
入侵植物石茅种子萌发及幼苗生长对干旱胁迫的响应特征
2023-09-06 10:01:16范志伟黄乔乔
草业科学 2023年8期
王 亚,刘 延,范志伟,黄乔乔
(农业农村部热带作物有害生物综合治理重点实验室 / 中国热带农业科学院环境与植物保护研究所, 海南 海口 571101)
石茅(Sorghum halepense)原产于地中海地区,是一种适应性强、竞争能力强、繁殖快、危害大、难以防除的禾本科高粱属多年生杂草[1],是农业上最危险的十大恶性杂草之一[2],也是我国禁止输入的有害生物[3]。目前在我国的海南、广东、香港、台湾、江苏、吉林等地均有分布。
外来植物入侵往往对当地生态系统、生物多样性、经济和环境造成巨大的负面影响[4]。在入侵过程中,入侵植物常会遭遇生物和非生物胁迫。干旱是入侵植物经常会遭遇的胁迫环境之一[5]。随着全球气温升高,降水格局发生变化,干旱等极端气候事件频繁发生[6-7],水分变化对植物的生长及分布影响愈发突出。入侵植物对环境具有较强的适应能力,可通过形态特征及生理适应性等的变化来适应不同的生境条件[8]。已有研究表明,外来植物种子的生长和萌发及其幼苗的形态特征及生理变化过程会受到干旱胁迫的影响[9-10]。如PEG 溶液渗透势小于-0.5 MPa 时,入侵杂草Caucalis platycarpos发芽率严重减少[9];入侵植物节节麦(Aegilops tauschi)幼苗在干旱胁迫后,其叶生物量比、比叶面积均下降,根生物量比及根冠比增加[11];三裂叶蟛蜞菊(Wedelia trilobata)的光合作用对干旱胁迫较敏感[12]。
石茅对不同生境具有很强适应性,不仅能在农田、耕地、荒地、公路边生长,在旱地、水田也能正常生长、抽穗、成熟。海南夏季高温及干湿季交替(5 月-10 月为雨季,降水量占全年的70%~90%)可使土壤表层时常处于比较干旱的状态,干旱在一年内常有发生[13]。以往针对石茅的研究主要集中于对其形态学特征、生物学特性[14]、化感作用[15]及细胞学指标[16]等方面。虽然对石茅种子萌发特征的影响已经有研究报道[17],但其种子萌发及幼苗生长对水分条件的生态适应性尚未涉及。因此,本研究以海南省恶性入侵杂草石茅为研究对象,采用培养皿滤纸法和盆栽控水法分别模拟不同程度的土壤干旱情况,以期观察和评价不同PEG-6000 质量浓度下石茅种子的萌发能力及出苗情况;研究不同土壤水分条件下其幼苗生长、生物量分配及生理指标变化特征。这有助于明确石茅对干旱的适应性及其相关机制,对预测其野外分布范围及扩散能力具有重要意义,也为采取有效防治措施提供理论依据。
1 材料与方法
1.1 材料
试验所用石茅种子于2021 年7 月在海南省儋州市宝岛新村(19°30′22.7″ N,109°23′23.0″ E)采集,分别从50 株石茅植株上采集种子,并将所有的种子混合,带回实验室自然风干后保存于4 ℃种子低温低湿储藏柜中备用。用万分之一天平测得其千粒重为2.890 g。试验在中国热带农业科学院儋州院区的实验室及温室内进行。
1.2 方法
萌发试验:采用培养皿滤纸法,于2021 年8 月20 日开始。首先用研磨法去除石茅种子的颖片,然后挑选出种皮完整、大小均一的颖果,并将所有的颖果用2%次氯酸钠消毒10 min 后用去离子水反复清洗干净备用。用聚乙二醇(PEG-6000)溶液模拟干旱胁迫,设7 个不同浓度:0 (去离子水)、2.5%、5.0%、7.5%、10%、12.5%、15%,并根据王慧慧等[18]将2.5%~5.0%作为轻度干旱处理,7.5%~10%作为中度干旱处理,12.5%~15%作为重度干旱处理。将消毒后的种子分别播种于放有双层滤纸、直径为9 cm的培养皿中。每个处理5 个重复,每个重复20 粒种子。萌发试验在温度为25 ℃、光照强度为12 000 lx的人工气候培养箱中进行,周期为光照和黑暗培养各12 h。试验开始后,每天观察并记录种子萌发情况,以胚根长度达到种子长度的1/2 作为种子萌发的标准,连续5 d 没有种子萌发视为萌发结束。采用质量平衡法[19],向培养皿中添加PEG-6000 溶液和去离子水,每2 d 换一次滤纸以减少水势变动。萌发试验持续20 d。萌发试验结束后从各重复中随机挑选5 粒正常发芽的种子,用游标卡尺(精度为0.02 mm)测量其胚芽和胚根的长度,并计算种子的发芽率、发芽指数、活力指数及胚根/胚芽。
式中:Gt为t日发芽数,Dt为相应的天数,S为平均初生胚根长[20]。
盆栽试验:试验开始于2021 年9 月15 日,首先将石茅种子播入穴盘育苗。30 d 后选取大小一致的苗移入装有3.5 kg 土壤(田间土壤 : 沙土 = 1 : 1)的花盆中。该土壤pH 为6.25,全氮、有机质、碱解氮、速效钾及有效磷的含量分别为0.61 g·kg-1、5.15 g·kg-1、44.12 mg·kg-1、39.21 mg·kg-1和11.16 mg·kg-1,田间持水量为24.78%。试验设置4 个不同的水分处理,其土壤含水量分别为田间持水量(WHC)的100%、80%、60%、40%,将80% WHC 作为轻度干旱胁迫,60%作为中度干旱胁迫,40%作为重度干旱处理,每个处理设置5 个重复。
试验过程中采用称重法控制土壤含水量,采用千分之一的电子天平每天17:30 至18:30 定时称重补水,使土壤含水量分别维持在24.78%、19.82%、14.87%和9.91%。花盆之间保持20 cm 距离,每周随机变换花盆位置一次。2 个月后,测量植物的株高,统计植物叶片数量,用SPAD-502Plus 叶绿素含量测定仪测量SPDA 值,YMJ-B 活体叶面积仪测量植物茎最上方完全展开叶片的面积。收获后,分别将植物的地上部和地下部洗干净放入纸袋中置于105 ℃烘箱中杀青,再于70 ℃烘干72 h,最后用万分之一天平称量各部分干重。并计算比叶面积、根冠比及总生物量。
1.3 数据分析
采用SPSS 22.0 对数据进行单因素方差分析(One-way ANOVA)。其中种子萌发的所有数据及相对叶绿数含量数据进行了对数转化以满足方差齐性,并采用Tukey 检验法进行各处理间的差异显著性分析。采用Origin 9.1 绘图。图中给出的数据为平均值 ± 标准差。
2 结果与分析
2.1 种子萌发特征
不同PEG 浓度处理下,石茅种子之间的发芽率有差异。石茅种子的发芽率随PEG浓度的增加先升高后降低(表1),2.5% PEG 浓度胁迫下石茅种子发芽率最高,达到73%;15% PEG 浓度条件下种子发芽率最低,为4%。在0、2.5%、5% PEG这3 个浓度条件下石茅种子发芽率无显著差异(P> 0.05)。与0 PEG 处理相比,7.5%、10%、12.5%、15% PEG 浓度条件下石茅种子的发芽率分别显著降低了69.23%、76.92%、87.69%、93.85% (P< 0.05)。
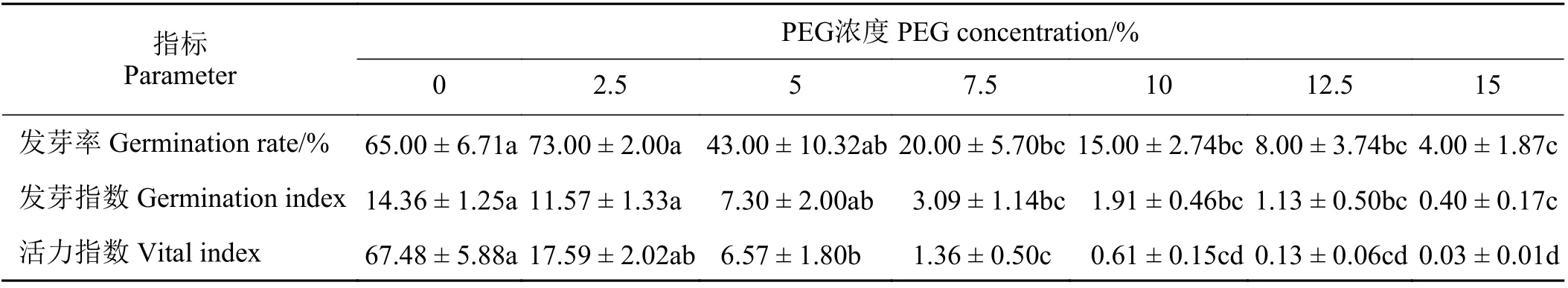
表1 不同PEG 浓度处理对石茅种子发芽率、发芽指数及活力指数的影响Table 1 Effects of drought stress on the germination rate, germination index, and vigor index of Sorghum halepense
不同PEG 浓度处理下,石茅种子的发芽指数及活力指数有差异。石茅种子发芽指数随PEG 浓度的增加而降低(表1)。在0、2.5%、5% PEG 这3 个浓度条件下石茅种子发芽指数无显著差异(P> 0.05)。与0 PEG 处理相比,7.5%、10%、12.5%、15% PEG 浓度条件下石茅种子的发芽指数分别显著降低了78.48%、86.68%、92.15%、97.19% (P< 0.05)。石茅种子活力指数也随PEG 浓度的增加而降低(表1)。与0 PEG 处理相比,5%、7.5%、10%、12.5%、15% PEG浓度条件下石茅种子的发芽指数分别显著降低了90.27%、97.99%、99.09%、99.81%、99.95% 。
不同PEG 浓度处理下,石茅种子的胚根、胚芽长度及胚根/胚芽有明显差异。随着PEG胁迫浓度的增加,石茅种子胚根、胚芽的长度均呈现缩短趋势(图1A)。不同PEG 浓度胁迫条件下石茅胚根长明显低于0 PEG 处理,2.5% PEG浓度条件下其胚根长已降低了67.66%,干旱胁迫明显抑制了石茅的胚根生长;而其胚芽长度在2.5%和5% PEG 浓度条件下与0 PEG 处理无显著差异(P> 0.05),与0 PEG 处理相比,7.5%、10%、12.5%和15% PEG浓度条件下胚芽长度分别降低了76.92%、88.89%、96.91%、98.23%(P< 0.05)。石茅的胚根/胚芽随PEG 胁迫浓度的增加先降低再升高(图1B),在5% PEG 浓度条件下其胚根/胚芽最低(0.79),而15% PEG 浓度时最高(2.37)。2.5%、5%、7.5% PEG 浓度条件下胚根/胚芽显著低于0 PEG 处 理(P< 0.05),分 别 降 低 了53.67%、60.71%,53.91%。10%、12.5%和15%浓度条件下其胚根/胚芽与0 PEG 处理无显著差异。
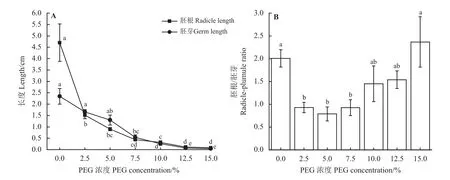
图1 不同PEG 浓度胁迫下石茅种子的胚根、胚芽长度及胚根/胚芽Figure 1 Radicle and germ length and the radicle/germ length ratio of S.halepenseseeds under different polyethylene glycol (PEG) treatments
2.2 幼苗生长特征
随着水分含量的降低石茅幼苗株高、叶片数、比叶面积及相对叶绿素含量呈现下降趋势(表2)。不同水分条件下石茅的株高、叶片数、相对叶绿素含量差异明显,但比叶面积无显著差异(P>0.05)。与100% WHC 相比,60%和40% WHC条件下石茅幼苗的株高分别显著降低了20.02%和39.53% (P<0.05),但在100% WHC 和80% WHC 条件之间差异不显著(P> 0.05)。在100% WHC 条件下石茅幼苗平均叶片数为10 个,60%和40% WHC 条件下幼苗叶片数分别显著降低49.02%和52.94%,但在100%和80% WHC,60%和40% WHC条件之间幼苗叶片数无显著差异。与100% WHC 相比,60%和40% WHC条件下石茅的相对叶绿素含量分别显著降低了53.24%和68.79%。但100%和80% WHC 条件之间无显著差异。

表2 不同土壤含水量条件下石茅种幼苗的形态特征及生物量特征Table 2 Morphological characteristics and biomass characteristics of Sorghum halepenseseedlings
不同水分条件下石茅幼苗的地上生物量、地下生物量、总生物量和根冠比均有明显差异。随着水分含量的降低石茅幼苗的地上生物量和总生物量都随之减少,与100% WHC 条件下相比,60%和40% WHC 条件下石茅幼苗的地上生物量分别显著减少50.73%和61.65% (P< 0.05),总生物量分别显著减少28.40%和32.07%。随水分含量的降低石茅幼苗的地下生物量和根冠比都随之增加(表2),与100% WHC 条件下相比,60%和40% WHC 条件下石茅幼苗的地下生物量分别显著增加49.93%和71.74%,根冠比分别显著增加196.77 和316.13%。
3 讨论
高萌发率是入侵植物能成功入侵并迅速扩散的关键。水分是影响杂草种子萌发及幼苗生长的关键生态因素之一,在干旱胁迫下种子能否萌发对幼苗生存和生长至关重要[8]。研究干旱胁迫条件下种子萌发特性是了解植物响应干旱逆境的第一步。本研究显示,在2.5% PEG 浓度下,石茅种子的发芽率达到最大值。当干旱胁迫超过一定程度(7.5% PEG),石茅种子的发芽率、发芽指数和活力指数随干旱胁迫的加剧而呈现下降趋势。这一结果与5% PEG 胁迫下有助于北美车前草(Plantago virginica)[21]、扭黄毛(Heteropogon contortus)[22]、中华羊茅(Festuca sinensis)[23]的种子萌发,超过种子对干旱的耐受度时,种子萌发率受到强烈抑制相类似。在15% PEG 浓度下石茅种子的种子仍能萌发,说明石茅的种子具有适应干旱生境的能力。在种子萌发过程中,胚根和胚芽长度是反映早期幼苗生长的关键指标[24-25],随PEG浓度增加,石茅的胚根和胚芽长度都减少了,干旱胁迫对石茅的胚根有明显的抑制作用。低浓度PEG(2.5%~5%)胁迫对胚芽的抑制作用不明显;高浓度PEG (10%~15%)胁迫下的胚根/胚芽均显著高于低浓度处理,说明重度干旱胁迫对石茅胚芽抑制作用要强于胚根,这可能是水分缺乏时石茅适应环境的一种表现。因此,在轻度干旱胁迫下,石茅种子可以顺利萌发并完成幼苗的形态建成;在中度和重度干旱胁迫下石茅种子表现出较强的生态适应性,说明石茅种子萌发所需的土壤水分生态位广泛,无论是干旱的荒地或是湿润的农田都可能成为其潜在的分布区域。
入侵种对环境具有较高的耐受性,能通过自身形态特征及内在生理机制的调节来适应逆境[12,26]。土壤含水量是决定了入侵植物的生存关键因素之一[27-28]。在水分胁迫的条件下,植物的形态特征及生理生化指标将发生一系列适应性改变,如会出现生长缓慢、株高、叶片数量、比叶面积、叶绿素相对含量的降低[25,29],且水分亏缺越严重,其降低程度越大等现象。盆栽试验结果显示,随着干旱胁迫的增强,石茅幼苗的株高、叶片数、相对叶绿素含量、比叶面积、地上生物量、总生物量均有下降,但其地下生物量及根冠比则显著增加了。且与100% WHC条件下相比,在80% WHC 条件下石茅的这些形态指标及生物量指标均没有显著差异,说明80% WHC条件下并不会限制石茅幼苗的生长。随着干旱胁迫的增强,石茅的株高及叶片数随之减少,这可能是其通过株高及叶片数的降低,来减少植物水分的蒸腾速率,以维持植物体内水分平衡,来适应干旱逆境[29-30]。水分胁迫下叶绿素含量的变化可以反映植物对光能的捕获能力,从而影响光合产物的积累,最终影响植株的生长[31]。干旱胁迫条件下石茅的叶绿素相对含量显著降低,这与董守坤等[32]发现随着干旱胁迫强度的增加,春大豆(Glycine max)叶绿素含量显著降低;王伟等[33]发现不同生育期不同水分亏缺程度均引起冬小麦(Triticum aestivum)株高、叶绿素相对含量的降低的研究结果一致。石茅的地上生物量及总生物量随干旱胁迫强度的增加而减少,这可能是因为水分的缺乏,影响到光合作用,导致同化产物合成减少,进而影响了干物质的积累和分配。同时,干旱促使石茅干物质分配发生了改变,随着干旱胁迫强度的增加,生物量优先向根系分配,根冠比增大,以适应干旱逆境。因此,轻度干旱胁迫并没有显著影响石茅幼苗的生长,中重度干旱胁迫条件下,石茅也通过减少叶片数量、降低蒸腾作用、调整物质分配、增加根冠比等措施来应对,表现出较强的适应性,这进一步提高了其竞争生长能力,加重了其危害性。
4 结论
综上所述,轻度干旱(2.5%~5% PEG)胁迫对外来入侵植物石茅种子萌发没有抑制作用。重度干旱(10%~15% PEG)条件下,其通过调节胚根/胚芽比来适应干旱环境,因此,石茅种子在干旱条件下也有萌发的可能。当土壤含水量小于60% WHC时,幼苗期的石茅可通过减少株高、叶片数、相对叶绿素含量、地上生物量及总生物量,增加地下生物量和根冠比等形态及生理变化对干旱胁迫条件做出响应并表现出一定的耐受及适应能力。可见,轻度干旱胁迫不能抑制石茅种子的萌发及其幼苗的生长,石茅可以通过种子萌发及幼苗生长策略应对生境中可能出现的干旱胁迫环境,从而提高在逆境中的生长及竞争优势,这可能是石茅种群能在海南岛干旱地区迅速扩散并成灾的重要原因之一。
猜你喜欢
广西林业科学(2021年6期)2022-01-21 07:39:28
山西农业科学(2021年8期)2021-08-17 06:20:10
青年文学家(2021年4期)2021-03-18 07:00:23
农民致富之友(2020年19期)2020-07-18 15:32:39
山西农业科学(2020年1期)2020-02-27 17:00:54
家庭百事通·健康一点通(2019年11期)2019-09-26 09:27:52
种子科技(2018年11期)2018-09-10 00:56:48
许昌学院学报(2018年8期)2018-09-05 02:05:18
天津农林科技(2015年1期)2015-12-30 13:05:14
饮食科学(2014年8期)2014-08-22 17:42:07
