
远近端人际压力与FKBP5基因对青少年自伤行为的联合影响:基于发展的视角*
2023-09-08 07:15高叶淼李金文
心理学报 2023年9期
白 荣 高叶淼 李金文 刘 霞
远近端人际压力与基因对青少年自伤行为的联合影响:基于发展的视角*
白 荣 高叶淼 李金文 刘 霞
(北京师范大学发展心理研究院, 北京 100875)
本研究采用为期2年的纵向追踪设计, 在436名青少年中考察远端童年期虐待与近端同伴侵害对青少年自伤的相对作用与影响模式, 以及基因在其中发挥的调节作用。结果发现:(1)远近端人际压力均能显著预测青少年自伤行为, 但二者的相对作用在青春期不同阶段会发生变化; (2)在全样本中, 远近端人际压力均以“压力放大”模式影响自伤行为, 即高童年期虐待放大近期同伴侵害对自伤行为的预测作用; (3)纳入基因后, 在AA纯合子青少年中仍符合“压力放大模型”, 但在AC和CC基因型青少年中, 影响模式则符合“压力敏感模型”, 即经历高童年期虐待的青少年对近期同伴侵害更加敏感, 在低同伴侵害下自伤得分相对较高。研究结果提示, 远近端人际压力对青少年自伤的特定影响模式因其遗传基因而有所差异。
人际压力, 自伤行为,基因, 压力放大模型, 压力敏感模型
1 问题提出
自伤行为是指个体通过切割、抓挠、撞击或灼伤等方式蓄意伤害自己身体组织的行为(Klonsky et al., 2014), 通常始发于青春期早期(Plener et al., 2018), 并在世界范围内的青少年群体中呈现出高发态势。国际流调数据显示, 大约22.9%的青少年在人生中至少自伤过一次, 18.6%的青少年在过去12个月中曾发生过自伤(Gillies et al., 2018), 在我国这一比例则高达26.9% (Liu et al., 2020)。自伤行为不仅会损伤青少年的身体组织, 还会使其面临抑郁、焦虑、人格障碍、药物滥用, 甚至自杀的风险(Nock, 2010), 世界卫生组织已将其列为影响青少年健康的五大威胁因素之一(WHO, 2014)。不过, 尽管青春期是自伤行为的易感期和高发期, 但同时这一阶段青少年神经系统的可塑性较大, 也为干预工作提供了潜在的机会窗口(Nock, 2010)。为此, 在青春期考察自伤行为的风险因素及其作用机制具有重要意义。
发展精神病理学的人际关系理论指出(interpersonal theories of developmental psychopathology) (Rudolph et al., 2016), 人际压力是在特定的人际关系与社会互动中产生的自我内部或自我与环境间失调的结果, 是青少年心理病理问题(如自伤行为)的风险因素, 主要来自父母和同伴这两个特定领域。此外, 人际压力可以进一步分为远端和近端人际压力(田相娟等, 2020)。对于青少年而言, 远端人际压力涉及童年期虐待、长期亲子分离、父母消极教养方式等, 近端人际压力涉及青春期经历的压力性生活事件、同伴侵害等(Davies et al., 2021; Rudolph et al., 2008; Ruttle et al., 2014; 田相娟等, 2020)。一方面, 早期来自父母的虐待这一远端的人际压力, 会引起青少年的消极情绪、较差的认知和行为控制以及社交应对能力, 增加自伤的概率(Nock, 2010)。另一方面, 步入青春期以后, 来自同伴的侵害作为近端的人际压力会影响青少年的心理社会功能, 导致其产生恐惧、抑郁和愤怒等情绪, 进而诱发自伤(Mo et al., 2019)。上述关于远端童年期虐待与近端同伴侵害对青少年自伤行为的预测作用已得到大量研究证实(He & Xiang, 2022; Vergara et al., 2019; Wang & Liu, 2022), 但以往研究多从单一人际压力源出发考察其与青少年自伤行为的关系。随着自主性与独立性的发展, 青少年的关系重心逐渐从家庭内部转移至家庭外部, 同伴的影响日益增强(Laursen & Collins, 2009)。童年期虐待与同伴侵害作为不同的人际压力源, 其性质和强度可能随年龄发生改变。因此在青春期不同阶段, 早期来自父母的虐待与近期来自同伴的侵害对青少年自伤行为的相对作用大小有待探讨。
此外, 早期逆境会改变青少年面临后续人际挑战时的压力反应, 从而导致差异化的发展结果(Rudolph et al., 2016), 远端童年期虐待很可能在近端同伴侵害与青少年自伤行为之间发挥重要的调节作用。目前, 有三种模型为此提供了理论框架。压力放大模型(stress amplification model)认为, 经历高水平童年期逆境的青少年在面临高近期压力时问题行为更多, 而经历低水平童年期逆境的个体, 无论近期压力高低, 其问题行为均相对较少(Rudolph et al., 2016)。压力敏感模型(stress sensitization model)则认为, 童年期逆境增加了个体对近期压力的敏感性, 经历高水平童年期逆境的个体只需要较低的近期压力便能触发问题行为, 因此无论近期压力是高是低, 问题行为都会维持在一个较高的水平; 而经历低水平童年期逆境的个体需要更高的近期压力才会产生问题行为, 也就是说, 随着近期压力的升高, 即便是经历低水平童年期逆境的个体也会出现问题行为(Monroe & Harkness, 2005)。而压力接种模型(stress inoculation model)则指出, 童年期逆境也许为个体提供了发展适应性应对技能的机会, 从而降低个体对后续压力的反应性, 缓冲近期压力对青少年的不良影响, 即经历高水平童年期逆境的个体在面临高近期压力时问题水平反而会降低(Rutter, 2014)。
以往研究在人际关系理论视角下对上述理论模型进行了检验, 但结果存在不一致性。例如, 有研究发现, 经历高童年期逆境的人在承受较大压力时更容易产生抑郁等内化问题, 支持压力放大模型(Morris et al., 2010; Ruttle et al., 2014)。另有研究发现, 童年期逆境增加了个体面临后续低水平压力时的脆弱性, 使个体更容易罹患抑郁症和双向情感障碍, 支持压力敏感模型(Slavich et al., 2011; Weiss et al., 2015)。还有研究则支持压力接种模型(Barzilay et al., 2021; Lyons et al., 2007), 如Barzilay等人(2021)发现经历早期创伤的个体在邻里环境较差时其自杀意念反而更低。总体而言, 现有关于远近端人际压力与青少年发展结果关系的研究主要聚焦于抑郁、自杀等, 考察其对青少年自伤行为作用的研究还相对较少。从人际关系理论视角出发, 结合自伤的四功能理论模型与实证研究结果可以发现, 具有不良人际关系的个体通过自伤来应对人际压力、获得人际利益的动机更加强烈(Nock & Prinstein, 2004; Wadman et al., 2018)。这可能会形成一种恶性循环, 即早期虐待等人际压力导致个体出现自伤行为, 使其由此形成不恰当的应对方式, 从而在青春期面临同伴侵害等近期压力时仍然采用自伤予以应对。例如, Madden等人(2018)发现, 经历高童年期情绪虐待的青少年, 在经历高近期同伴侵害时自伤行为更多, 符合压力放大模型。但也有研究发现了不同结果, 如Hessels等人(2018)在边缘性人格障碍青少年中发现, 经历越多早期逆境的青少年, 在面临较差的近期亲子关系时其自伤行为反而较少, 符合压力接种模型。可见, 人际关系理论模型在解释远近端人际压力对青少年自伤行为的交互作用时存在分歧, 是否存在重要的边界条件以区分不同的压力模型仍有待探讨。
近年来, 来自分子遗传学的研究进展提示, 探究个体基因在远近端人际压力与青少年自伤行为关系中的作用可能为阐明人际关系理论的分歧提供重要契机(Nederhof, 2012; 田相娟等, 2020)。自伤的病因学模型也指出, 环境风险对自伤行为的影响受到个体生物脆弱性因素的调节(Derbidge & Beauchaine, 2014)。下丘脑—垂体—肾上腺(hypothalamic-pituitary-adrenal axis, HPA)轴是机体应对压力的重要调节系统(Normann & Buttenschn, 2020), 与之相关的基因成为研究压力相关精神疾病的重要候选基因。基因位于染色体6p21.31, 编码结合蛋白, 是一种内源性应激神经内分泌系统调节因子, rs3800373是该基因的一个功能性位点, 参与糖皮质激素受体(glucocorticoid receptors, GR)敏感性的调节, 会导致GR信号通路和HPA轴失调(Binder, 2009)。该位点包括AA、AC和CC三种基因型, 其中C等位基因为风险基因型(即AC或CC基因型), 由其引起的水平上升增加了糖皮质激素受体的抵抗性, 进而导致皮质醇对压力和创伤的反应延长, 降低压力应对能力, 增强压力反应性(Touma et al., 2011)。以往研究也证明, 与AA基因型个体相比, C等位基因携带者暴露于单一逆境后, 容易产生抑郁、创伤后应激障碍、认知障碍等精神病理问题(Matosin et al., 2018)。此外, 多重逆境与适应框架(the three hit concept of vulnerability and resilience)基于青少年大脑发育关键阶段的基因环境相互作用提出, 来自遗传基因(hit-1)、早期生活环境(hit-2)与近期生活环境(hit-3)等多个水平的因素共同决定着个体的压力适应行为(Daskalakis et al., 2013)。在经历早期压力后,基因rs3800373位点的C等位基因引发调节稳态的破坏, 会导致参与情绪调节的神经回路发生长期变化, 如杏仁核、眶额皮质等(Matosin et al., 2018), 引发消极的认知与情绪, 增强个体对环境威胁的敏感性(Halldorsdottir et al., 2017; Kwon et al., 2021; White et al., 2012)。即相比于AA纯合子的青少年, 携带C等位基因的青少年由于自身的遗传脆弱性, 在经历早期压力后容易产生神经系统和认知情绪上的脆弱性, 使个体对后续压力的不良影响更加易感。据我们所知, 仅有一项研究考察了远近端人际压力与基因对青少年抑郁的共同影响。研究结果发现, 在含CC纯合子在内的男青少年组中, 母亲消极教养和同伴侵害倾向以压力放大模式影响抑郁, 在含AA纯合子在内的基因组中, 二者对抑郁的影响则符合压力接种模型(田相娟等, 2020), 该结果证实了远近端人际压力对不同基因型青少年的作用模式存在差异。但目前相关的研究仍比较有限, 还需要更多的研究证据予以补充。
回顾自伤领域的研究发现, 仅有少量考察了多巴胺和五羟色胺系统基因与单一人际压力对青少年自伤行为的影响(Kaess et al., 2021), 这直接制约着基于心理生理学或神经调控等实证证据所开发的干预疗法的应用转化。而以往的理论与研究均表明, 无论是环境层面的人际压力, 还是个体层面的压力反应系统, 均与自伤行为存在密切关系。因此本研究推测, 远端童年期虐待、近端同伴侵害与基因rs3800373多态性对青少年自伤行为同样存在交互作用, 在携带C等位基因青少年中, 远近端人际压力的交互模式符合压力放大或压力敏感模型, 在AA纯合子青少年中, 远近端人际压力的交互模式符合压力接种模型。此外, 现有基因环境交互作用的研究多为横断研究, 而基因功能可能在整个青春期阶段发生变化(Zhang et al., 2015), 如的表达会随着年龄的增长而增加, 这可能会加剧不良环境对风险等位基因携带者的影响(Matosin et al., 2018)。因此, 有必要在青春期不同发展阶段考察远端童年期虐待、近端同伴侵害与基因对青少年自伤的动态交互作用。
综上所述, 本研究以童年期虐待和同伴侵害作为远端和近端的人际压力指标, 以基因rs3800373多态性作为遗传基因指标, 采用纵向研究范式, 从青少年初一追踪至初三年级, 分别在青春期早期和中期考察远近端人际压力与基因rs3800373多态性对青少年自伤行为的动态联合作用, 以期为青少年自伤行为的预防与干预提供科学依据。
2 方法
2.1 被试
采用方便抽样的方式选取贵州省4所初中, 在每所中学随机选取4个班级进行被试招募。第一次(T1)数据收集时, 测量青少年的童年期虐待、同伴侵害、自伤行为和家庭基本信息, 2年后进行第二次(T2)数据收集, 测量青少年近期的同伴侵害和自伤行为, 并采集基因数据。T1时共有517名被试, 由于被试转学、请假等原因, T2时有81名被试未能参与施测, 失访率为15.7%, 最终将436名被试(男生49.8%)纳入数据分析, T1时被试的平均年龄为12.84 ± 0.89岁。前人研究表明, 基因环境交互作用效应量通常为0.01~0.02 (Starr et al., 2014), 若要达到80%以上的统计检验力(α = 0.05), 需要395~787名被试(田相娟等, 2020), 本研究样本量经G*power 3.1.9.7软件计算符合统计学要求。
卡方检验结果显示, 失访与未失访被试在性别上没有显著差异(χ2= 0.139,= 1,= 0.709), 独立样本检验结果表明, 失访与未失访被试在年龄(= −0.15,= 512,= 0.885)、父亲受教育程度(= 0.72,= 505,= 0.472)、母亲受教育程度(= −0.12,= 500,= 0.901)和家庭月收入(= −1.39,= 503,= 0.165)等人口学变量上没有显著差异; 在T1童年期虐待(= 0.15,= 515,= 0.880)、T1同伴侵害(= 1.36,= 515,= 0.175)和T1自伤行为(= 1.76,= 515,= 0.079)上均无显著差异, 说明被试不存在结构性流失。
最终436名被试的家庭基本信息如下:在父母受教育程度方面, 65.3%的母亲和39.8%的父亲受教育水平在小学及以下, 28.5%的母亲和51.3%的父亲受教育水平为初中, 4.5%的母亲和6.3%的父亲受教育水平为高中或中专, 1.7%的母亲和2.7%的父亲受教育水平为大学本科或大专以上。此外, 家庭月收入在1000元以下的占37.4%, 在1000~4000元之间的占43.6%, 在4000~8000元之间的占15.8%, 在8000元以上的占3.3%。
2.2 研究工具
2.2.1 童年期虐待
采用Straus等人(1998)编制的亲子冲突解决策略量表(Parent-Child Conflict Tactics Scale, CTSPC)测量青少年初中之前所经历的虐待情况。该量表包括非暴力管教、心理攻击、体罚、身体虐待和严重身体虐待5个分量表。本研究选取心理攻击、体罚、身体虐待和严重身体虐待4个分量表, 在第一次施测时让青少年对其所经历的童年期虐待情况进行评定, 共计18个条目, 例如“父母会冲我大吼大叫” “父母直接用手打我的屁股”等。青少年从0~6进行7点评分, 0~6分别表示“0次” “1次” “2次” “3~5次” “6~10次” “11~20次” “20次以上”。根据量表使用说明, 需对每一条目的得分取中位数进行转化(0 = 0次, 1 = 1次, 2 = 2次, 3 = 4次, 4 = 8次, 5 = 15次, 6 =25次), 得分越高表明青少年经历的童年期虐待越多。该量表已在我国背景下使用, 具有良好的信效度(生笑笑等, 2019)。本研究童年期虐待量表的Cronbach’s α系数为0.84。
2.2.2 同伴侵害
采用Mynard和Joseph (2000)编制, 郭海英等人(2017)修订的多维同伴侵害量表(Multidimensional Peer Victimization Scale, MPVS)测量青少年近一个学期经历的同伴侵害状况。量表包括身体侵害、关系侵害、言语侵害和财务侵害4个维度, 共计18个条目, 例如“在这一学期, 别的同学骂过我” “在这一学期, 别的同学故意在背后说我的坏话、造谣”等。青少年从“没有发生过”到“经常发生”进行4点评分, 得分越高表明青少年受到的同伴侵害越多。本研究T1和T2同伴侵害量表的Cronbach’s α系数分别为0.87和0.92。
2.2.3 自伤行为
采用Gratz (2001)编制Lundh等人(2007)修订的简版自伤行为量表测量青少年的自伤行为。该量表包括9个条目, 例如“故意不让伤口愈合” “故意把自己打出、撞出淤青”等, 由青少年从“从不”到“5次以上”进行4点评分, 得分越高表明青少年的自伤行为越多。简版自伤行为量表已被运用于中国青少年群体, 具有良好的信效度(黄垣成等, 2021)。本研究T1、T2自伤行为量表的Cronbach’s α系数分别为0.90和0.82。
2.2.4 家庭基本信息
采用家庭信息调查表收集青少年性别、年龄、父母受教育程度、家庭月收入等情况。其中, 父母受教育程度分为“小学及以下” “初中” “高中或中专” “大学本科或大专以上”4个等级。家庭月收入为“1000元以下” “1000~4000元” “4000~8000元” “8000元以上”4个等级。把父母受教育程度和家庭月收入分别标准化后取均值, 计算被试的家庭社会经济地位状况。
2.3 基因检测
由经过严格培训的研究生主试按照规范流程采集青少年被试的唾液样本, 并按要求存放至DNA提取试剂盒。利用Assay Designer 3.1软件进行PCR反应引物设计和单碱基延伸引物设计, 使用Agena MassArray系统进行基因分型, 分型结果由MassArray Typer 4.0软件读取, 并检查数据结果的完整性和正确性, 本研究基因分型有效率大于99%。
2.4 施测程序
征得青少年及其家长和学校的知情同意后, 在青少年初一时以班级为单位收集青少年的童年期虐待、同伴侵害、自伤行为以及家庭基本信息, 2年后在青少年初三时再次收集同伴侵害和自伤行为数据, 并进行基因数据的采集。采集基因数据前一天, 由班主任老师通知学生在采样前30分钟内不能进食、饮水、嚼口香糖等, 并确保体温正常。采集过程中, 研究生采集被试的唾液样本并按要求存放, 后交由专业的生物科技公司进行提取、纯化和分型。
2.5 数据处理与分析
首先, 使用SHEsis软件检验基因rs3800373多态性的基因型分布是否符合哈迪温伯格平衡定律。其次, 使用SPSS 26.0进行数据处理与分析。采用皮尔逊积差相关考察各变量之间的相关。然后, 以T1和T2自伤行为作为结果变量, 采用分层回归分析, 在青春期早期和中期, 分别考察青少年经历的童年期虐待、同伴侵害与基因对自伤行为的交互作用, 并在不同基因型青少年分组中检验交互作用符合的理论模型。具体建模方式如下:在青春期早期时, 以T1童年期虐待为远端人际压力, T1同伴侵害为近端人际压力, 将除基因以外的变量标准化, 以T1自伤行为作为因变量进行分层回归, 第一层放入T1童年期虐待、T1同伴侵害和基因rs3800373多态性; 第二层放入T1童年期虐待与T1同伴侵害、T1童年期虐待与基因以及T1同伴侵害与基因三个两项交互; 第三层放入T1童年期虐待、T1同伴侵害、基因的三项交互; 若交互作用显著, 则进行简单斜率分析。最后, 为了进一步检验基因与环境的交互效应随被试年龄变化的结果稳定性, 在青春期中期时, 以T1童年期虐待为远端人际压力, 以T2同伴侵害为近端人际压力, 以T2自伤行为为因变量, 按照T1时的建模思路与方式再次进行数据分析。
3 结果
3.1 FKBP5基因rs3800373多态性基因型分布
基因rs3800373多态性的基因型分布如下:CC = 8.3% (36人), AC = 45.6% (199人), AA = 46.1% (201人)。基因型观测值与期望值吻合良好(χ2= 1.87,= 1,= 0.172), 符合哈迪温伯格平衡定律。次等位基因分布概率(minor allele frequency, MAF)大于5%。
3.2 变量的描述统计与相关分析结果
各变量的平均数、标准差和相关系数见表1。T1童年期虐待、T1和T2同伴侵害与基因rs3800373多态性的相关不显著, 排除了基因—环境相关(rGE)。基因rs3800373多态性与自伤行为的相关不显著。T1童年期虐待、T1和T2同伴侵害与自伤行为均呈显著正相关。T1与T2自伤行为之间显著正相关, 说明青少年自伤行为具有一定的稳定性, 在以T2自伤为因变量的分层回归分析中将T1自伤作为控制变量。
3.3 童年期虐待、同伴侵害与FKBP5基因对自伤行为的交互作用检验
3.3.1 T1童年期虐待、T1同伴侵害与基因对T1自伤行为的交互作用
以T1自伤行为作为结果变量进行分层回归分析, 结果如表2所示。T1童年期虐待、T1同伴侵害对T1自伤行为的主效应显著(= 0.30,= 4.19,< 0.001;= 0.18,= 2.68,= 0.008),基因rs3800373多态性的主效应不具统计学意义(= 0.01,= 0.14,= 0.886)。T1童年期虐待与T1同伴侵害的交互效应显著(= 0.31,= 4.33,< 0.001), T1童年期虐待、T1同伴侵害与基因rs3800373多态性的交互效应不具统计学意义(= −0.03,= −0.40,= 0.687;= −0.01,= −0.02,= 0.987)。T1童年期虐待、T1同伴侵害、基因rs3800373多态性的交互效应显著(= −0.45,= −6.08,< 0.001)。
对T1童年期虐待×T1同伴侵害这一显著的交互项进行简单斜率分析发现, 在全样本青少年中, 当童年期虐待水平较高时, 同伴侵害对青少年自伤行为的作用显著(= 0.22,= 5.62,< 0.001), 即高童年期虐待放大了近期同伴侵害对自伤行为的影响; 当童年期虐待水平较低时, 同伴侵害对青少年自伤行为的作用不具统计学意义(= −0.07,= −1.246,= 0.146); 这一结果符合压力放大模型(见图1a)。
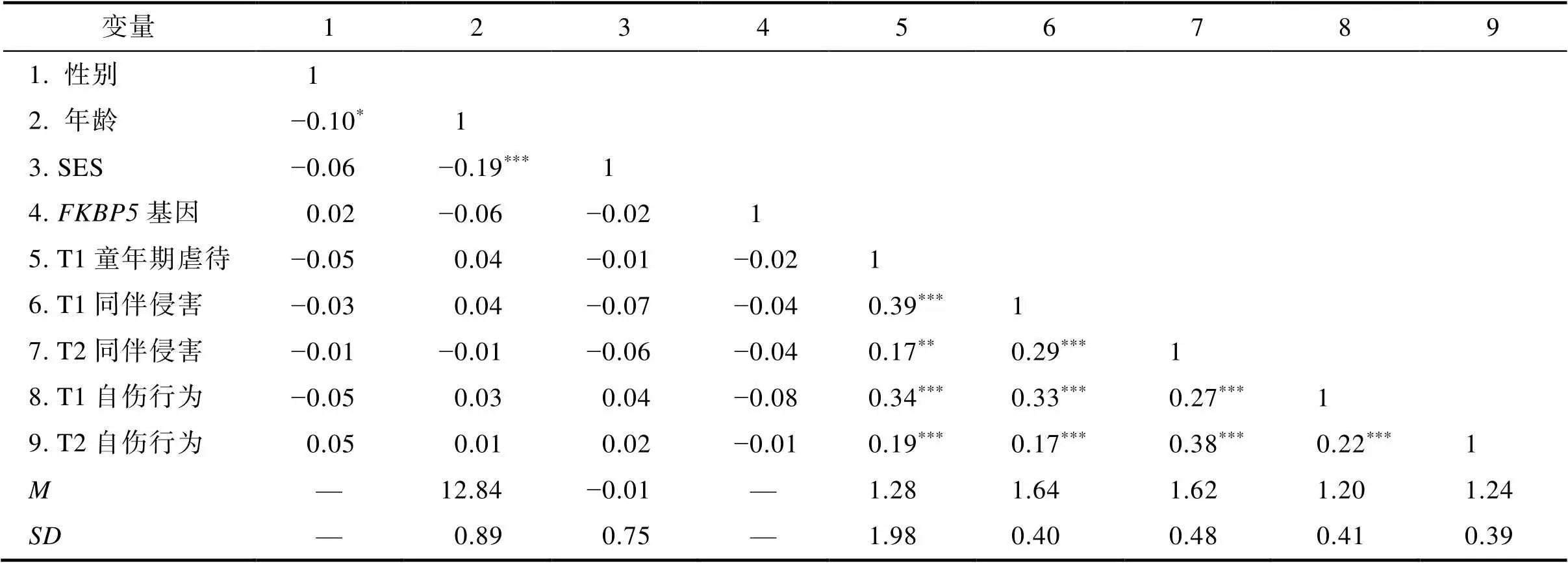
表1 各变量的描述性统计与相关结果
注:SES为家庭社会经济地位的缩写。基于以往研究(Halldorsdottir et al., 2017; Richter et al., 2021), 将AA基因型虚拟编码为0, 将AC/CC基因型虚拟编码为1。*< 0.05;**< 0.01;***< 0.001。
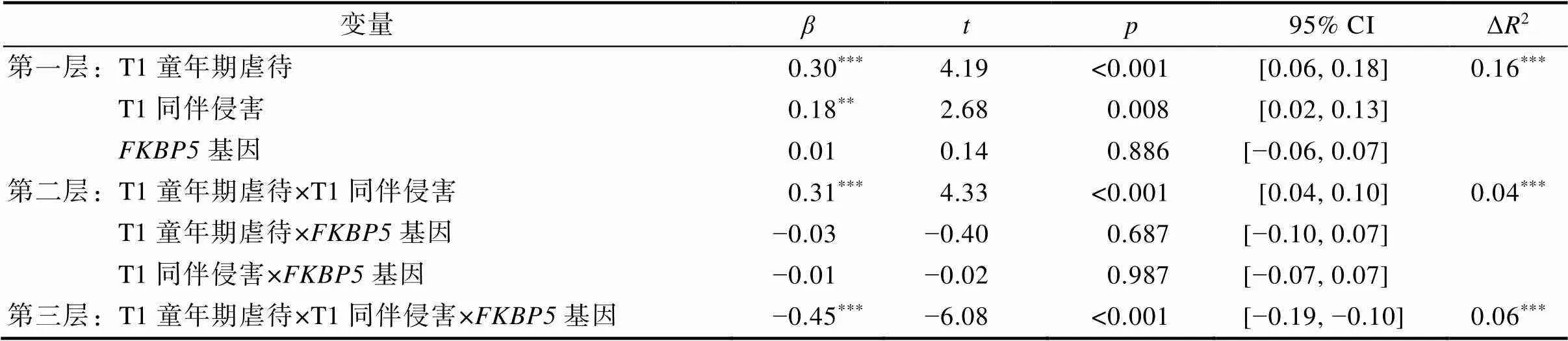
表2 T1童年期虐待、T1同伴侵害与FKBP5基因对T1自伤行为的影响
注:AA基因型 = 0, AC/CC基因型 = 1。*< 0.05;**< 0.01;***< 0.001。
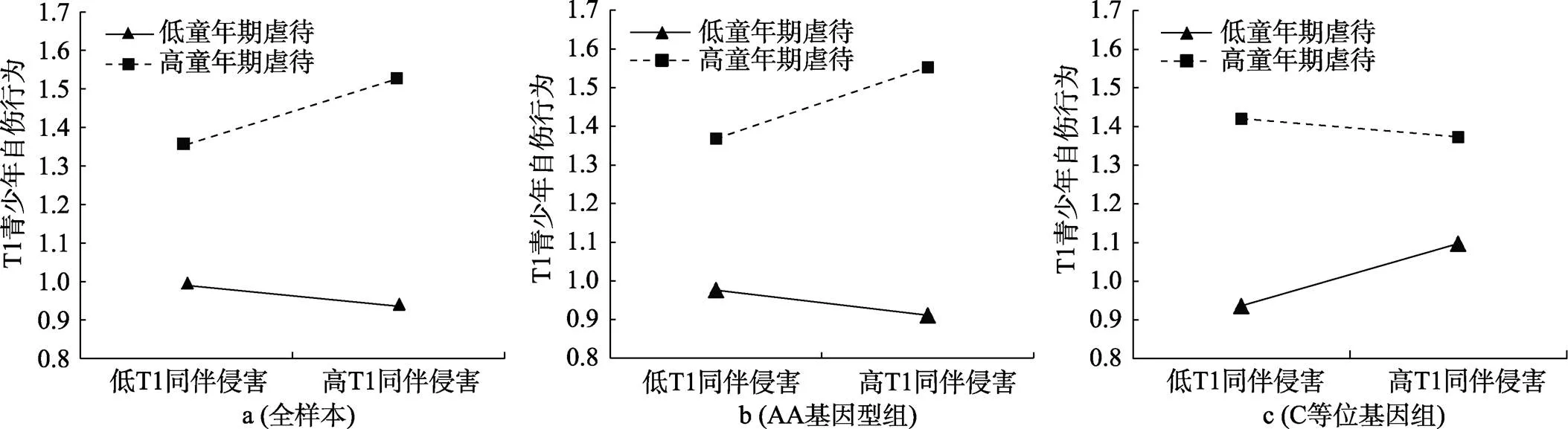
图1 T1童年期虐待与T1同伴侵害对T1青少年自伤行为的交互作用
进一步, 对T1童年期虐待×T1同伴侵害×基因这一显著的三项交互进行分组检验发现, 在携带AA纯合子的青少年中, T1童年期虐待和T1同伴侵害的交互作用显著(= 0.32,= 4.86,< 0.001)。具体来说, 当T1童年期虐待水平较高时, T1同伴侵害对T1自伤行为的作用显著(= 0.23,= 6.33,< 0.001), 即高童年期虐待放大了近期同伴侵害对自伤行为的影响; 当T1童年期虐待水平较低时, T1同伴侵害对T1自伤行为的作用不具统计学意义(= −0.08,= −1.82,= 0.070), 因此符合压力放大模型(见图1b)。在携带C等位基因(即AC/CC基因型)的青少年中, T1童年期虐待和T1同伴侵害的交互作用显著(= −0.31,= −3.93,< 0.001)。当T1童年期虐待水平较高时, 虽然T1同伴侵害对T1自伤行为的效应不具统计学意义(= −0.06,= −1.41,= 0.160), 但此时AC/CC基因型青少年在低近期同伴侵害下自伤行为的得分便整体较高, 即此时青少年对近期人际压力更加敏感, 自伤的阈值更低; 而当T1童年期虐待水平较低时, T1同伴侵害对T1自伤行为作用达到显著水平(= 0.21,= 4.60,< 0.001), 也就是说, 即便童年期虐待水平较低, AC/CC基因型青少年在经历高近期同伴侵害时也容易产生自伤行为, 因此这一结果符合压力敏感模型(见图1c)。综上, 远近端人际压力对不同基因型青少年自伤行为的交互作用模式存在差异, 在AA纯合子青少年中符合压力放大模型, 在C等位基因青少年中符合压力敏感模型。
3.3.2 T1童年期虐待、T2同伴侵害与基因对T2自伤行为的交互作用检验
以T2自伤行为作为结果变量进行分层回归分析, 结果见表3。在控制T1自伤行为后, T1童年期虐待和T2同伴侵害正向预测T2自伤行为(= 0.18,= 2.83,= 0.005;= 0.40,= 6.39,< 0.001),基因rs3800373多态性的主效应不具统计学意义(= 0.04,= 0.94,= 0.347)。T1童年期虐待与T2同伴侵害的交互效应具有统计学意义(= 0.24,= 3.79,< 0.001), T1童年期虐待、T2同伴侵害与基因rs3800373多态性的交互效应不显著= −0.10,= −1.62,= 0.105;= −0.05,= −0.87,= 0.387), T1童年期虐待、T2同伴侵害、基因rs3800373多态性的交互效应达到显著水平(= −0.27,= −4.17,< 0.001)。
对T1童年期虐待×T2同伴侵害这一显著的交互项进行简单斜率分析发现, 在全样本青少年中, 当童年期虐待水平较低时, 同伴侵害对青少年自伤行为的预测作用不显著(= −0.01,= −0.05,= 0.961); 当童年期虐待水平较高时, 同伴侵害显著正向预测青少年自伤行为(= 0.31,= 6.54,< 0.001), 符合压力放大模型(见图2a)。
进一步, 对T1童年期虐待×T2同伴侵害×基因这一显著的三项交互进行分组检验发现, 在携带AA纯合子的青少年中, T1童年期虐待和T2同伴侵害的交互作用显著(= 0.23,= 3.76,< 0.001), 当T1童年期虐待水平较高时, T2同伴侵害显著正向预测T2自伤行为(= 0.32,= 6.25,< 0.001); 当T1童年期虐待水平较低时, T2同伴侵害对T2自伤行为的预测作用不显著(= −0.02,= −0.39,= 0.696), 结果符合压力放大模型(见图2b)。在携带C等位基因青少年中, T1童年期虐待和T2同伴侵害的交互作用显著(= −0.15,= −2.23,= 0.027), 当T1童年期虐待水平较高时, T2同伴侵害对T2自伤行为的预测作用不显著(= 0.04,= 0.81,= 0.420), 但此时同伴侵害较低时自伤行为得分便处于较高水平; 当T1童年期虐待水平较低时, T2同伴侵害显著正向预测T2自伤行为(= 0.22,= 4.12,< 0.001), 说明即使童年期虐待水平较低, 近期同伴侵害的增加也会导致AC/CC基因型青少年产生自伤行为, 结果符合压力敏感模型(见图2c)。上述结果与青春期早期发现的模式一致, 即在不同基因型的青少年中, 远近端人际压力的作用模式符合不同的压力模型。
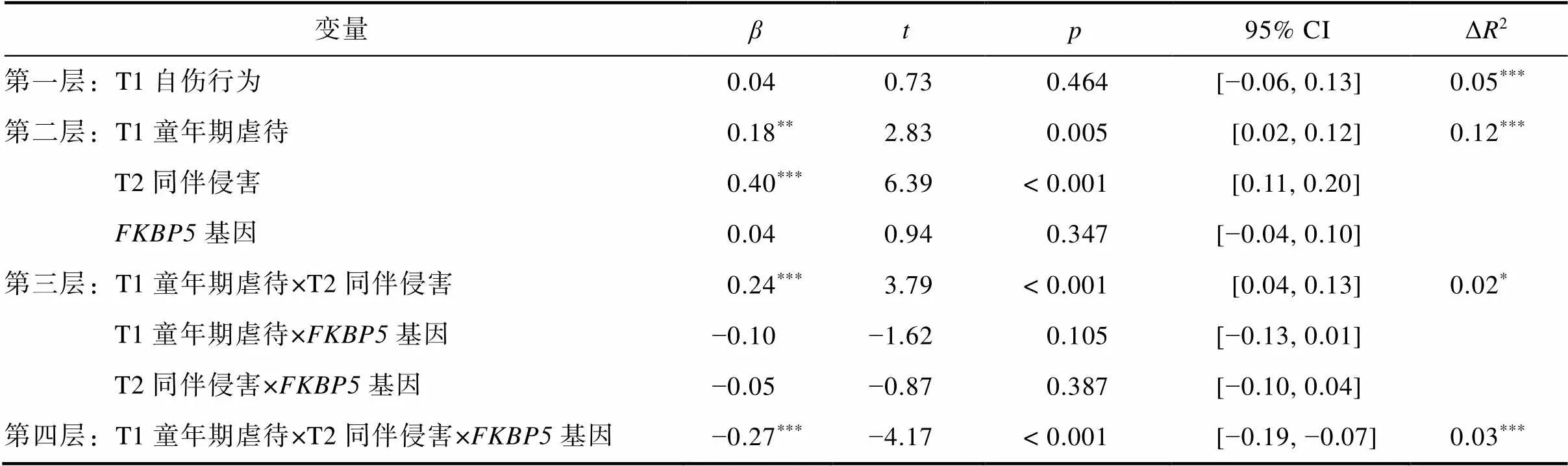
表3 T1童年期虐待、T2同伴侵害和FKBP5基因对T2青少年自伤的影响
注:*< 0.05;**< 0.01;***< 0.001。
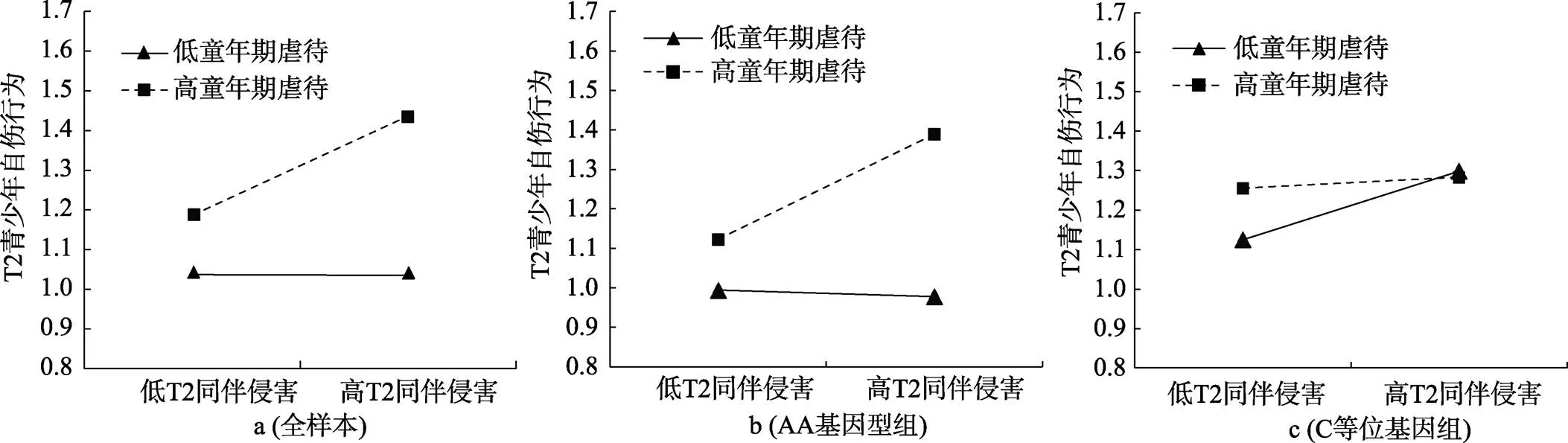
图2 T1童年期虐待与T2同伴侵害对T2青少年自伤的交互作用
3.4 敏感性分析:内部验证
为检验研究结果的可靠性, 本研究进行了内部一致性分析。首先, 本研究将被试随机分为两个子样本, 子样本1 (= 207)和子样本2 (= 229)在性别(= −1.14,= 0.253)、年龄(= 1.49,= 0.138)、基因型(χ2= 0.73,= 0.394)、T1和T2同伴侵害(= −1.59,= 0.113;= 1.32,= 0.188)、T1和T2自伤行为(= −0.89,= 0.372;= −0.52,= 0.603)等均不存在显著差异, 但在T1童年期虐待上差异显著(= −2.03,= 0.043)。在两个子样本中, 以T1和T2自伤为结果变量, 分别进行远近端人际压力与基因的交互作用检验。结果发现, 在子样本1中, T1童年期虐待、T1同伴侵害与基因对T1自伤的交互作用仍然显著(= −0.18,= −2.26,= 0.025), 交互作用方向与总样本保持一致; 但T1童年期虐待、T2同伴侵害与基因对T2自伤的交互作用不再显著(= −0.17,= −1.54,= 0.126)。在子样本2中, T1童年期虐待、T1同伴侵害与基因对T1自伤行为的交互作用(= −0.59,= −5.54,< 0.001), 以及 T1童年期虐待、T2同伴侵害与基因对T2自伤行为的交互作用仍然显著(= −0.24,= −2.61,= 0.010), 交互作用方向均与总样本保持一致。
4 讨论
本研究通过纵向设计在青春期早期和中期, 分别考察了远端童年期虐待、近端同伴侵害与基因对青少年自伤行为的联合作用。研究结果揭示了远近端人际压力对青少年自伤行为的相对作用及交互模式, 并且通过将基因纳入人际关系理论, 进一步揭示了远近端人际压力在不同基因型青少年中的差异化交互模式, 为厘清人际关系理论的适应性及开展更有针对性的干预工作提供了实证支持。
本研究发现, 童年期虐待和同伴侵害均能显著预测青少年早期和中期的自伤行为, 这与以往研究是一致的(He & Xiang, 2022; Vergara et al., 2019; Wang & Liu, 2022)。不同的是, 二者在青春期不同阶段的相对作用发生了变化。具体来说, 在青春期早期, 童年期虐待对青少年自伤行为的预测作用更大, 而到了青春期中期, 同伴侵害对青少年自伤行为的预测作用更大。一方面, 这是因为进入青春期中期, 家庭对青少年的影响逐渐变小。研究发现, 当同时考察童年期虐待与其他风险因素时, 童年期虐待作为远端风险因素对青少年自伤的作用相对减弱(Klonsky & Moyer, 2008)。另一方面, 进入青春期中期后, 同伴的影响逐渐增强。研究数据显示, 在4026名青少年中, 仅有8%受到虐待, 但受到同伴侵害的比例高达30% (Lereya et al., 2015)。除了受影响比例更高之外, 当童年期虐待或父母严厉管教等早期逆境与同伴侵害直接比较时, 同伴侵害对个体自伤行为的不利影响更加严重(Victor et al., 2019)。此外, 与早期的童年期虐待相比, 本研究第二次收集的同伴侵害与自伤行为在发生时间上更加接近, 这可能也是青春期中期的自伤行为更多由同伴侵害而非童年期虐待所解释的原因之一。
根据人际关系理论观点(Rudolph et al., 2016), 远近端人际压力并非“静态孤立”地对青少年自伤行为产生影响, 而是“动态交互”地发挥作用。为此, 本研究通过采用纵向研究范式考察了远端童年期虐待与近端同伴侵害对自伤行为的交互作用。结果显示, 无论在青春期早期还是中期, 远端童年期虐待都会放大近端同伴侵害对自伤行为的预测作用。以往研究发现, 早期逆境使个体暴露在不适应的父母社会化模式与紧张的人际环境中, 削弱了个体发展中的自我意识、情绪调节、应对技能和人际交往能力, 会使青少年沿着越来越不适应的路径前进, 如“滚雪球”般给其带来更严重的人际挑战(Rudolph et al., 2008)。结合本研究发现的第一个结果可以看出, 虽然童年期虐待对青少年自伤行为的直接作用相对减弱, 但却增加了青少年面临后续人际挑战时的脆弱性, 这种脆弱性会放大后期压力的消极影响。研究表明, 经历早期逆境的青少年在近期面临较多负性生活事件时, 其抑郁、焦虑等消极情绪更多(Morris et al., 2010; Ruttle et al., 2014), 容易采用自伤行为来缓解情绪(Nock, 2010)。因此, 当青少年面临近端同伴侵害时, 由于被主流同龄群体边缘化, 应对资源不堪重负, 消极情绪增加(Parker et al., 2006), 此时, 远端童年期虐待便以“压力放大”的模式来增强近端同伴侵害对青少年自伤行为的不良影响。
此外, 远近端人际压力影响青少年自伤行为的交互模式可能因青少年个体遗传差异而有所区别, 本研究根据自伤的病因学模型以及多重逆境与适应框架, 进一步在不同基因型分组的青少年中进行探究。结果发现, 在AA纯合子青少年中, 无论在青春期早期还是中期, 经历高水平童年期虐待的个体在高近期同伴侵害下会产生更多的自伤行为, 这与总样本中的结果一致, 说明远近端人际压力以“压力放大”模式而非“压力接种”模式对AA纯合子青少年的自伤产生影响。这一结果与田相娟等人(2020)的研究结果不一致, 可能是因为本研究关注的远端压力是父母虐待, 这比父母消极教养更加严重。只有早期暴露于具有挑战性的中度压力环境方能促进积极应对, 产生“压力接种”, 而早期暴露于严重的压力则会增加脆弱性, 产生“压力放大” (Daskalakis et al., 2013)。而携带C等位基因青少年的结果则符合压力敏感模型, 这些青少年不仅受到远近端压力的不利影响, 也会受到自身遗传的脆弱性影响, 对人际压力更加敏感。具体来说, 对于经历高水平童年期虐待的C等位基因青少年而言, 轻微的近期同伴侵害便能引发青少年自伤, 这类青少年对压力更加敏感, 产生自伤的阈限降低, 因此无论在低还是高水平近期压力下, 自伤得分都相对较高; 而对于即便只经历了低水平童年期虐待的个体, 他们仍然会对近期压力具有一定的敏感性, 近期压力的升高也会导致其自伤行为的产生。首先, 经历高童年期虐待的C等位基因青少年对近期人际压力更加敏感, 自伤阈限更低、得分更高。这是因为远端的压力体验会影响负责调节情绪、认知及行为功能的大脑回路(Taylor, 2010), 而这些回路是个体后期面临压力时进行行为适应的重要认知和情绪基础(Daskalakis et al., 2013)。一方面, 携带C等位基因的个体在经历儿童期情绪虐待后, 其背侧杏仁核反应性增强, 导致个体面临压力时皮质醇升高和延长, 使其对环境威胁的敏感随之增强(White et al., 2012)。另一方面, 携带C等位基因的个体在高童年期创伤条件下, 左侧眶额皮质和左侧颞中回体积增大, 这与消极的认知(例如, 反刍)和受损的控制能力呈正相关(Kwon et al., 2021), 而这些因素与自伤显著正相关(Hatzopoulos et al., 2022; Zhao et al., 2023)。其次, 即便经历低童年期虐待的C等位基因青少年, 也会随着近端压力的增加而引发自伤。从分子功能角度来看, 随着压力的增多, C等位基因引起了的上调, 降低了GR的可用性, 即减少了GR核易位和对皮质类固醇的敏感性, 从而减少HPA轴的负反馈抑制, 增加了发展精神障碍的风险(Binder, 2009), 如自伤行为。综上, 相比于AA纯合子的青少年, AC/CC基因型的个体在经历高水平远端人际压力后, 轻微的近端人际压力便能导致自伤; 而对于经历低水平远端人际压力的AC/CC基因型个体, 近期人际压力增多也会导致其产生自伤行为。
本研究在一个整合、动态和发展的框架下, 通过同时纳入远端和近端的人际压力以及青少年自身的遗传因素, 在不同发展阶段探索了三者对青少年自伤的联合效应。研究结果揭示了不同时期、不同类型的人际压力源对不同基因型青少年自伤的差异化交互模式, 一方面厘清了人际压力理论模型在解释青少年自伤行为时出现分歧的重要边界条件, 即个体遗传差异, 为压力理论提供了一定的补充, 另一方面为更准确地识别高自伤风险青少年, 制定更具针对性的预防和干预策略提供了实证支持。然而, 本研究仍存在一些局限性以待改进。首先, 本研究样本量经G*power计算虽已符合统计学要求, 但基因环境交互作用研究领域普遍面临效应量小、研究结果不稳健等问题(Border & Keller, 2017)。本研究通过纵向设计, 在青春期早期和中期分别进行了远近端人际压力与基因交互作用的检验, 相当于在发展过程中对基因环境交互作用进行了两次历时性的检验。后续又通过内部一致性检验发现, 除了一个不显著结果外, 其余两个子样本中的结果仍与总样本中保持一致。这些分析为结果的稳定性与可靠性提供了支持, 但鉴于本研究分样本人数较少, 未来研究可以在更大的样本中进行验证。其次, 已有研究发现在人际关系领域, 女生具有更强的人际交往参与和敏感性, 与男生相比可能会表现出更高的压力暴露和反应(Shih et al., 2006)。未来研究可以进一步深入考察远近端人际压力与自伤行为关系的性别差异。最后, 本研究只纳入了基因rs3800373多态性, 可能存在单基因研究中解释力低的问题(Bulik-Sullivan et al., 2015)。但尽管如此, 仍不能小觑重要候选基因与环境对个体发展结果的重要意义(Zannas & Binder, 2014)。未来研究可以通过单倍型分析(Dackis et al., 2012)或者多基因得分(田相娟等, 2020)进一步探讨多基因与多压力环境对青少年自伤行为的联合效应。
5 结论
本研究发现:(1)作为远端和近端的人际压力, 童年期虐待和同伴侵害与青少年自伤行为具有显著关联, 青春期早期时童年期虐待对自伤的相对作用较大, 青春期中期时同伴侵害对自伤的相对作用较大; (2)远近端人际压力对青少年自伤行为存在动态交互作用, 远端童年期虐待放大了近期同伴侵害对自伤行为的不利影响; (3)远近端人际压力对不同基因型的青少年具有差异化影响, AC/CC基因型的青少年经历高童年期虐待后, 对近期同伴侵害更加敏感, 加剧了自伤行为。
Barzilay, R., Moore, T. M., Calkins, M. E., Maliackel, L., Jones, J. D., Boyd, R. C., ... Gur, R. E. (2021). Deconstructing the role of the exposome in youth suicidal ideation: Trauma, neighborhood environment, developmental and gender effects., 100314.
Binder, E. B. (2009). The role of FKBP5, a co-chaperone of the glucocorticoid receptor in the pathogenesis and therapy of affective and anxiety disorders.(1), S186−S195.
Border, R., & Keller, M. C. (2017). Commentary: Fundamental problems with candidate gene-by-environment interaction studies -- reflections on Moore and Thoemmes (2016).(3), 328− 330.
Bulik-Sullivan, B. K., Daly, M. J., Neale, B. M., Loh, P.-R., Finucane, H. K., Ripke, S., Yang, J., Patterson, N., & Price, A. L. (2015). LD Score regression distinguishes confounding from polygenicity in genome-wide association studies.(3), 291−295.
Dackis, M. N., Rogosch, F. A., Oshri, A., & Cicchetti, D. (2012). The role of limbic system irritability in linking history of childhood maltreatment and psychiatric outcomes in low-income, high-risk women: Moderation by FK506 binding protein 5 haplotype.(4), 1237−1252.
Daskalakis, N. P., Bagot, R. C., Parker, K. J., Vinkers, C. H., & de Kloet, E. R. (2013). The three-hit concept of vulnerability and resilience: Toward understanding adaptation to early- life adversity outcome.(9), 1858−1873.
Davies, P. T., Thompson, M. J., Martin, M. J., & Cummings, E.M. (2021). The vestiges of childhood interparental conflict: Adolescent sensitization to recent interparental conflict.(3), 1154−1172.
Derbidge, C., & Beauchaine, T. (2014). A developmental model of self-inflicted injury, borderline personality, and suicide risk. In M. Lewis , & K. Rudolph (Eds). Springer Boston, MA.
Gillies, D., Christou, M. A., Dixon, A. C., Featherston, O. J., Rapti, I., Garcia-Anguita, A., … Christou, P. A. (2018). Prevalence and characteristics of self-harm in adolescents: Meta-analyses of community-based studies 1990-2015.(10), 733−741.
Gratz, K. L. (2001). Measurement of deliberate self-harm: Preliminary data on the deliberate self-harm inventory.(4), 253−263.
Guo, H. Y., Chen, L. H., Ye, Z., Pan, J., & Lin, D. H. (2017). Characteristics of peer victimization and the bidirectional relationship between peer victimization and internalizing problems among rural-to-urban migrant children in China: A longitudinal study.(3), 336−348.
[郭海英, 陈丽华, 叶枝, 潘瑾, 林丹华. (2017). 流动儿童同伴侵害的特点及与内化问题的循环作用关系: 一项追踪研究.(3), 336−348.]
Halldorsdottir, T., de Matos, A. P. S., Awaloff, Y., Arnarson, E. Ö., Craighead, W. E., & Binder, E. B. (2017). FKBP5 moderation of the relationship between childhood trauma and maladaptive emotion regulation strategies in adolescents., 61−65.
Hatzopoulos, K., Boyes, M., & Hasking, P. (2022). Relationships between dimensions of emotional experience, rumination, and nonsuicidal self-injury: An application of the emotional cascade model.(4), 692−709.
He, N., & Xiang, Y. (2022). Child maltreatment and nonsuicidal self-injury among Chinese adolescents: The mediating effect of psychological resilience and loneliness., 106335.
Hessels, C. J., Laceulle, O. M., van Aken, M. A. G., Resch, F., & Kaess, M. (2018). Differentiating BPD in adolescents with NSSI disorder: The role of adverse childhood experiences and current social relationships., 20.
Huang, Y. C., Zhao, Q. L., & Li, C. N. (2021). How interpersonal factors impact the co-development of depression and non-suicidal self-injury in Chinese early adolescents.(5), 515−526.
[黄垣成, 赵清玲, 李彩娜. (2021). 青少年早期抑郁和自伤的联合发展轨迹:人际因素的作用.(5), 515−526.]
Kaess, M., Hooley, J. M., Klimes-Dougan, B., Koenig, J., Plener, P. L., Reichl, C., ... Cullen, K. R. (2021). Advancing a temporal framework for understanding the biology of nonsuicidal self- injury: An expert review., 228−239.
Klonsky, E. D., & Moyer, A. (2008). Childhood sexual abuse and non-suicidal self-injury: Meta-analysis.(3), 166−170.
Klonsky, E. D., Victor, S. E., & Saffer, B. Y. (2014). Nonsuicidal self-injury: What we know, and what we need to know.(11), 565−568.
Kwon, A., Kim, S., Jeon, H., Lee, H. S., & Lee, S.-H. (2021). Influence of FKBP5 variants and childhood trauma on brain volume in non-clinical individuals., 663052.
Laursen, B., & Collins, W. A. (2009). Parent-child relationships during adolescence. In R. M. Lerner & L. Steinberg (Eds.),(pp. 1−42). Wiley.
Lereya, S. T., Copeland, W. E., Costello, E. J., & Wolke, D. (2015). Adult mental health consequences of peer bullying and maltreatment in childhood: Two cohorts in two countries.(6), 524−531.
Liu, S., You, J., Ying, J., Li, X., & Shi, Q. (2020). Emotion reactivity, nonsuicidal self-injury, and regulatory emotional self-efficacy: A moderated mediation model of suicide ideation., 82−89.
Lundh, L., Karim, J., & Quilisch, E. (2007). Deliberate self-harm in 15-year-old adolescents: A pilot study with a modified version of the Deliberate Self-Harm Inventory.(1), 33−41.
Lyons, D. M., & Parker, K. J. (2007). Stress inoculation- induced indications of resilience in monkeys.(4), 423−433.
Madden, A. R., Are, F., Rea, H., Thomassin, K., & Shaffer, A. (2018). Perceived negative peer relationships moderate the association between childhood emotional abuse and nonsuicidal self-injury.(12), 3994−4000.
Matosin, N., Halldorsdottir, T., & Binder, E. B. (2018). Understanding the molecular mechanisms underpinning gene by environment interactions in psychiatric disorders: The FKBP5 model.(10), 821−830.
Mo, J., Wang, C., Niu, X., Jia, X., Liu, T., & Lin, L. (2019). The relationship between impulsivity and self-injury in Chinese undergraduates: The chain mediating role of stressful life events and negative affect., 259−266.
Monroe, S. M., & Harkness, K. L. (2005). Life Stress, the “Kindling” hypothesis, and the recurrence of depression: Considerations from a life stress perspective.(2), 417−445.
Morris, M. C., Ciesla, J. A., & Garber, J. (2010). A prospective study of stress autonomy versus stress sensitization in adolescents at varied risk for depression.(2), 341−354.
Mynard, H., & Joseph, S. (2000). Development of the multidimensional peer-victimization scale.(2), 169−178.
Nederhof, E. (2012). The mismatch hypothesis of psychiatric disease.(5), 689−690.
Nock, M. K. (2010). Self-injury., 339−363.
Nock, M. K., & Prinstein, M. J. (2004). A functional approach to the assessment of self-mutilative behavior.(5), 885−890.
Normann, C., & Buttenschøn, H. N. (2020). Gene-environment interactions between HPA-axis genes and childhood maltreatment in depression: A systematic review.(3), 1−30.
Parker, J. G., Rubin, K. H., Erath, S. A., Wojslawowicz, J. C., & Buskirk, A. A. (2006). Peer relationships, child development, and adjustment: A developmental psychopathology perspective. In D. Cicchetti & D. J. Cohen. (Eds.),(Vol. 1, 2nd ed., pp. 419−493). Hoboken, NJ: John Wiley
Plener, P. L., Kaess, K., Schmahl, C., Pollak, S., Fegert, J. M., Brown, R. C. (2018). Nonsuicidal self-injury in adolescents.(3), 23−30.
Richter, A., Al-Bayati, M., Paraskevopoulou, F., Krämer, B., Pruessner, J. C., Binder, E. B., & Gruber, O. (2021). Interaction of FKBP5 variant rs3800373 and city living alters the neural stress response in the anterior cingulate cortex.(4), 421−429.
Rudolph, K. D., Flynn, M., & Abaied, J. L. (2008). A developmental perspective on interpersonal theories of youth depression. In J. R. Z. Abela & B. L. Hankin (Eds.),(pp. 79−102). New York, NY: Guilford.
Rudolph, K. D., Lansford, J. E., & Rodkin, P. C. (2016). Interpersonal Theories of Developmental Psychopathology. In D. Cicchetti (Eds.),(pp. 1−69). John Wiley & Sons, Inc.
Rutter, M. (2014). Nature-nurture integration. In M. Lewis & K. D. Rudolph (Eds.),(3rd ed., pp. 45−65). New York, NY: Plenum.
Ruttle, P. L., Armstrong, J. M., Klein, M. H., & Essex, M. J. (2014). Adolescent internalizing symptoms and negative life events: The sensitizing effects of earlier life stress and cortisol.(4 Pt 2), 1411−1422.
Sheng, X. X., Li, J. Y., Niu, H., & Wang, M. F. (2019). Parental harsh discipline and adolescents’ externalizing problem behaviors: The moderating role of puberty timing.,(6), 1199− 1204+1209.
[生笑笑, 李静雅, 牛骅, 王美芳. (2019). 父母严厉管教与青少年外化问题行为的关系:青春发育时相与性别的调节作用.(6), 1199−1204+1209.]
Shih, J. H., Eberhart, N. K., Hammen, C. L., & Brennan, P. A. (2006). Differential exposure and reactivity to interpersonal stress predict sex differences in adolescent depression.(1), 103−115.
Slavich, G. M., Monroe, S. M., & Gotlib, I. H. (2011). Early parental loss and depression history: Associations with recent life stress in major depressive disorder.(9), 1146−1152.
Starr, L. R., Hammen, C., Conway, C. C., Raposa, E., & Brennan, P. A. (2014). Sensitizing effect of early adversity on depressive reactions to later proximal stress: Moderation by polymorphisms in serotonin transporter and corticotropin releasing hormone receptor genes in a 20-year longitudinal study.(4pt2), 1241− 1254.
Straus, M. A., Hamby, S. L., Finkelhor, D., Moore, D. W., & Runyan, D. (1998). Identification of child maltreatment with the parent-child conflict tactics scales: Development and psychometric data for a national sample of American parents.(4), 249−270.
Taylor, S. E. (2010). Mechanisms linking early life stress to adult health outcomes.(19), 8507−8512.
Tian, X. J., Cao, Y. M., & Zhang, W. X. (2020). The influence of maternal negative parenting, peer victimization and FKBP5 gene on adolescent depressive symptoms.(12), 1407−1420.
[田相娟, 曹衍淼, 张文新. (2020). 母亲消极教养、同伴侵害与FKBP5基因对青少年抑郁的影响.(12), 1407−1420.]
Touma, C., Gassen, N. C., Herrmann, L., Cheung-Flynn, J., Büll, D. R., Ionescu, I. A., ... Rein, T. (2011). FK506 binding protein 5 shapes stress responsiveness: Modulation of neuroendocrine reactivity and coping behavior.(10), 928−936.
Vergara, G. A., Stewart, J. G., Cosby, E. A., Lincoln, S. H., & Auerbach, R. P. (2019). Non-suicidal self-injury and suicide in depressed adolescents: Impact of peer victimization and bullying., 744−749.
Victor, S. E., Hipwell, A. E., Stepp, S. D., & Scott, L. N. (2019). Parent and peer relationships as longitudinal predictors of adolescent non-suicidal self-injury onset.(1), 1−13.
Wadman, R., Vostanis, P., Sayal, K., Majumder, P., Harroe, C., Clarke, D., Armstrong, M., & Townsend, E. (2018). An interpretative phenomenological analysis of young people’s self-harm in the context of interpersonal stressors and supports: Parents, peers, and clinical services., 120−128.
Wang, Q., & Liu, X. (2022). Child abuse and non-suicidal self-injury among Chinese migrant adolescents: The moderating roles of beliefs about adversity and family socioeconomic status.(3−4), 3165−3190.
Weiss, R. B., Stange, J. P., Boland, E. M., Black, S. K., LaBelle, D. R., Abramson, L. Y., & Alloy, L. B. (2015). Kindling of life stress in bipolar disorder: Comparison of sensitization and autonomy models.(1), 4−16.
White, M. G., Bogdan, R., Fisher, P. M., Muñoz, K. E., Williamson, D. E., & Hariri, A. R. (2012). FKBP5 and emotional neglect interact to predict individual differences in amygdala reactivity.(7), 869−878.
World Health Organization. (2014).. Retrieved Aug 17, 2022, from https:// www.who.int/publications/i/item/WHO-FWC-MCA-14.05
Zannas, A. S., & Binder, E. B. (2014). Gene-environment interactions at the FKBP5 locus: Sensitive periods, mechanisms and pleiotropism.(1), 25−37.
Zhang, W., Cao, Y., Wang, M., Ji, L., Chen, L., & Deater- Deckard, K. (2015). The dopamine D2 receptor polymorphism (DRD2 TaqIA) interacts with maternal parenting in predicting early adolescent depressive symptoms: Evidence of differential susceptibility and age differences.(7), 1428− 1440.
Zhao, L., Zhou, D., Ma, L., Hu, J., Chen, R., He, X., … Kuang, L. (2023). Changes in emotion-related EEG components and brain lateralization response to negative emotions in adolescents with nonsuicidal self-injury: An ERP study.(8), 114324.
Combined effects of distal and proximal interpersonal stress andgene on adolescent self-injury behavior: The developmental perspective
BAI Rong, GAO Yemiao, LI Jinwen, LIU Xia
(Institute of Developmental Psychology, Beijing Normal University, Beijing 100875, China)
Self-injury usually emerges in early adolescence and has a high incidence among adolescents worldwide. Self-injury not only damages body tissue but is also associated with depression, anxiety, personality disorders, substance abuse, and a higher-than-average risk of suicide. Given the high incidence of self-injury and the severity of its consequences, it is important to explore its predictors and specific mechanisms. Interpersonal theories of developmental psychopathology maintain that interpersonal stress is a critical risk factor for adolescent self-injury behavior. However, the ways the source and duration of exposure to that stress affect adolescent self-injury behavior are unclear. Adolescents also differ in their sensitivity to interpersonal stress. Stress-related genetic factors may play an important moderating role. The current study selected child abuse and recent peer victimization as distal and proximal interpersonal stress, respectively, andgene rs3800373 polymorphism as the genetic factor. The purpose of this study was to build upon the results of previous studies by exploring the relative and interactive effects of distal and proximal interpersonal stress on adolescent self-injury behavior.
The participants were 436 adolescents (12.84 ± 0.89 years, 49.8% males) recruited from four junior high schools in Guizhou Province. All were tracked from grade 7 to grade 9. At Time 1, adolescents reported child abuse via the Parent-Child Conflict Tactics Scale, reported peer victimization via the Multidimensional Peer Victimization Scale, and reported self-injury behavior via the Short Version of Self-Injury Behavior Scale. At Time 2, adolescents reported peer victimization and self-injury behavior, and saliva samples were collected. Genotyping with respect to thegene was performed with Agena MassArray software, and the corresponding typing results were analyzed using MassARRAY Typer software version 4.0.
Results showed that both distal and proximal interpersonal stress significantly predicted adolescent self-injury behavior, but the relative effect sizes differed in early and middle adolescence. Across the entire sample, distal and proximal interpersonal stress had an interactive effect on adolescent self-injury, and the interaction pattern was consistent with the stress amplification model. However, when thegene was considered, the interaction pattern was found to differ between adolescents in different genotype groups. Specifically, compared with AA homozygous adolescents who experienced less childhood abuse, those who experienced more child abuse were easily to be impacted by recent peer victimization and engage in NSSI. This was consistent with the stress amplification model. However, in adolescents with the AC/CC genotype who experienced more childhood abuse, mild recent peer victimization triggered adolescent self-injury. These participants showed lower self-injury thresholds and higher scores for self-injury than those who experienced less childhood abuse, which was consistent with a stress sensitization model. These relationships were stable in both early and middle adolescence.
These findings showed different patterns of interaction between interpersonal and intrapersonal factors on self-injury behavior in adolescents of different genotypes. Using an integrative, dynamic, and developmental framework, this study provides important insights into the relevant interpersonal theories. It is also valuable for the accurate identification of adolescents at high risk of self-injury and for both prevention and intervention.
interpersonal stress, self-injury behavior,gene, stress amplification model, stress sensitization model
2022-09-23
* 国家自然科学基金项目(31900772)。
刘霞, E-mail: liuxia@bnu.edu.cn
B844; B845
猜你喜欢
大学(2021年2期)2021-06-11
意林·全彩Color(2019年6期)2019-07-24
新闻传播(2018年12期)2018-09-19
青少年科技博览(中学版)(2017年10期)2018-01-19
公民与法治(2016年12期)2016-05-17
剑南文学(2015年2期)2015-02-28
现代检验医学杂志(2015年6期)2015-02-06
实验动物与比较医学(2014年5期)2014-02-28
当代修辞学(2013年3期)2013-01-23
中国糖料(2013年1期)2013-01-22
