
长江流域外来鲟入侵风险评估、适生区预测及影响因素分析*
2023-09-01 04:49:52李奥璞范俊韬王艺霖
湖泊科学 2023年5期
李奥璞,范俊韬**,钱 锋,2,王 瑜,2,王艺霖,时 悦
(1:中国环境科学研究院环境基准与风险评估国家重点实验室,北京 100012) (2:中国环境科学研究院水生态环境研究所,北京 100012)
长江作为我国第一、世界第三大河流,全长6300 km,流域总面积约为1.8 × 106km2[1]。流域内的复杂地形、发达水系和多样气候造就了丰富的鱼类资源[2-3]。历史上,长江流域存在着白鲟(Psephurusgladius)、中华鲟(Acipensersinensis)以及长江鲟(Acipenserdabryanus)等3种本地鲟[4]。得益于当时较好的生态环境,流域内鲟资源丰富。近年来,由于过度捕捞和河流连通性显著降低等多种因素影响,长江流域本地鲟资源状况急转直下:白鲟已灭绝[5],长江鲟被世界自然保护联盟(IUCN)宣告野外灭绝[5],中华鲟在葛洲坝建成之后出现了明显的产卵异常,2017-2020年连续4年无自然产卵记录[6],种群衰退严重,被IUCN红色物种名录列为极危(critically endangered)状态。
与此同时,随着我国鲟养殖规模不断扩大,长江流域养殖的非长江流域本地鲟(简称为外来鲟)种类不断增多,数量也不断扩大[7]。养殖品种主要为大杂交鲟(达氏鳇(Husodauricus)(♀)×施氏鲟(Acipenserschrenckii)(♂))、西杂鲟(西伯利亚鲟(Acipenserbaerii)(♀)×施氏鲟(Acipenserschrenckii)(♂))、小鲟鳇(欧洲鳇(Husohuso)(♀) × 小体鲟(Acipenserruthenus)(♂))、西伯利亚鲟(Acipenserbaerii)、欧洲鳇(Husohuso)、俄罗斯鲟(Acipensergueldenstaedtii)、匙吻鲟(Polyodonspathula)、达氏鳇(Husodauricus)、施氏鲟(Acipenserschrenckii)和小体鲟(Acipenserruthenus)等[8-11],通常被用作商品售卖。在上述10种主要养殖外来鲟中,虽然纯种鲟均被IUCN红色物种名录列为易危(vulnerable)、濒危(endangered)和极危状态[4],但是根据世界鱼类数据库(FishBase, https://www.fishbase.de/)以及已有资料[12-13],西伯利亚鲟已被列为德国和奥地利的入侵物种[14],匙吻鲟已被列为白俄罗斯的入侵物种[15]。
养殖逃逸事件[16]是导致外来鲟入侵至长江流域的主要原因。例如2016年湖北省隔河岩和高坝洲库区养殖的外来鲟发生大规模逃逸[16],共计近万吨外来鲟进入长江流域,而且逃逸发生的地点靠近湖北宜昌中华鲟自然保护区。近年来,外来鲟逃逸的潜在生态影响成为研究热点。研究表明,养殖鲟逃逸带来的生物入侵是一个威胁长江本地鲟生存的潜在因素[17]。除了在逃逸时可能携带容易使本地鲟患病的病原体[16],数量众多的外来鲟还可能和数量稀少的本地鲟在食物和栖息地上发生不对称竞争[17-18],影响本地鲟生存。因此,评估外来鲟可能产生的影响[16-17],避免外来鲟可能对本土鲟产生的危害,是长江本地鲟保护工作的重点之一。然而,目前对于外来鲟的入侵风险和适生区缺乏相应的量化,不利于为本土鲟保护提供完善的资料。
基于上述情况,本文采用水生物种入侵性筛查工具(aquatic species invasiveness screening kit, AS-ISK)评估外来鲟的入侵风险,筛选出入侵风险最高的鲟,进而使用最大熵建模(maximum entropy modeling),对入侵风险最高的鲟在长江流域当前和未来时期(2021-2040年)的适生区进行预测,并对适生区的面积、区域分布、质心迁移以及影响因素进行分析,以期为防范长江流域外来物种入侵提供参考。
1 材料与方法
1.1 研究区域概况
长江流域是我国第一大流域,含多种气候与生境类型,具有湖北宜昌中华鲟自然保护区和上海长江口中华鲟自然保护区等珍稀鱼类的保护区(图1)。在“长江大保护”政策实施前,流域内外来鲟养殖业发达,曾有两次严重的外来鲟逃逸事件,分别为2016年湖北清江养殖鲟逃逸事件以及2020年甘肃文县养殖鲟逃逸事件[16,19]。
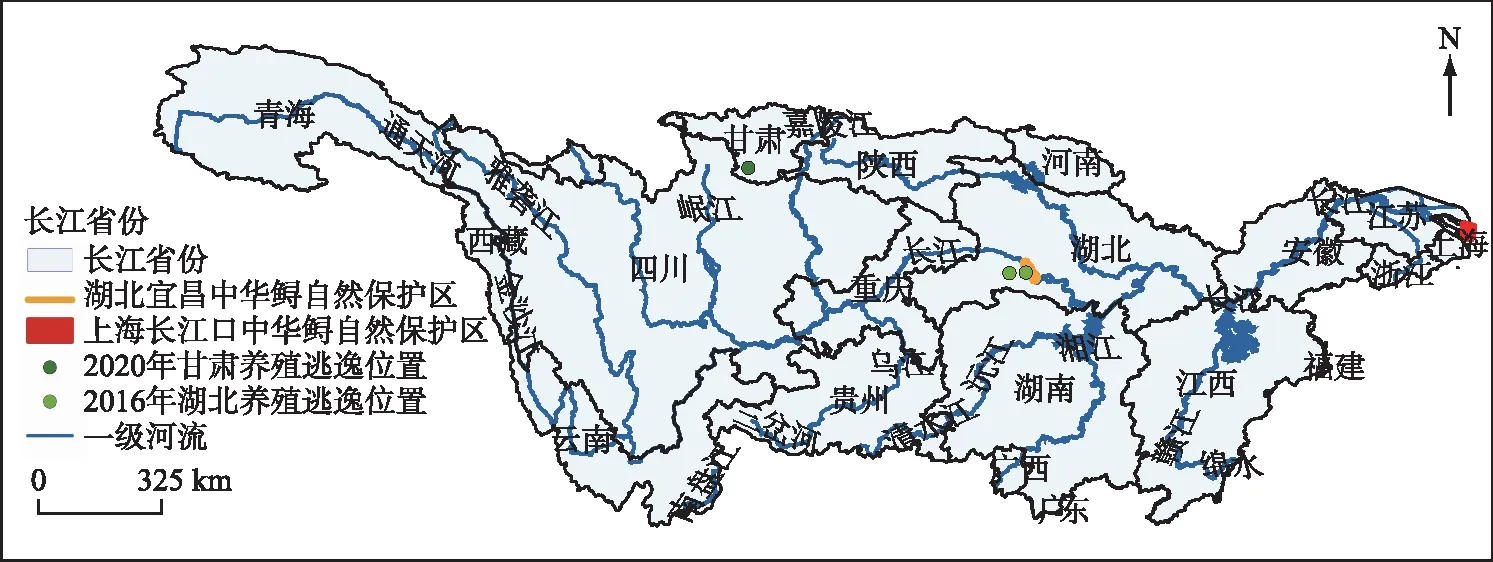
图1 研究区域Fig.1 Study area
1.2 外来鲟入侵风险评估
本文使用水生物种入侵性筛查工具评估外来鲟的入侵风险[20]。该工具在评估全球120个地区的800余种外来水生生物的潜在入侵风险时,取得了较好效果[21-22]。
水生物种入侵性筛查工具的前49道题为基础风险评估项(basic risk assessment, BRA),后6道题为气候变化项(climate change assessment, CCA)。问题共9个部分,其中第2和第3部分在问题和得分上存在关联;各部分名称和得分范围为[22]:(1)驯化/栽培(得分范围-2~4分);(2)气候/分布及引入风险;(3)是否在别处入侵(与第2部分得分总和的范围为-1~20分);(4)不受欢迎(或续存)的特征(得分范围-1~12分);(5)资源开采(得分范围0~7分);(6)繁殖(得分范围-4~7分);(7)扩散机制(得分范围-6~9分);(8)耐受性特征(得分范围-6~9分);(9)气候变化(得分范围-12~12分)。
评估者回答问题时,需列出依据和答案置信度(分4个等级,低=1,中=2,高=3,非常高=4)。最终得分被汇总为BRA项分数和BRA+CCA项分数(得分范围为-32~80)。分数体现了物种入侵的能力以及利于入侵的优势条件[22-23];分数<1时,为低或无入侵风险[22];分数≥1即有入侵风险,且分数越高,入侵风险越高[23]。置信度被汇总为BRA项置信因子、CCA项置信因子和BRA+CCA项置信因子,其中BRA+CCA项置信因子计算公式为[22]:
(1)
式中,CF为置信因子,CLQi为第i个问题的置信度。
评估所需的资料来自FishBase、国际应用生物科学中心数据库(CABI, https://www.cabi.org/isc/)、全球入侵物种数据库(GISD, http://www.iucngisd.org/gisd/)和文献资料[4,24]等。因为外来鲟主要是底层肉食性鱼类[4],还包含原产于我国其他地区的鲟,本文为精确地评估外来鲟入侵风险,并使评估物种数达到15种,满足计算阈值的要求[25],添加了流域内常见的5种与外来鲟在食物或者栖息水层上的生态需求相似的外来鱼类作参考,即常与外来鲟一起养殖的虹鳟(Oncorhynchusmykiss)、原产于长江流域以北的丁鱥(Tincatinca)[12]、原产于长江流域以南的鲮(Cirrhinusmolitorella)[26]、流域内广泛养殖的斑点叉尾鮰(Ictaluruspunctatus)[27]和尼罗罗非鱼(Oreochromisniloticus)[28]。
本文根据BRA和BRA+CCA项分数,以及鱼类是否被列为入侵物种(作为先验分类变量),绘制受试者工作特征曲线(receiver operating characteristic curve, ROC);根据ROC的线下面积(area under the curve,AUC),来衡量评估质量。依据文献[21],AUC越接近1,表明对物种入侵性的评估越准确。同时,根据ROC的灵敏性(Sensitivity)与特异性(Specificity),计算约登指数(Youden’sindex),公式为:
Youden’sindex=Sensitivity+Specificity-1
(2)
约登指数最大时对应的分数,具有最高的真阳性和最低的假阴性可能[22]。参照相关研究[29],本文计算BRA项中约登指数最大时对应的分数,将其作为划分BRA项和BRA+CCA项的中高入侵风险的阈值。依据有关研究[21-23],高入侵风险的物种入侵能力和有利于入侵的优势较多,需要重视和优先管理;中入侵风险的物种有一定的入侵能力和有利于入侵的优势,需要持续观察;低入侵风险的物种入侵可能性较低。
为精确分析外来鲟在各部分得分情况,本文根据外来鲟在各部分的得分计算得分率,绘制雷达图;在其中将问题和得分存在关联性的第2和第3部分合并计算得分率。同时,在图中标绘出每部分得分为0时的得分率和5种相关鱼类在各部分得分率的均值。
1.3 外来鲟适生区预测
本文使用最大熵建模预测入侵风险最高的外来鲟在长江流域当前和未来时期的适生区,并分析影响因素;使用ArcGIS处理预测结果,划分适生区等级和计算各级适生区面积,结合RStudio的“SDMTools”和“geosphere”包,对于适生区的分布和质心迁移进行分析。
1.3.1 环境变量筛选和建模参数设置 建模所需的外来鲟分布点来自全球生物多样性信息网络(GBIF, https://www.gbif.org/)[30]以及文献[31-35],筛去重复记录,共获得136个分布点。为防止分布点距离过近而存在空间自相关[36],使用RStudio的“spThin”包对两个距离小于1 km的点进行随机筛选,共获得103个分布点。所需的矢量地理数据集(全球国家行政边界数据、2015年中国省级行政边界数据、中国一级河流空间分布数据集和2015年中国地市行政边界数据)和栅格地理数据集(2020年中国土地利用遥感监测数据),来自中国科学院资源环境科学与数据中心(https://www.resdc.cn/)。
建模使用的环境变量为生物气候变量和海拔。气候中的极端温度和降水等条件对鱼类生存有明显的影响,而海拔与溶氧等因素联系,可能对于鱼类分布有着显著影响[37]。本文所需的当前和未来时期(2021-2040年)的19个生物气候变量和1个海拔地理数据集来自WorldClim网站[38-39],空间分辨率为2.5 arc-minutes。19个生物气候变量为:年平均温度(BIO1)、昼夜温差的月平均值(BIO2)、等温性(BIO3)、温度季节性变异系数(BIO4)、最热月份最高温度(BIO5)、最冷月份最低温度(BIO6)、温度年变化范围(BIO7)、最潮湿季的平均温度(BIO8)、最干燥季的平均温度(BIO9)、最热季的平均温度(BIO10)、最冷季的平均温度(BIO11)、年降水量(BIO12)、最潮湿月份的降水量(BIO13)、最干燥月份的降水量(BIO14)、降水季节性变异系数(BIO15)、最潮湿季的降水量(BIO16)、最干燥季的降水量(BIO17)、最热季降水量(BIO18)和最冷季的降水量(BIO19)。未来时期的生物气候变量来自CMIP6中3种气候模式(BCC-CSM2-MR、CNRM-ESM2-1和MIROC6模式)的预测结果[40-43]。每种模式含4种共享社会经济路径(shared socioeconomic pathway, SSP)和典型浓度路径(representative concentration pathway, RCP)的组合情景;4种情景分别为:SSP126(低社会脆弱性、低减缓压力和低辐射强迫)、SSP245(中等社会脆弱性与中等辐射强迫)、SSP370(相对高的社会脆弱性与相对高的辐射强迫)和SSP585(高社会脆弱性和高辐射强迫)[43-44]。每种模式根据不同的未来社会发展和辐射强迫情况,在4种情景下分别预测了对应的未来时期气候[36,42]。
为防止生物气候变量间存在多重共线性和空间相关性影响建模精度[45],本文使用ArcGIS提取分布点的19个当前生物气候变量的数值;使用方差膨胀因子(variance inflation factor,VIF)表征变量的多重共线性,当其小于10 时,变量间多重共线性将大大降低[46-47],使用Pearson相关系数|r|表征两个变量的空间相关性,当|r|≥ 0.8时,表明相关性较高,需要筛选[48]。同时,分析19个当前生物气候变量对于物种在原产地分布的影响,以贡献率表征影响大小;贡献率相同时,根据置换重要性表征影响大小。变量筛选时,首先依据资料[4,49]提供的匙吻鲟生物学特性,参考文献[50]所筛选的变量,选择出具有较强实际生物学意义的必选变量。其次,选择出VIF小于10的变量,筛去其中和必选变量相关性较高的(Pearson相关系数|r|≥0.8)。余下的VIF小于10的变量中,如果两个变量间Pearson相关系数|r|≥ 0.8,选择对于物种在原产地分布影响较大的变量。最终,获得参与建模的变量。未来时期的3种气候模式的4种情景下,使用挑选的当前生物气候变量对应的未来时期生物气候变量,参与适生区预测。在预测未来时期适生区时,假定海拔在未来时期不变。
建模运行前,将分布点、原分布区与长江流域的环境变量地理数据集导入最大熵建模。参考相关研究[45],将随机抽取的75%的位点作为训练集,余下25%的位点作为测试集;使用偏差文件(Bias file)功能以优化建模[51];将建模阈值规则设置为最大化训练集的灵敏性和特异性之和[51];其余参数保持默认,运行建模,输出结果为物种适宜概率。
1.3.2 预测结果处理 参考文献[48],本文依据ArcGIS的“自然间断点分级法”,将所得当前时期适生区分为非适生区(P<0.03)、低适生区(Ⅰ级适生区,0.03≤P<0.09)、中适生区(Ⅱ级适生区,0.09≤P<0.16)和高适生区(Ⅲ级适生区,P≥0.16)。同时依据2020年中国土地利用遥感监测数据,提取长江流域内的河渠、湖泊和水库坑塘等水体,以计算各级适生区的(水域)面积。为减少各气候模式对未来气候状况预测的不确定性,降低其对适生区预测的影响,本文参考文献[41],使用ArcGIS的局部分析工具,计算同一情景下,根据3个气候模式的生物气候变量和海拔分别预测出的物种适宜概率均值,将其作为该情景的最终预测结果;使用与当前时期同样的划分标准,进行适生区分级,并且假定流域内水域面积在未来时期不出现明显变化,进而根据2020年中国土地利用遥感监测数据来计算各级适生区(水域)面积。同时,对于当前和未来时期的适生区,使用RStudio的“SDMTools”包,分析适生区的质心位置;使用RStudio的“geosphere”包,分析适生区质心在未来时期的迁移。
2 结果与分析
2.1 外来鲟入侵风险评估结果
2.1.1 外来鲟评估质量 结果中,BRA项置信因子为0.610±0.051(mean ± sd,下同),CCA项置信因子为0.535±0.038,BRA+CCA项的置信因子为0.601±0.047,表明可信度处于中上水平。结果ROC中BRA项AUC为0.954,BRA+CCA项AUC为0.926,均大于0.9,表明评价效果良好,准确度较高。
2.1.2 外来鲟入侵评分和风险等级 依据公式(2)计算可得,阈值为4分。依据阈值,BRA项和BRA+CCA项中,<1分为低入侵风险,1~4分是中风险,>4分是高风险。BRA项中,西伯利亚鲟、匙吻鲟、虹鳟、斑点叉尾鮰、尼罗罗非鱼、鲮和丁鱥为高入侵风险,施氏鲟和欧洲鳇为中入侵风险,其余为低入侵风险。BRA+CCA项中,匙吻鲟、虹鳟、斑点叉尾鮰、尼罗罗非鱼、鲮和丁鱥为高入侵风险,西伯利亚鲟为中入侵风险,其余为低入侵风险。在外来鲟中,匙吻鲟的BRA项得分为9分,BRA+CCA项得分为13分,得分均最高,表明在评估的外来鲟中,其具有最高的入侵风险(附表I)。
外来鲟主要得分部分有:驯化/栽培部分,外来鲟得分率大多超过60%,获得2分;不受欢迎(或续存)的特征部分,大多数外来鲟得分率为23%,获得2分;资源开采部分,大部分外来鲟得分率超过70%,获得5分。表明外来鲟在这些部分具有较多有利于入侵的优势特性。匙吻鲟额外得分较高部分有:气候/分布及引入风险部分和是否在别处入侵部分,得分率为38%,获得7分;气候变化部分,得分率为67%,获得4分。表明匙吻鲟在这些部分具有更多有利于入侵的优势。外来鲟得分较低部分为:繁殖部分,大多数外来鲟得分率为9%,获得-3分;扩散机制部分,大多数外来鲟得分率为33%,获得-1分;气候变化部分,大多数外来鲟得分率为33%,获得-4分。说明外来鲟在上述部分具有较少利于入侵的优势。此外,外来鲟在耐受性特征部分得分差异较大,最高获得-1分,最低获得-6分,说明外来鲟各品种间与入侵相关的耐受性特征差异较大。相比之下,参考鱼类在所有的部分实际得分均较高,在各部分均具较多有利于入侵的优势,其中在驯化/栽培和气候变化部分得分率超过80%(图2)。
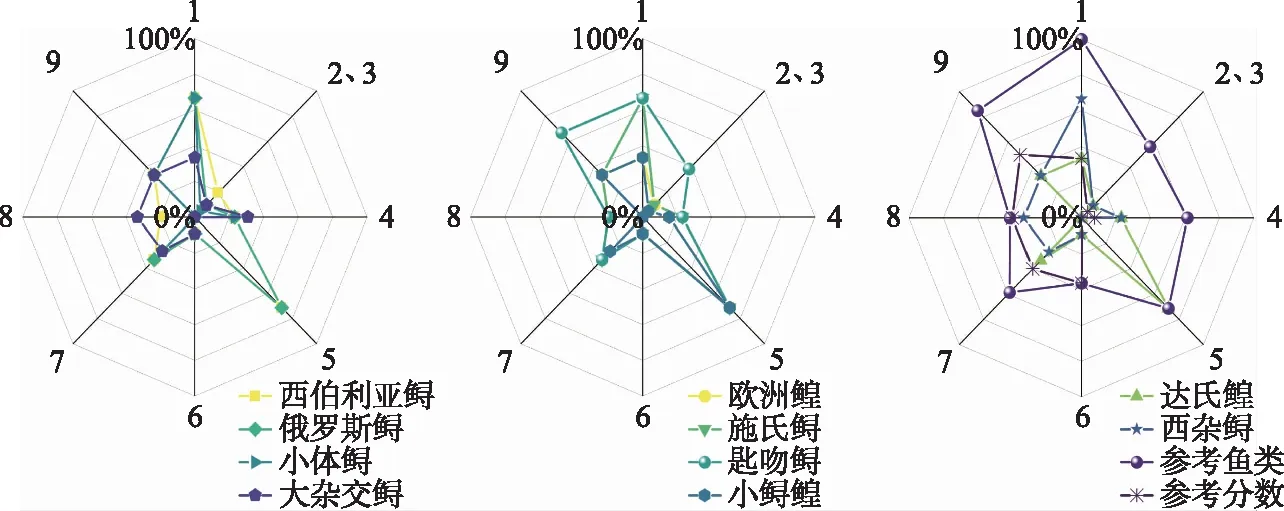
图2 外来鲟在各部分得分率(1:驯化/栽培; 2:气候/分布及引入风险; 3:是否在别处入侵; 4:不受欢迎(或续存)的特征;5:资源开采; 6:繁殖; 7:扩散机制; 8:耐受性特征; 9:气候变化)Fig.2 The scoring rate of exotic sturgeon and paddlefish in each part(1: Domestication/cultivation; 2: Climate/distribution and introduction risk; 3: Invasive elsewhere; 4: Undesirable (or persistence) traits; 5: Resource exploitation; 6: Reproduction; 7: Dispersal mechanisms; 8: Tolerance attributes; 9: Climate change)
2.2 匙吻鲟适生区预测结果
因为匙吻鲟在10种外来鲟中得分最高,具有最高入侵风险,所以对其在长江流域的适生区进行预测。
2.2.1 环境变量筛选结果 温度变量中,最热月份最高温度(BIO5)和最冷月份最低温度(BIO6)与匙吻鲟耐受性特征紧密相关,对于分布具有重要影响;降水变量中,年降水量(BIO12)和降水季节性变异系数(BIO15)对于匙吻鲟栖息地影响较多,对物种生存具有重要的影响。本文将BIO5、BIO6、BIO12和BIO15作为具有较强实际生物学意义的必选变量。
本文筛选出VIF小于10的生物气候变量为:BIO2、BIO4、BIO5、BIO8、BIO9、BIO13、BIO15和BIO18。由建模分析出VIF小于10的变量对于匙吻鲟在原产地分布的影响从大到小依次为:BIO4、BIO5、BIO2、BIO18、BIO8、BIO15、BIO13、BIO9(图3)。

图3 生物气候变量Pearson相关系数图(VIF<10的变量为红色)Fig.3 Pearson correlation coefficient plot of bioclimatic variables (variables with VIF<10 shown as red)
综上,最终选择参与预测适生区的生物气候变量为:BIO2、BIO5、BIO6、BIO8、BIO12、BIO15和BIO18。
2.2.2 适生区预测结果 最大熵建模预测当前适生区时,ROC中,训练集AUC为0.992,测试集AUC为 0.991,均大于0.9。在预测未来时期的适生区时,4种情景下,训练集AUC和测试集AUC也均大于0.9。上述结果表示建模效果较好,结果可信。
依据2020年中国土地利用遥感监测数据,流域内的河渠、湖泊和水库坑塘等水域的面积约为42479.0 km2。当前时期,匙吻鲟Ⅲ级适生区面积为2996.8 km2,区域内长江干流长约210 km;Ⅱ级适生区面积为8386.2 km2;Ⅰ级适生区水域面积为13000.3 km2。适生区主要包括湖南、湖北、安徽和江西等省的部分地区(图4)。根据2015年中国地市行政边界数据,分析Ⅲ级适生区主要包含的市级行政单位发现,九江市东部、安庆市南部、上饶市西部、宜春市东部和鹰潭市北部的Ⅲ级适生区面积较大(附表Ⅱ)。
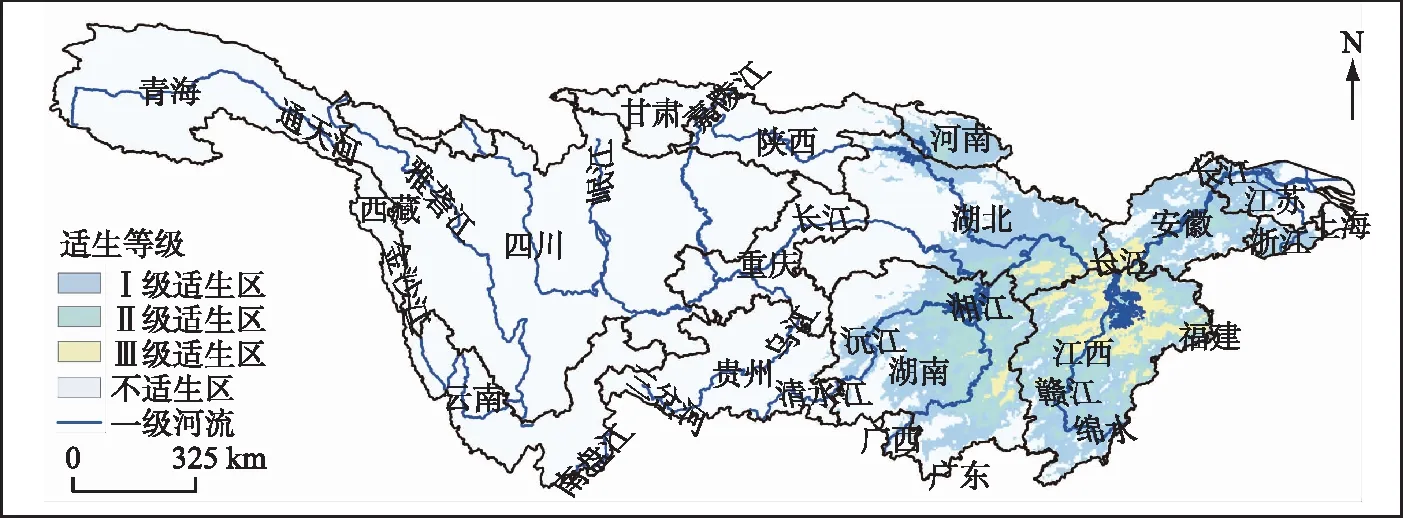
图4 匙吻鲟当前适生区Fig.4 Contemporary suitability regions of the paddlefish
未来时期,在SSP126、SSP245、SSP370和SSP585情景中,社会脆弱性和辐射强迫依次增加,气候受干扰程度逐渐增加,相应的适生区变化程度不同(图5)。同当前时期相比,SSP126情景下,Ⅲ级适生区内干流长度增加约1.53倍,Ⅲ级适生区面积增加约1.41倍,总适生区面积增加约0.27倍;SSP245情景下,Ⅲ级适生区内干流长度增加约2.62倍,Ⅲ级适生区面积增加约1.72倍,总适生区面积增加约0.28倍;SSP370情景下,Ⅲ级适生区内干流长度增加约2.47倍,Ⅲ级适生区面积增加约1.78倍,总适生区面积增加约0.28倍;SSP585情景下,Ⅲ级适生区内干流长度增加约2.18倍,Ⅲ级适生区面积增加约1.74倍,总适生区面积增加约0.24倍(附表Ⅲ)。对于整体的适生区格局来说,本文使用RStudio的“SDMTools”和“geosphere”包进行质心分析,发现当前时期适生区质心位于江西省九江市西北部。在未来时期的4种情景下,适生区质心均向东北方向的湖北省黄石市和黄冈市交界处移动,质心迁移距离依次为:SSP126情景下,迁移142.90 km;SSP245情景下,迁移150.62 km;SSP370情景下,迁移163.20 km;SSP585情景下,迁移137.97 km。

图5 匙吻鲟未来时期适生区Fig.5 Future suitability regions of the paddlefish
2.2.3 适生区影响因素分析 最大熵建模结果表明,在当前适生区预测中,BIO18的贡献率为18.9%,置换重要性为23.0%;BIO12的贡献率为17.3%,置换重要性为2.0%;BIO5的贡献率为16.3%,置换重要性为44.8%;BIO6的贡献率为15.8%,置换重要性为18.4%;BIO2的贡献率为12.9%,置换重要性为1.1%;BIO15的贡献率为10.9%,置换重要性为9.6%。上述变量贡献率总和为92.1%,置换重要性总和为98.9%,是影响匙吻鲟在长江流域分布的主要变量,响应曲线如图6所示。
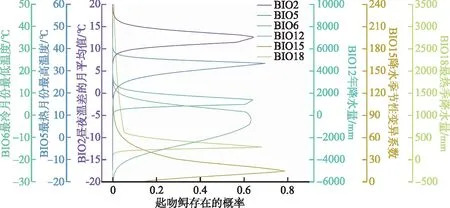
图6 主要变量响应曲线Fig.6 Response curves of main variables
由图6可知,所有主要变量对于匙吻鲟分布的影响都表现为不同程度的单峰曲线,即有一个最适宜匙吻鲟分布的区间。BIO18(最热季降水量)为300~500 mm时,匙吻鲟分布的概率较高。BIO12(年降水量)为1000~1400 mm时,匙吻鲟分布的概率较高。BIO5(最热月份最高温度)为28~34℃时,匙吻鲟分布的概率较高。BIO6(最冷月份最低温度)为-3~3℃时,匙吻鲟分布的概率较高。BIO2(昼夜温差的月平均值)为11~14℃时,匙吻鲟分布的概率较高。BIO15(降水季节性变异系数)为15~25时,匙吻鲟分布的概率较高。
在未来时期气候模式的4种情景中,上述变量依然对匙吻鲟的分布起到主导性作用。
3 讨论
3.1 外来鲟的入侵性
近年来,本土鲟种群生存恶化的情况突出表现为产卵行为减少,甚至消失不见[5],这主要由水体连通性降低导致适宜产卵场和亲本性腺发育所需的适宜环境减少,繁殖条件难以达到造成[1,4]。对于外来鲟来说,繁殖所需条件虽然存在一定差异,但是均较为复杂,外来鲟在原产地的繁殖均在不同程度上受到水体连通性降低的消极影响[4]。进入流域后,外来鲟的繁殖迁移可能同样受水坝的消极影响,相应的以繁殖体和幼体为主体的扩散出现可能性也较低,这在一定程度上限制了其扩散。因此,外来鲟在水生物种入侵性筛查工具评估的繁殖部分和扩散机制部分得分较低。同时,外来鲟对于水质要求高,对于化学污染敏感,对干扰的耐受能力较差[4],这些耐受性特征不利于入侵过程,使得外来鲟在耐受性特征部分得分普遍小于0;而外来鲟中一些洄游性鲟和杂交鲟可以耐受一定盐度[4],部分杂交鲟还能耐受长时间干露[9],比其他外来鲟具有更多优势,使外来鲟在耐受性特征部分得分差异较大。
依据图2和各部分分数设置,外来鲟主要得分部分为驯化/栽培、不受欢迎(或续存)的特征以及资源开采部分。驯化/栽培部分评估了外来鲟的驯化历史、近亲物种的入侵性和物种利用形式,因为近亲种中的西伯利亚鲟[14]和匙吻鲟[15]被列入侵物种,并且外来鲟中的纯种鲟均被IUCN红色物种名录列为易危、濒危或者极危状态[4],在野外捕获后,很有可能以活体形式被保护(利用)下来,所以外来鲟大多在该部分得分高。不受欢迎(或续存)的特征部分考虑了外来鲟对于生境适应性和可能的影响。因为大多数外来鲟有一定的洄游性[4]和游泳能力[52],所以可耐受的流速范围较广。除此之外,鲟存在着一些共患病[9],外来鲟可能作为相关病原体传播的媒介[16],所以大多数外来鲟在此部分得分较高。资源开采部分评估了外来鲟利用评估区资源的可能性,外来鲟多为肉食性,食物包括鮈等小型鱼类[4];即便是主食浮游动物的匙吻鲟,也具有肉食性鱼类的消化道结构特征[49],存在取食黄黝鱼等小型鱼类的记录[53]。因此,外来鲟大多具有捕食小型濒危鱼类的特点,从而在此部分得分较高。
另外,匙吻鲟在气候/分布及引入风险、是否在别处入侵和气候变化部分得分高。气候/分布及引入风险部分和是否在别处入侵部分评估了外来鲟当前的分布与入侵情况,匙吻鲟原分布区和长江流域气候相似度高,并且已被列为白俄罗斯的入侵物种[15],所以得分较高。对于气候变化部分,根据预测,未来长江流域主要有两种气候变化:一是降水增加[54],二是温度缓慢的上升[55]。降水增加,导致径流增加,可能使流域内原有的外来鲟扩散增加;还可能伴随着升高的温度带来洪水等灾害,使外来鲟从陆上养殖区被动扩散进入流域内。只是大多数外来鲟原产地纬度较高,最适生长温度较低,对升温耐受性差,温度升高,建群风险将降低。例如西伯利亚鲟生长最适温度为18~22℃,水温超出最适范围,摄食强度将明显下降[56-57];其在夏季繁殖时所需水温为9~18℃[56],水温低于或高于所需温度,繁殖活动将受抑制。相比之下,匙吻鲟原产地纬度较低,适温范围为0~37℃,最适生长温度为15~25℃,且对极端温度具有较强的耐受性[49]。但是繁殖所需条件较为复杂[4],气候变化后,其在流域内的繁殖仍可能受到水坝的阻碍,因此建群风险不变。因为建群风险的变化,大多数外来鲟对生态系统的影响减小,而匙吻鲟的影响保持不变,所以匙吻鲟在此部分得分较高。
综上,虽然大多数外来鲟有近亲种被列为入侵物种以及在流域内存在的记录,并且具有一定的适应生境和影响流域内生物的能力,但是在流域内繁殖和扩散的能力较差,对于干扰的耐受能力较差,所以被评为中低入侵风险。匙吻鲟因为原产地和流域内气候相似度高,具有在其他区域的入侵历史,并且在应对气候变化时比其他外来鲟更有优势,所以在BRA项和BRA+CCA项得分均最高,均为高入侵风险。此外,之前的养殖逃逸[16]还使得匙吻鲟在流域内个体数量较多、分布较广,匙吻鲟可以通过竞争等方式对本地鲟种群产生消极影响;并且参考相关研究[21-23],在AS-ISK评估中被列为高入侵风险的外来鱼类需要被重视和优先管理,因此在当前和未来时期,匙吻鲟的相关影响应该被重视。
3.2 匙吻鲟的适生区
由图4可知,当前时期匙吻鲟的Ⅲ级适生区和Ⅱ级适生区主要位于长江流域中下游地区。根据Köppen-Geiger气候分类法[24],上述地区主要属于温暖带常湿温暖气候类型;在原产地,匙吻鲟的分布区主要也是温暖带常湿温暖气候和部分冷温带常湿冷温气候的气候类型,这间接证实了预测具有较高的准确性。
当前时期,降水和温度对于匙吻鲟的分布具有重要影响。降水变量中,BIO18(最热季降水量)、BIO12(年降水量)和BIO15(降水季节性变异系数)对于维持匙吻鲟的栖息地十分重要,影响着栖息地的水环境条件。温度变量中,BIO5(最热月份最高温度)、BIO6(最冷月份最低温度)以及BIO2(昼夜温差的月平均值)对于匙吻鲟的生长发育和分布范围具有关键影响。其中,BIO5具有最高的置换重要性,在其为28~34℃时,匙吻鲟分布的概率较高,这体现了匙吻鲟原分布区的环境特征。研究表明,匙吻鲟在原产地常经历夏季高水温的情况,对于高水温具有一定的耐受性[49],80日龄的匙吻鲟可耐受的最高水温为35.2℃,且耐受能力随年龄增长而增加[58-59]。但是在高温下,其生长速度不高,还存在生长受抑制的情况[59]。与此同时,为了适应夏季高温,匙吻鲟发展了适应性行为,倾向于在温度较高的白天进入较深的水层来躲避高水温[49,60]。
未来时期,社会发展对于环境的影响和辐射强迫等变化将导致长江流域的降水和温度缓慢增加趋势[54-55]。未来时期气候模式的4种情景因为假定条件不同,19个生物气候变量对于大趋势的响应也不同,主要体现为影响匙吻鲟分布的重要变量BIO18(最热季降水量)和BIO5(最热月份最高温度)在4种情景变化不同。根据WorldClim中2021至2040年的4种情景的气候在3个模式(BCC-CSM2-MR气候模式、CNRM-ESM2-1气候模式和MIROC6气候模式)的平均状态,流域内BIO18(最热季降水量)的均值在4种情景依次增加:在SSP126情景下,增加1.25 mm;在SSP245情景下,增加2.46 mm;在SSP370情景下,增加3.43 mm;在SSP585情景下,增加7.04 mm。流域内BIO5(最热月份最高温度)的均值在各情景的增幅不同:在SSP126情景下,增加1.44℃;在SSP245情景下,增加1.40℃;在SSP370情景下,增加1.25℃;在SSP585情景下,增加1.41℃。升温显著区域集中在长江流域上游地区,中下游地区温度升高幅度较小。总的来说,4种情景中的气候变化使流域中下游和匙吻鲟原产地气候特征相似的地区增多,进而使得Ⅲ级适生区面积增加。其中,Ⅲ级适生区在中下游流域水域总面积占比从9.08%(当前时期)分别增加至21.86%(SSP126情景)、24.75%(SSP245情景)、25.22%(SSP370情景)和24.85%(SSP585情景),这有利于匙吻鲟分布。此外,在未来时期,洞庭湖和鄱阳湖地区Ⅱ级适生区面积减少,Ⅲ级适生区面积增加而匙吻鲟对于湖库生活具有较好的适应能力[49],且湖库地区饵料相对丰富[1],所以未来时期匙吻鲟在洞庭湖和鄱阳湖的分布可能较广。
对于匙吻鲟来说,其在原产地主要受到的胁迫为水体连通性降低、环境污染和过度捕捞[4,61]。当前和未来时期,匙吻鲟在长江流域的适生区主要位于中下游,长江干流自葛洲坝以后江段再无水坝,且长江干流和洞庭湖以及鄱阳湖保持着正常连通状态,水体连通性对匙吻鲟迁移的阻碍较小,为其寻找育肥场和产卵场提供了必要条件。此外,在长江大保护和长江禁渔政策实施的背景下,长江流域的水质和水生生物生存状况已经出现了改善迹象[62],这些条件将为匙吻鲟生存提供有利环境。
4 结论
本文通过使用水生物种入侵性筛查工具和最大熵建模,对于外来鲟的入侵风险进行评估,对外来鲟当前和未来时期的适生区进行量化,获得以下结论:
1)本文使用了水生物种入侵性筛查工具对于10种主要养殖外来鲟的入侵风险进行了评估,评价效果良好。研究表明,10种外来鲟中,匙吻鲟在长江流域存在着最高的入侵风险,这主要因为匙吻鲟原分布区和长江流域气候相似度高、具有在其他区域的入侵历史以及在应对气候变化时比其他外来鲟更有优势。
2)本文使用最大熵建模,依据分布位点、原分布区环境变量地理数据集与长江流域环境变量地理数据集,对匙吻鲟在长江流域的当前时期适生区进行预测,发现匙吻鲟在长江流域中下游地区存在着一定面积的适生区,主要包括湖南、湖北、安徽和江西等省的部分地区。生物气候变量中的最热季降水量、年降水量、最热月份最高温度、最冷月份最低温度、昼夜温差的月平均值和降水季节性变异系数是影响匙吻鲟在长江流域分布的主要变量。
3)未来时期(2021-2040年),在未来气候模式的4种情景下,匙吻鲟的Ⅲ级适生区面积平均增加1.41~1.78倍。质心分析说明,4种情景下,总适生区质心将向东北方向迁移137.97~163.20 km,从江西省九江市西北部移向湖北省黄石市和黄冈市交界处。未来气候背景下,需要注意匙吻鲟在长江流域中下游地区的Ⅲ级适生区明显扩大,这将有利于其分布。
当前长江流域正在执行十年禁渔的政策,使得外来鲟被渔业捕捞所发现的概率降低。因此在未来,长江生物多样性保护不仅要关注和保护本土水生生物资源以及栖息地,还应该做好外来鱼类的监测与管理,避免中华鲟等诸多原生珍稀鱼类受到生物入侵的干扰,更好地保护长江本土水生生物。
5 附录
附表Ⅰ~Ⅲ见电子版(DOI: 10.18307/2023.0531)。
猜你喜欢
中国野生植物资源(2023年9期)2023-10-20 09:11:34
中国中医药信息杂志(2023年6期)2023-06-13 02:05:36
植物资源与环境学报(2020年2期)2020-04-20 07:18:02
疯狂英语·新悦读(2019年12期)2020-01-06 03:28:02
竹子学报(2018年2期)2018-10-25 02:02:12
少儿科学周刊·儿童版(2018年12期)2018-01-26 12:02:22
少儿科学周刊·少年版(2018年12期)2018-01-26 12:01:02
学习月刊(2016年14期)2016-07-11 01:54:38
三峡大学学报(自然科学版)(2016年6期)2016-04-16 05:02:38
作文评点报·低幼版(2014年41期)2014-11-03 23:46:53
