白组;dP<0.01 vs FDM组;fP<0.01 vs 1ng组;gP<0.05,hP<0.01 vs 5ng组。
3 讨论
近年来,近视的病因及机制一直是眼科研究的热点,尽管具体的机制目前仍不清楚,但多数研究认为眼轴长度的异常增加是引发近视的主要原因,巩膜细胞外基质重塑是眼轴增长的主要机制[7-8,18]。当巩膜重塑发生时,其细胞外基质中胶原降解增加、蛋白聚糖合成减少、成纤维细胞向肌成纤维细胞转化,巩膜原纤维组装紊乱,巩膜生物力学变弱,巩膜逐渐变薄和延伸,眼轴随之延长,近视也逐渐发展[9,19]。由此可见,巩膜重塑在近视的发生和发展中起着关键作用,可能是近视机制的一个关键靶点。
巩膜重塑是一个受多因素调控的动态过程,其中局部缺氧是该过程的主要诱因。研究表明,缺氧可以调控巩膜重塑相关因子的活动,引起细胞外基质中胶原含量的下降和肌成纤维细胞的转分化,最终导致巩膜重塑[20-21]。同时,MMP-2在该过程中也发挥重要作用,其含量的增加不仅可使巩膜中Ⅰ型胶原的降解增加,使胶原纤维逐渐变细,引起巩膜重塑,近视的启动也依赖于MMP-2的表达增加[22-23];而TIMP-2是MMP-2的内源性抑制因子,可使巩膜中胶原的降解减少,两者之间的失衡可引起巩膜细胞外基质代谢异常,胶原蛋白水平下降,从而导致细胞外基质重塑[24-25]。α-SMA在巩膜重塑中也发挥关键作用,其具有细胞收缩特性,可以促进近视发生时巩膜的变薄和延伸,同时α-SMA表达量的增加标志着肌成纤维细胞的转分化和细胞应力的增加[26-27]。TGF-β在调节细胞外基质成分的转换中发挥重要作用,不仅可以调节成纤维细胞的转化和胶原蛋白的含量,还可以特异性调节巩膜细胞的收缩特性,影响α-SMA的表达[26],其中TGF-β1不仅可以促进巩膜成纤维细胞的增殖,还可调控Ⅰ型胶原的合成,增加基质中胶原的含量[28],而TGF-β2主要调节基质中成纤维细胞的增殖[29],可以呈剂量依赖性地促进大鼠巩膜中成纤维细胞的增殖,使巩膜增厚[30]。综上所述,MMP-2、TIMP-2、TGF-β1、TGF-β2、α-SMA在调控巩膜细胞外基质重塑中发挥关键作用,因此通过测定上述指标的表达,可以有效观察巩膜重塑情况。现已有大量研究表明,随着近视的发生,巩膜中MMP-2、TGF-β2、α-SMA的表达增加[9,26],TIMP-2、TGF-β1的表达降低[24,28],这与本研究中FDM组豚鼠右眼巩膜中各种因子的表达一致,均代表着近视性巩膜重塑的建立。
视网膜中的DA是传递视觉信息的视网膜-巩膜信号级联的起点,与眼球的屈光发育息息相关[13]。研究表明,DA活性的改变可以影响视网膜与其色素上皮层中的信号通路,进而影响脉络膜和巩膜之间的信息传递,最终通过巩膜的细胞外基质重塑调节眼睛的生长发育[12,20]。同时,VEGF已经被证明是一种潜在的治疗中枢神经系统疾病的药物,可以通过营养DA神经元,增加神经递质DA的分泌[17,31],其中VEGF-A165是主要的同种型,在海马神经节、背根神经节和视网膜神经元中均具有神经保护作用,对神经元具有直接的抗细胞毒性作用[15-16]。所以我们推测VEGF-A165在眼内可以增加视网膜中DA的活性,对巩膜重塑产生影响,从而干扰近视发展。本课题组前期研究发现,玻璃体腔内注射抗VEGF药物(康柏西普)会降低FDM豚鼠视网膜中DA含量,促进近视发展[32],间接证明了VEGF对视网膜中DA含量的影响。
本研究通过玻璃体腔内注射VEGF-A165,验证其对FDM豚鼠巩膜重塑的影响,以及对FDM的干扰效果。Beazley-Long等[16]研究发现,玻璃体腔内注射10ng VEGF-A165对大鼠视网膜神经元具有神经保护作用,可减少视网膜神经节细胞和内核层细胞凋亡;同时,Meng等[17]研究提出,VEGF对DA神经元的保护作用具有剂量依赖性,在1ng/mL组和10ng/mL组中,1ng/mL组的神经保护作用更强。因此,本研究采用了3种不同剂量(1、5、10ng)进行注射,探究不同剂量VEGF对FDM豚鼠巩膜重塑的作用是否也会有差异。结果显示,与FDM组相比,VEGF-A165处理组豚鼠视网膜中DA含量均增加,这与Sheikh等[31]研究结果一致,该研究表明VEGF对DA神经元具有保护作用,本研究进一步证明了这种神经保护可以作用于视网膜DA神经元,增加视网膜中DA的含量。同时,本研究发现,1ng组、5ng组、10ng组豚鼠近视程度均降低,眼轴增长趋势明显减缓,巩膜中MMP-2、TGF-β2、α-SMA表达量均减少,TIMP-2、TGF-β1表达量均增加,而Yu等[9]和Jobling等[26]研究认为,近视恢复期豚鼠巩膜中MMP-2、TGF-β2、α-SMA表达的降低以及TIMP-2、TGF-β1表达的升高,代表着基质中胶原含量的升高、巩膜厚度的增加等,这些均提示近视性巩膜重塑受到抑制。因此,本研究结果表明VEGF-A165可以增加视网膜中DA含量,对巩膜重塑产生抑制作用,抑制近视进展。Lin等[33]研究表明,玻璃体腔内注射DA主要通过调节巩膜发育抑制近视发展,而本研究结果提示巩膜细胞外基质的增加会抑制近视的发展,间接证明了视网膜与巩膜之间的级联信号在近视发生发展中的作用,同时还提出了一种可以提高视网膜中DA浓度的方法,即玻璃体腔内注射VEGF-A165。
此外,本研究发现,随着VEGF-A165剂量的增加,近视偏移程度加重,视网膜中DA含量逐渐减少,巩膜中MMP-2、TGF-β2、α-SMA表达量均逐渐增加,TIMP-2、TGF-β1表达量均逐渐减少,但巩膜中MMP-2在5ng组与10ng组中的表达无显著差异。分析其原因是由于巩膜缺氧时缺氧诱导因子-1α(hypoxia-inducible factor-1α,HIF-1α)主要被激活[21],引起巩膜重塑相关因子的改变,而缺氧时MMP-2的上调依赖于HIF-2α的表达而不是HIF-1α的表达[34],且房水中MMP-2浓度达到一定范围时与VEGF-A呈负相关[35],但具体机制还需进一步探讨。本研究中,增加VEGF-A165的剂量反而会减弱对豚鼠巩膜重塑的抑制作用,其中1ng剂量的抑制效果最好。推测其可能的机制主要有以下两种:(1)视网膜中DA含量的降低导致了这种结果。由于DA主要通过影响巩膜的发育和重塑调控近视的发生发展[12],且降低视网膜中DA的含量会促进近视发展[32],因此本研究中玻璃体腔注射VEGF-A165剂量增加引起视网膜中DA含量相对降低,可能通过级联信号减弱对巩膜重塑的抑制作用,从而发挥对近视的促进作用;而视网膜中DA含量由1ng组到10ng组逐渐减少的原因,可能是由于VEGF-A165对DA神经元的作用在低剂量下获益最大[17]导致的;(2)VEGF-A165过量可能透过血-视网膜屏障直接作用于脉络膜[36],诱导病理性血管生成,使脉络膜毛细血管被血栓阻塞[37],进而减少了脉络膜血流灌注,加重了巩膜的缺氧[20],从而相对性地促进了5ng组与10ng组豚鼠巩膜细胞外基质的重塑。但对于以上两种推测的验证,还需通过对VEGF-A165作用机制的进一步研究阐明。
本研究通过在FDM豚鼠玻璃体腔内注射不同剂量的VEGF-A165,发现均会增加视网膜中DA含量,对豚鼠巩膜重塑起到抑制作用,抑制FDM进展。其机制可能是VEGF-A165通过营养和保护视网膜DA神经元,增加视网膜中DA的活性,进而通过信号级联抑制巩膜重塑,从而发挥对近视的抑制作用,但具体的发生机制还需进一步实验研究。同时本研究发现,1、5、10ng VEGF-A165对巩膜重塑的抑制作用存在差异,其中1ng效果最好,5ng次之,10ng最差。本文不足之处在于仅观察了3种剂量VEGF-A165作用的差异,对于更小或更大浓度梯度是否还会存在差异,以及0.5ng或2ng是否会有比1ng具有更好的效果等问题尚不明确,因此还需在此研究基础上进一步探讨VEGF-A165作用的最佳剂量范围,准确把握剂量差异对个体造成的影响,从而为以后VEGF-A165用于临床近视防控提供实验基础。
猜你喜欢
人生与伴侣·共同关注(2022年2期)2022-03-14
检验医学与临床(2020年20期)2020-11-02
文萃报·周二版(2020年25期)2020-07-06
广东医科大学学报(2020年6期)2020-02-06
小读者(2019年24期)2020-01-19
阅读(低年级)(2019年6期)2019-08-27
国际眼科杂志(2019年7期)2019-07-25
小小说大世界(2019年3期)2019-07-02
中华老年多器官疾病杂志(2016年9期)2016-04-28
军事文摘(2015年6期)2015-06-16
