
黄土高原蓝藻和藓类生物结皮对草本植物多样性及生物量的影响
2023-09-11 07:35:24王彦峰汪万福余星兴
生态学报 2023年16期
王彦峰,肖 波,汪万福,余星兴,张 雪
1 中国科学院西北生态环境资源研究院, 沙漠与沙漠化重点实验室, 兰州 730000 2 中国科学院水利部水土保持研究所黄土高原土壤侵蚀与旱地农业国家重点实验室, 杨凌 712100 3 中国科学院大学,北京 100049
生物结皮是干旱和半干旱气候区常见的地表覆盖物,其盖度最高可达60%—70%[1],是由隐花植物(藻类、地衣、苔藓)、土壤微生物(细菌、真菌和古生菌)以及其他微小生命体通过菌丝体、假根和多聚糖等分泌物与土壤表层颗粒胶结形成的复合体[2—3]。生物结皮介导了干旱和半干旱气候区土壤边界层物质与能量的输入、转运和输出,被喻为“荒漠生态工程师”[4—5]。作为先锋物种,生物结皮发育改变了土壤边界层结构、水分循环过程、土壤温度以及土壤养分循环,从而影响草本植物的种子传播、萌发和幼苗的建植与生长,进而影响草本植物的群落结构、物种丰度和生物量[6]。同时,有研究认为,生物结皮良好的抗逆性可缓冲气候变化的负面效应,为维管束植物提供庇佑,有利于维系区域生态系统的稳定与生态安全[7]。因此,研究生物结皮对草本植物群落的影响,对于明晰干旱和半干旱气候区草本植物的建群机制及其对生态功能的影响具有重要意义,同时对明晰植物群落结构与系统功能对气候变化的响应机制具有重要意义。
生物结皮与草本植物镶嵌形成斑块式分布格局[8],通过影响草本植物的土壤种子库、种子萌发、幼苗建植等过程来影响草本植物的多样性[9—10]。国内外有关生物结皮对草本植物多样性影响的研究已有很多。其中,一些研究认为,生物结皮增加了土壤表面粗糙度,有利于土壤种子着床[11],同时生物结皮为幼苗的发育提供了稳定的土壤环境和必要的水分和养分支持[12—13],从而促进了草本植物多样性[14—15]。例如,陈孟晨等[16]在沙坡头的研究显示,地衣结皮和藓结皮显著增加了土壤种子库丰度;Muoz等[17]的研究认为,蓝藻结皮发育有利于Sennanotabilis和Acaciahilliana种子的萌发;张元明等[18]在准噶尔盆地的研究认为,地衣结皮使角果藜(Ceratocarpusarenarius)的生长速率提升了21%。但也有研究认为,生物结皮在地表形成机械屏障,减少了种子和土壤的接触机率,不利于种子萌发和出苗[6,19],从而降低了草本植物的多样性。Song等[20]在腾格里沙漠、Li等[21]在高寒草甸以及Guan等[22]在毛乌素沙地的研究都证实了生物结皮对种子萌发有抑制作用。综上,目前关于生物结皮对草本植物多样性影响的研究仍存在较大争议,且相关研究多局限于风沙土区,而关于黄绵土区的研究还较少。因此,生物结皮对草本植物多样性影响的机理尚不明晰,其影响程度和作用途径仍有待进一步研究。
本研究在黄土高原的风沙土和黄绵土上分别选取蓝藻结皮和藓结皮样地,以无结皮为对照,通过样方法调查了两种土壤上生物结皮和无结皮的群落组成差异,测定并计算了其草本植物的多样性指数和生物量,继而结合灰色关联分析和结构方程模型,剖析了生物结皮对草本植物多样性和生物量的影响程度和作用途径。研究结果旨在明晰黄土高原生物结皮对草本植物多样性和生物量的影响程度和作用途径,为进一步理解干旱和半干旱气候区草本植物的建群机制和生态系统功能提供科学依据。
1 材料与方法
1.1 研究区概况
研究区位于黄土高原北部的陕西省神木市六道沟流域(38°34′—38°51′N,110°21′—110°23′E),其海拔1094.0—1273.9 m,面积6.89 km2。流域属温带半干旱气候,地形为典型的盖沙黄土丘陵地貌,降雨集中在6—9月,多年平均降雨量为408.5 mm。流域分为东西两部分,西部以风沙土为主,东部以黄绵土为主。退耕还林(草)工程实施后,流域内的生态环境得到明显改善,同时蓝藻结皮和藓结皮在流域内广泛发育,成为该区一种常见地表覆盖物。其中,藓结皮中的优势藓种有真藓(Bryumargenteum)、双色真藓(Bryumbicolor)和尖叶对齿藓(Didymodonconstrictus)等;而蓝藻结皮中的优势藻属为念珠藻属(Nostoc)、色球藻属(Chroococcus)和微鞘藻属(Microcolus)等。
1.2 样地选取与草本植物调查方法
在对流域总体进行调查的基础上,于2021年7月—8月开展样地选取与草本植物群落调查。针对风沙土和黄绵土两种土壤的无结皮、蓝藻结皮和藓结皮6种处理选取样地。所选样地为封禁年限(20年)一致的撂荒草地,样地大小10 m×10 m,同时每种土壤上无结皮蓝藻结皮和藓结皮样地间隔大于20 m。在每种样地随机布设10个1 m×1 m的样方调查草本植物群落,共调查60个样方。
草本植物多样性调查时,先对样方中的草本植物进行照片记录,而后参考《内蒙古植物志》和《中国植物志》进行物种鉴定。草本植物多样性指数的计算方法为:Patrick指数D=S,S为物种数;Shannon指数H′=-∑pilnpi,pi为种i占全部个体的比例[23]。此外,收集样方中所有草本植物的地上部分,于烘箱内85℃杀青30 min,后于65℃烘干至恒重并称重,以此计算单位面积草本植物的生物量。
1.3 土壤理化性质测定
草本植物样品收集前,用土壤水分和温度传感器(5TM,美国METER公司)搭配手持式读表(ProCheck,美国METER公司)测定样方表层5 cm的土壤体积含水率和温度。测定期间天气晴好,测定时间为10:00—11:00。同时,用环刀(100 cm3)取原状土样后,于室内用半自动土壤贯入仪(UTM6102,中国三思纵横科技股份有限公司)测定表层2 cm的土壤穿透阻力。
将植物样品用剪刀齐根剪起后,于纸质档案袋中保存。随后,在样方中按五点法采集0—5 cm的土壤样品,并将同一样方中的5个样品混合后存于自封袋中,于实验室内测定土壤化学性质,共采集60个土壤样品。其中,硝态氮用紫外分光光度法,铵态氮用浸提-靛蓝比色法测定,速效磷用碳酸氢钠浸提-钼锑抗比色法测定,有机质用重铬酸钾-硫酸消化法测定。
1.4 数据处理
使用R语言中vegan包计算草本植物的多样性指数,使用IBM SPSS 22.0 软件的ANOVA和LSD多重比较分析不同样地间草本植物多样性指数、生物量和土壤属性的差异,用OriginPro 2021b中PCA应用程序分析不同样地间草本植物群落结构的差异;同时,用灰色关联分析筛选对草本植物多样性和生物量影响的主要土壤因子,并构建结构方程模型检验生物结皮类型、土壤类型和主要土壤影响因子对草本植物多样性和生物量的影响途径和方式;此外,使用OriginPro 2021b绘制图表。
2 结果与分析
2.1 生物结皮的结构和组成
表1可见,风沙土和黄绵土上,蓝藻结皮和藓结皮的盖度、厚度、叶绿素含量均存在明显差异(P<0.05)。风沙土上,藓结皮的盖度、厚度、叶绿素含量分别是蓝藻结皮的1.3、1.4倍和7.1倍;而黄绵土上,藓结皮的三种基本特性分别是蓝藻结皮的1.1、1.3倍和2.9倍。此外,两种壤质上蓝藻结皮的优势属均为念珠藻属(Nostoc)、色球藻属(Chroococcus)和微鞘藻属(Microcolus);而藓结皮的优势种则分别为尖叶对齿藓(Didymodonconstrictus)和真藓(Bryumargenteum)。

表1 样地中生物结皮的基本特征
2.2 生物结皮对草本植物物种组成的影响
风沙土上,无结皮样地中有草本植物7科15属15种,优势种为狗尾草(Setariaviridis)、猪毛菜(Salsolacollina)和角蒿(Incarvilleasinensis),蓝藻结皮中有6科10属10种,优势种为茵陈蒿(Artemisiacapillaris)、狗尾草和猪毛菜,而藓结皮有8科14属14种,优势种为狗尾草和茵陈蒿(表2);黄绵土上,无结皮样地中有草本植物12科25属27种,优势种为虎尾草(Chlorisvirgata)、铁杆蒿(Artemisiagmelinii)、芨芨草(Achnatherumsplendens)和狗尾草,蓝藻结皮中有7科14属15种,优势种为兴安胡枝子(Lespedezadaurica)、远志(Polygalatenuifolia)和铁杆蒿,而藓结皮有7科14属15种,优势种为三芒草(Aristidaadscensionis)和狗尾草。
风沙土上,无结皮样地的优势科为禾本科(Gramineae,5种)和菊科(Compositae,3种),而蓝藻结皮样地的优势科为藜科(Chenopodiaceae,4种),藓结皮样地的优势科为禾本科(5种);与无结皮相比,风沙土的蓝藻结皮和藓结皮样地中多年生草本植物株数占比分别增加了30%和16%,而多年生草本植物株数则分别减少了30%和16%。黄绵土上,无结皮样地的优势科为菊科(8种)、禾本科(5种)和豆科(Leguminosae,4种),蓝藻结皮样地的优势科同为菊科(4种)、禾本科(4种)和豆科(3种),而藓结皮样地的优势科为禾本科(7种)和菊科(3种);此外,与无结皮相比,黄绵土蓝藻结皮样地中的多年生草本植物株数增加了45%,而藓结皮样地中则减少了46%(图1),这些结果表明,生物结皮改变了草本植物群落的优势科和生活型株数占比。
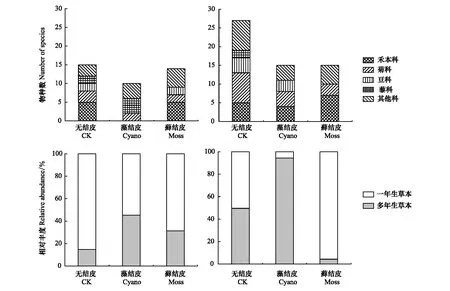
图1 生物结皮与无结皮样地中草本植物群落的优势科和生活型Fig.1 The dominant families and life forms of herbaceous plants from the plots of biocrusts and uncrusted soil
主成分分析结果显示(图2),在风沙土上,PC1和PC2解释了无结皮、蓝藻结皮和藓结皮样地中草本植物群落结构42.2%的总方差,且生物结皮和无结皮样地分聚两类。而黄绵土上,PC1和PC2解释了三种样地中草本植物群落结构47.5%的总方差,且三种样地分聚为三类。可见,生物结皮可能会改变草本植物的群落结构。

图2 生物结皮与无结皮样地中草本植物群落结构的PCA分析Fig.2 The PCA analysis results of herbaceous plant community from the plots of biocrusts and uncrusted soilPCA表示主成分分析 Principal components analysis
2.3 生物结皮对草本植物多样性的影响
风沙土上(图3),蓝藻结皮样地中草本植物的Patrick和Shannon指数分别为7.8和1.3,分别比无结皮降低了11%和16%(P<0.05);而藓结皮样地中草本植物的两种多样性指数分别为5.4和1.1,依次比无结皮降低了39%和30%(P<0.05),表明生物结皮显著降低了草本植物的多样性。同时,藓结皮样地的Patrick和Shannon指数与蓝藻结皮差异显著(P<0.05),显示藓结皮对草本植物多样性的影响大于蓝藻结皮。黄绵土上(图3),无结皮样地中草本植物的两种多样性指数分别为12.1和1.5,分别是蓝藻结皮样地的1.7倍和1.1倍,是藓结皮样地的1.5倍和1.4倍。可见黄绵土上生物结皮覆盖显著降低了草本植物的多样性(P<0.05)。

图3 生物结皮与无结皮样地中草本植物的多样性指数Fig.3 The diversity indices of herbaceous plants from biocrusts and uncrusted soil plots 图中a,b,c表示不同处理之间草本植物的多样性指数差异显著(P<0.05)
2.4 生物结皮对草本植物生物量的影响
两种土壤上的生物结皮均显著降低了草本植物的生物量,其中藓结皮对草本植物生物量的降幅大于蓝藻结皮(图4)。风沙土上,蓝藻结皮和藓结皮样地中的生物量分别为123.2 g/m2和89.7 g/m2,分别比无结皮减少了111.5 g/m2和145.0 g/m2;而黄绵土上,两种生物结皮样地的生物量分别为117.7 g/m2和73.2 g/m2,分别比无结皮减少了127.8 g/m2和172.2 g/m2。方差分析可知,两种土壤上生物结皮样地中草本植物的生物量与无结皮差异显著,即生物结皮显著降低了草本植物的生物量。由图4可知,风沙土上蓝藻结皮和藓结皮样地中草本植物的生物量降幅分别为47.5%和61.8%,而黄绵土上分别为52.1%和70.2%,即黄绵土上生物结皮对草本生物量的降幅更大。
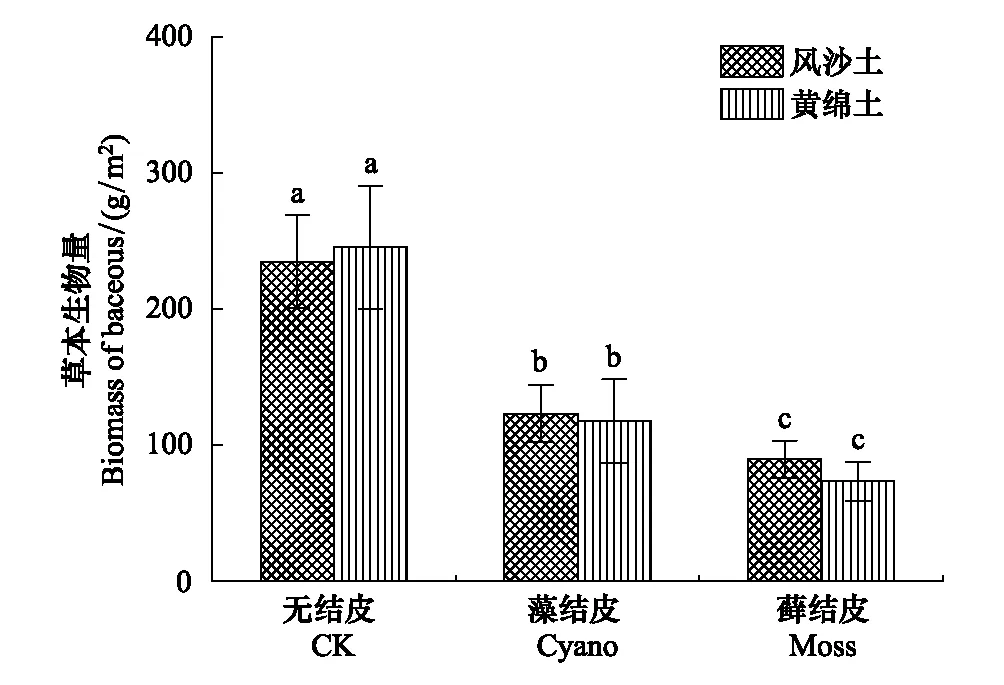
图4 生物结皮与无结皮样地中草本植物的生物量 Fig.4 Biomass of herbaceous plants from biocrusts and uncrusted soil plots
2.5 生物结皮对表层土壤属性的影响


表3 生物结皮与无结皮样地的土壤属性
2.6 生物结皮对草本植物多样性和生物量的影响机制分析

表4 草本植物的多样性和生物量与土壤属性的关联度


图5 生物结皮影响草本植物多样性和生物量的结构方程模型Fig.5 SEM showing the effects of biocrusts on herbaceous plant diversity and biomass SW: 土壤含水量;PR: 土壤穿透阻力;AP: 速效磷含量;ST: 土壤温度;线条粗细程度与路径系数成正比;实线代表正效应;虚线代表负效应
3 讨论
3.1 生物结皮对草本植物多样性的作用机制
本研究表明,与无结皮相比,风沙土上蓝藻结皮和藓结皮样地中草本植物的Patrick和Shannon指数分别降低了1.0、0.2和3.4、0.5;而黄绵土上则分别降低了4.8、0.2和4.2、0.4,即蓝藻结皮和藓结皮明显降低了草本植物的多样性指数,这与庄伟伟等[24]和Corvalán等[12]的研究结果相似。灰色关联度分析表明,土壤穿透阻力、土壤含水量和速效磷含量是影响草本植物多样性的主要因素。这是因为:(1)生物结皮增加了表层土壤的穿透阻力,而土壤穿透阻力的增加限制了植物种子的萌发和幼苗根系的伸展,从而影响了草本植物多样性[6,25];(2)生物结皮增加了土壤水分和养分的表聚现象,减少了深根系草本植物的水分和养分获取,从而影响草本植物多样性[26—27]。庄伟伟等[24]的结果显示,地衣结皮使草本植物的Shannon指数下降了51%,而本研究中,风沙土上蓝藻结皮和藓结皮样地中的Shannon指数分别下降了15%和30%,黄绵土上则分别下降了11%和25%,降幅远小于庄伟伟等的研究。一方面,受气候条件和土壤环境影响,古尔班通古特沙漠草本植物的群落结构可能更不稳定,更容易受生物结皮的影响;另一方面,可能与生物结皮类型和样地选取有关,庄伟伟等的研究主要针对丘间低地的地衣结皮,而本研究主要针对平地上的蓝藻结皮和藓结皮。同时,本研究中,黄绵土上草本植物丰度的降低幅度明显高于风沙土,这是因为,受土壤质地影响,黄绵土的土壤穿透阻力本身就大于风沙土,而生物结皮进一步增加了黄绵土的穿透阻力,使其明显大于风沙土,从而使黄绵土上植物种子的萌发和幼苗根系的伸展更为困难,进而导致草本植物丰度下降[28—29]。需要指出的是,由于生物结皮的复杂性、草本植物类型差异、研究方法差异以及环境条件的不可比性,目前相关研究存在较大争议(表5)。例如,在古尔班通古特沙漠[15]、腾格里沙漠[30]和青藏高原[31]的研究表明,生物结皮有利于提升草本植物的多样性;而在古尔班通古特沙漠南缘[24]和毛乌素沙地[22]的研究结果则显示,生物结皮导致草本植物的多样性降低。因此,仅基于单一区域的研究可能很难全面了解生物结皮对草本植物多样性的影响机制,今后应通过多区域研究来进一步全面揭示生物结皮对草本植物多样性的影响机制。

表5 我国不同区域中生物结皮对草本植物多样性影响的差异
3.2 草本植物群落结构对生物结皮覆盖的响应
本研究发现,生物结皮导致草本植物群落结构发生了显著变化,其中草本植物群落的优势科明显变化,生活型株数占比明显调整,这与多数研究结果相似。例如:在青藏高原,生物结皮使禾本科植物占比上升、莎草科(Cyperaceae)和杂类草占比下降,而豆科占比无显著变化[31];在腾格里沙漠,蓝藻结皮使十字花科(Cruciferae)的比例上升,而藓结皮和地衣结皮增加了牻牛儿苗科(Geraniaceae)的比例[15]。尽管这些研究中的结皮类型、草本植物类型、土壤基质以及气候条件有所差异,但这些研究都一致表明生物结皮改变了草本植物的群落结构,这可能与生物结皮的“筛选”机制和草本植物本身的生物学特性有关[24]。首先,生物结皮在土壤表层形成了致密的毛垫状壳层,致使一些大型种子或表面光滑的种子难以附着并进入土壤,而对一些带“针”或“芒”的种子影响很小,由此改变了土壤种子库结构,进而改变草本植物的群落结构[6,18,22]。其次,生物结皮分泌的多糖物质会附着于种子表面形成包裹层,其对薄皮种子无明显影响,但可能会抑制厚皮种子萌发,从而改变草本植物群落结构[36]。另外,生物结皮导致土壤水分浅层化,不利于深根系草本植物的生长,这可能也是生物结皮影响草本植物群落的重要原因[22]。但也有研究则认为[37],生物结皮对多年生草本植物没有明显影响。这可能与生物结皮盖度有关,生物结皮盖度对草本植物群落的影响存在一定阈值,在达到阈值之前,土壤属性没有发生实质变化,生物结皮的“筛选”作用有限,因此生物结皮不会对草本植物的群落结构产生明显影响。这是这些研究与本研究存在差异的主要原因。
3.3 生物结皮对草本植物生物量的调控机制
4 结论
研究表明,生物结皮覆盖明显改变了黄土高原半干旱气候区的草本植物群落结构,同时显著降低了草本植物的多样性和生物量。研究结果显示,草本植物的多样性和生物量受土壤物理和化学属性两方面的影响。生物结皮对土壤穿透阻力的增加,可能降低了草本植物的种子萌发几率,并限制了幼苗的根系伸展,从而降低了草本植物的多样性;此外,生物结皮通过与草本植物的土壤氮竞争降低了草本植物的生物量。本研究揭示了黄土高原生物结皮对草本植物多样性和生物量的影响程度和作用途径,对于深入了解干旱和半干旱气候区植物的建群机制具有重要意义。
猜你喜欢
新疆大学学报(自然科学版)(中英文)(2022年3期)2022-06-04 13:45:40
草业学报(2021年6期)2021-06-28 11:01:36
读者·校园版(2020年20期)2020-10-13 05:32:58
水土保持通报(2020年6期)2020-03-10 01:46:52
当代水产(2019年9期)2019-10-08 08:02:42
文萃报·周五版(2019年12期)2019-09-10 07:22:44
当代水产(2018年8期)2018-11-02 05:30:42
散文选刊·下半月(2018年2期)2018-02-23 21:11:50
新疆大学学报(自然科学版)(中英文)(2018年4期)2018-02-10 00:09:15
水科学与工程技术(2016年3期)2016-07-10 15:12:44
