
卷烟主流烟气有害成分遗传分析
2023-08-18 08:31:53童治军唐石云徐永明方敦煌陈学军冯颖杰杨宗灿刘文召张婷婷杨金初肖炳光
中国烟草科学 2023年3期
童治军 唐石云 徐永明 方敦煌 陈学军 冯颖杰 杨宗灿 刘文召 张婷婷 杨金初 肖炳光
摘要:為深入解析卷烟主流烟气中有害成分的遗传机制,以烤烟 Y3和 K326为亲本构建的重组自交系群体(RIL )为材料,采用植物数量性状的主基因+多基因混合遗传模型对该群体连续3个世代(F6:7、F7:8和 F8:9)的一氧化碳(CO )、氢氰酸(HCN)、4-甲基亚硝胺基-1-3-吡啶基-1-丁酮(NNK )、氨(NH3)、苯并芘{B[a]P}、苯酚(PHE)、巴豆醛(CRO)和焦油(TAR )等8个主流烟气有害成分性状进行遗传分析。结果表明:(1)8个主流烟气有害成分性状在连续3个世代中均呈单峰或多峰的正态或偏正态分布,属典型的数量性状。(2)在连续3个世代中,8个性状的最优遗传模型均为4对部分等加性主基因模型,其中, CO 、HCN、NNK、NH3、B[a]P 和 CRO 的最优遗传模型为4MG-EEA ;PHE 和 TAR 的最优遗传模型为4MG-EEEA。(3)上述性状在连续3个世代中均受主基因遗传模型控制,主基因遗传率极高(均值为89.01%)且远大于环境(非遗传)因素影响。综上,这些性状主要由遗传基础决定的,可为选育烟草低危害良种提供理论依据。
关键词:重组自交系群体;主流烟气有害成分;主基因+多基因;遗传分析
中图分类号: S572.03 文献标识码: A 文章编号:1007-5119(2023)03-0016-07
Genetic Analysis of Harmful Components in Main Stream Cigarette Smoke
TONG Zhijun1, TANG Shiyun2, XU Yongming3, FANG Dunhuang1, CHEN Xuejun1, FENG Yingjie3, YANG Zongcan3, LIU Wenzhao3, ZHANG Tingting3, YANG Jinchu3*, XIAO Bingguang1*
(1. Yunnan Academy of Tobacco Agricultural Sciences, Key Laboratory of Tobacco Biotechnological Breeding, National TobaccoGenetic Engineering Research Center, Kunming 650021, China;2. Technology Centre, China Tobacco Yunnan Industrial Co., Ltd.,Kunming 650231, China;3. Technology Center, China Tobacco Henan Industrial Co., Ltd., Zhengzhou 450000, China)
Abstract: In order to analyze the genetic mechanisms of harmful components in mainstream cigarette smoke, the recombinant inbred lines (RIL) crossed by flue-cured tobaccos Y3 and K326 were used in the major gene plus polygene mixed genetic model method analyses for Carbon Monoxide (CO), Hydrocyanic acid (HCN), 4-(Methylnitrosamino)-1-(3-pyridyl)-1-butanone (NNK:), Ammonia (NH3), Benzo[a]pyrene (B[a]P), Phenol (PHE), Crotonaldehyde (CRO) and Tar (TAR) in three consecutive generations (2018F6:7, 2019F7:8 and 2020F8:9). The main results were as follows:(1) The 8 traits value of RIL populations in three generations all showed normal or partial normal distribution of single or multi peaks, belonging to typical quantitative inheritance.(2) The optimal genetic models for the 8 traits were all controlled by four major genes with partially equally additive (PEA) effects inheritance in three consecutive years. The optimal genetic model of CO, HCN, NNK, NH3, B[a]P and CRO was 4MG-EEA, controlled by four major genes with partially equally additive 1(PEA1) effect mixed inheritance; The optimal genetic model of PHE and TAR was 4MG-EEEA, controlled by four major-genes with partially equally additive 2(PEA2) effect mixed inheritance.(3) These typical harmful components in mainstream cigarette smoke were only controlled by the major gene genetic model in 3 generations, and the heritability of the main genes was higher than 85%(mean 89.01%) and much greater than that of the environmental effect. In conclusion, these traits are mainly determined by genetic basis, which can provide theoretical basis for cultivating low-harm tobacco varieties.
Keywords: recombinant inbred lines (RIL); harmful components in main stream cigarette smoke; major gene plus polygene; genetic analysis
烟草(Nicotiana tabacum L.)是茄科烟草属一年生经济作物,因其能產生一种特有植物碱——烟碱,且该物质能使烟民吸食时获得极大的满足感和愉悦感而被广泛种植。然而,烟草又是一种备受争议的经济作物。故此培育低危害优质烟草品种成为烟草新品种选育的重要方向[1-2]。卷烟主流烟气中的有害成分是判断低危害烟草品种选育成败的核心依据[3]。研究表明,卷烟烟气是一种极其复杂的混合物,是在卷烟抽吸过程中由烟草燃烧、裂解和蒸馏而产生的[3-8]。谢剑平等[3]的研究确定了对卷烟主流烟气危害性影响最大的7项有害成分指标为一氧化碳(CO)、氢氰酸(HCN)、4-甲基亚硝胺基-1-3-吡啶基-1-丁酮(NNK)、氨(NH3)、苯并芘(B[a]P)、苯酚(PHE)、巴豆醛(CRO)。此外,鉴于卷烟主流烟气中焦油(TAR)对人体健康的极大危害性,行业内将焦油与上述7种有害成分统筹考虑,科学评价卷烟主流烟气的危害性[6-8]。
近年来,数量性状主基因+多基因遗传分离分析体系[9-14]在许多植物育种研究中得到了广泛的应用,并初步获得各类植物育种性状的遗传组成,为作物和园艺蔬菜等植物提供了有意义的遗传信息。但迄今,尚未见到针对烟草主流烟气有害成分性状的遗传特性研究。本研究以2个烤烟品种 Y3和 K326为亲本构建含有271个重组自交系(RIL)的连续3个世代群体(2018F6:7、2019F7:8和2020F8:9)为研究材料,首次采用主基因+多基因混合遗传模型研究烟草主流烟气中7种代表性有害成分和焦油释放量性状,以期揭示烟草主流烟气有害成分性状的遗传机制,评估主基因的遗传效应及其遗传率,为烟草主流烟气有害成分释放量性状的遗传分析, QTL 定位及低危害优质新品种选育奠定基础。
1 材料与方法
1.1 试验材料
以Y3为母本、K326为父本配制杂交组合,2012年在云南省烟草农业科学研究院试验基地的温室种植亲本,杂交获得 F1代。同年,种植 F1并采用单粒传法,连续套袋自交,分别于2018年、2019年和2020年构建获得 F6:7、F7:8和 F8:9重组自交系(RIL),且每代 RIL 群体均含271个株系。
1.2 田间试验
连续3个世代(2018年、2019年和2020年),将双亲和RIL 群体同期播种在云南省烟草农业科学研究院试验基地,采用完全随机区组设计,设置3个重复,每个株系(包括双亲材料)种植一个小区,每个小区种植45个单株,行距1.2 m,株距0.60 m。田间管理按照优质烤烟生产技术措施进行。
1.3 卷烟主流烟气有害成分调查
对株系内各单株挂牌、采烤,烘烤后每个株系和双亲分别随机选30个和60个单株的中部叶片(C3F)混合切丝,并进行烟支卷制及主流烟气有害成分释放量数据的测量。具体方法如下:
(1)卷烟制作:将烤后烟叶样品用切丝机切成宽度1.0 mm,长度15~20 mm 的烟丝,然后用卷烟机卷制成84 mm 长的正常卷烟。每个株系卷制20支烟支,且烟支质量控制在(0.90±0.015)g。
(2)吸烟试验:按照 GB/T 19609—2004规定,将卷烟置于温度(22±2)℃、相对湿度(60±5)%的恒温恒湿箱中平衡48 h,进行质量和吸阻选择;在 SM450型直线型吸烟机上抽吸,抽吸时间2 s ,抽吸间隔58 s,抽吸容量35 mL,用剑桥滤片捕集烟气总粒相物。每份烟样(株系)每次抽吸6支烟支,重复3次。
(3)卷烟主流烟气有害成分释放量测定:将主流烟气的总粒相物溶解于萃取剂中,对各项烟气指标进行测定:NH3,离子色谱法(YC/T 377—2010《卷烟主流烟气中氨的测定》);HCN ,连续流动法(YC/T 253—2008《卷烟主流烟气中氰化氢的测定》);苯酚,高效液相色谱法(YC/T 255—2008《卷烟主流烟气中主要酚类化合物的测定》);巴豆醛,高效液相色谱法(YC/T 254—2008《卷烟主流烟气中主要羰基化合物的测定》);B[a]P,气相色谱-质谱联用法(GB/T 21130—2007《卷烟烟气总粒相物中苯并[“]芘的测定》);NNK,气相色谱-热能分析联用法(GB/T 23228—2008《卷烟主流烟气总粒相物中烟草特有 N-亚硝胺的测定》);CO ,非散射红外法(GB/T 23356—2009《卷烟烟气气相中一氧化碳的测定》);焦油,差重法(GB/T 19609—2004《卷烟用常规分析用吸烟机测定总粒相物和焦油》)。
1.4 数据统计分析
利用数量性状主基因+多基因混合遗传分析 R 软件包 SEA v2.0 SEA(https://cran.r-project.org/web/ packages/SEA/)分析 RIL 群体各株系连续3个世代的主流烟气代表性有害成分和焦油释放量表型值,按照王靖天等[11]的报道进行相关参数设置。
2 结果
2.1 主流烟气有害成分表型数据分析
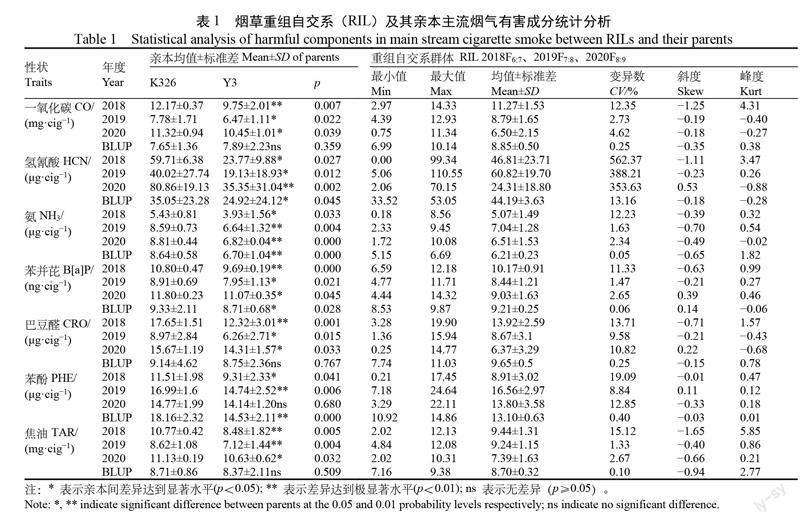
由表1可知,在3个世代和 BLUP(Best Linear Unbiased Prediction ,最佳线性无偏预测)中,除 NNK 外,剩余7个性状在双亲间呈现显著或极显著差异。由斜度和峰度值可知,8个性状分布均具有明显的多峰分布特点,表明其存在效应值较大的主基因。其中,2020年的 HCN 呈现出明显的左偏离正态分布;2018年 CO 和 TAR 则呈现出显著的右偏离正态分布趋势;2018年的 CO 、HCN 、TAR ,2019年的 NH3、CRO,2020年的 HCN、NH3、TAR 性状分布具有明显连续多峰分布特点。上述结果表明,该 RIL 群体的8个主流烟气有害成分性状在3个世代中呈连续性分布,具有显著的数量性状遗传特性。
2.2 遗传模型的选择
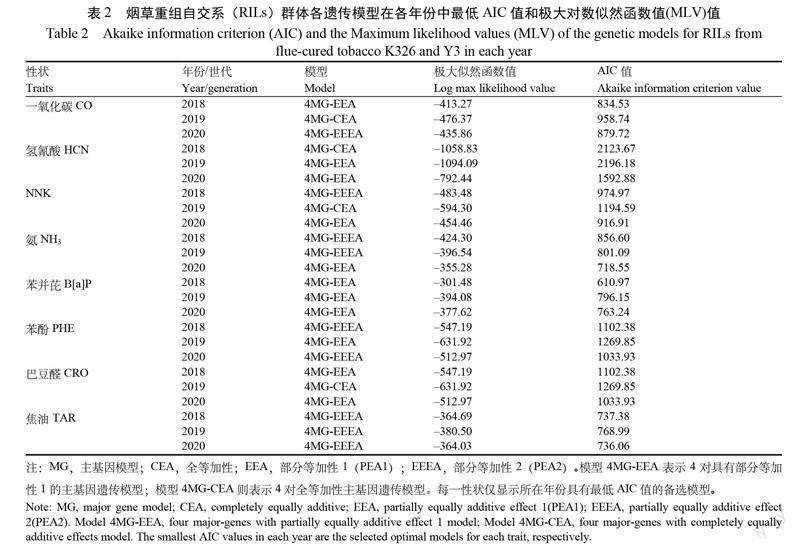
根据 AIC 值最小原则进行模型初选,选取各性状每年中 AIC 值最小的遗传模型作为该性状的候选模型(表2)。结果表明,主流烟气中 CO 的4MG-EEA ,4MG-CEA 和4MG-EEEA 模型具有最低 AIC 值,2018、2019和2020年的 AIC 值分别为834.53、958.74和879.72,可作为候选模型。同理, HCN 的3个候选模型为4MG-CEA 、4MG-EEA 和4MG-EEA ;NNK 的候选模型为4MG-EEEA 、4MG-CEA 和4MG-EEA ;NH3 的候选模型为4MG-EEEA,4MG-EEEA 和4MG-EEA;B[a]P 的3个候选模型均为4MG-EEA ;PHE 的4MG-EEEA、4MG-EEA 和4MG-EEEA 模型为候选模型; CRO的4MG-EEA、4MG-CEA 和4MG-EEA 模型为候选模型; TAR 的3个候选模型也均为4MG-EEEA。
2.3 候选模型的适合性检测与最优遗传模型估算
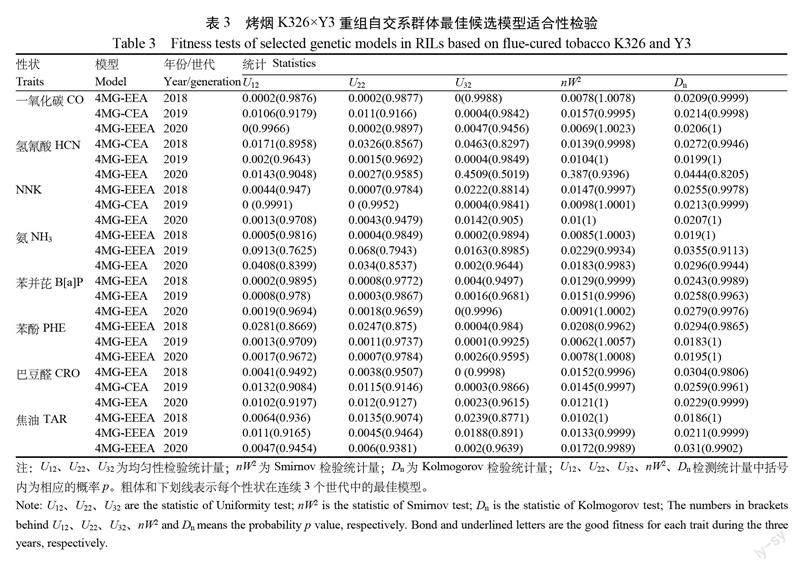
表3显示了各性状在每个世代的最适遗传模型,同时对每个性状在3个世代中的候选模型进行适合性检验(包括均匀性检验 U12、U22、U32;Smirnov检验 nW2和 Kolmogorov 检验Dn),按照王靖天等[11]方法选择各性状在连续3个世代中的最优模型。结果表明,CO 三个候选模型在连续3个世代共15个统计量中达到显著水平(p<0.05)的个数均是0,因此, CO 的最优遗传模型为4MG-EEA,受4对具有部分等加性1的主基因混合遗传模型控制。与之类似, HCN 、NNK、NH3、B[a]P 和 CRO 的最优遗传模型也为4MG-EEA;PHE 和 TAR 的最优遗传模型则为4MG-EEEA,即,受4对具有部分等加性2的主基因混合遗传模型控制。
2.4 遗传参数估计
由8个主流烟气有害成分最优遗传模型的一、二阶遗传参数(表4)可知:控制 CO 的第1对和第2对主基因的加性效应值(da 和db)分别是0.76 和0.47,表现出较高的正向遗传效应。其主基因的遗传率高达97.16%,表明控制 CO 的4个主基因具有明显的累加正效应,结果表现出主效多基因遗传,且前2个主基因起主导作用。此外,因主基因决定了该表型变异的97.16%,环境因素仅占表型变异的2.84%,表明 CO 性状主要受遗传因素控制,非遗传(环境)因素对该性状的影响可忽略不计。
控制 HCN 、NNK、NH3、B[a]P 、CRO 与 CO 的最佳遗传模型相同(4MG-EEA),其各自的第1、2对主基因加性效应值均为正,表现出较高的正向遗传效应;除 HCN 外,其余4个性状的第1对主基因加性效应远大于第2对主基因加性效应(约为2倍),表明上述性状的主基因加性效应中以第1对主基因为主。同样,因仅存主基因,且各性状的主基因遗传率分别为98.04%、74.00%、99.01%、80.46%和79.52%,非遗传(环境)因素约占1.96%、26.00%、0.99%、19.54%和20.48%,表明控制 HCN、NNK、 NH3、B[a]P 和 CRO 性状的主基因遗传因素占主导地位。
控制主流烟气中 PHE 和 TAR 性状的最佳遗传模型相同(4MG-EEEA),均符合4对具有部分等加性2的主基因混合遗传模型。其中,控制该2个性状的第1、2对主基因加性效应值也均为正值,且起增效遗传作用;此外,控制上述2个性状的第2对主基因的加性效应值(db)远大于第1对主基因加性效应值(da),说明主基因的加性效应中以第2对主基因为主。PHE 与 TAR 的主基因遗传率分别为87.13%和96.77%,表明控制 PHE 和 TAR 性状的4对主基因在其遗传变异种起绝对支配作用,而非遗传因素(环境条件)对其遗传变异的影响相对较小。
3 讨论
作物品质性状大部分属于受微效多基因控制和环境因素影响的复杂数量性状。前人对作物复杂数量性状的遗传研究,主要采用传统统计方法获得加性方差、显性方差、表型方差及环境方差等参数,进而估算和分析其性状的整体基因效应[9]。因上述传统分析方法获得的信息量有限,且分析结果在育种实践中存在较大的局限性,盖钧镒等[9-10]提出并建立了主基因+多基因混合遗传模型是植物数量性状遗传研究的重要方法,已广泛用于植物数量性状遗传分析中并获得了较理想的结果。该方法在烟草中也有报道,但主要集中在烟草农艺性状[13,15-19]、烘烤特性[20-21]、抗病性[13,22-24]、部分烟叶化学成分[12,14,25]及杂种优势[17,26]中,迄今尚未有针对烟草主流烟气有害成分性状的遗传分析研究。本研究的8个主流烟气有害成分释放量性状在连续3个世代的 RIL 群体中呈现出连续性且符合正态或偏正态分布,属典型的数量性状特征,表明了利用主基因+多基因混合遗传模型对烟草主流烟气代表性有害成分性状遗传分析的可行性和有效性。对其遗传规律的分析表明,8个主流烟气有害成分释放量性状在连续3年中的遗传变异均受4对具有部分等加性的主基因遗传模型控制,其中, CO 、HCN 、NNK 、NH3、 B[a]P 和 CRO 均受4对具有部分等加性1的主基因遗传模型(4MG-EEA)控制; PHE 和 TAR 则均符合4 对具有部分等加性2 的主基因遗传模型(4MG-EEEA)。
对烟草主流烟气中有害成分释放量性状遗传率的分析发现,8个性状的主基因遗传率范围为74.00%~99.01%,其中, CO 、HCN 、NH3和 TAR 等4個性状的主基因遗传率均高达96.00%以上,由此可知该4个性状的遗传完全受主基因控制,为后期进一步针对上述性状开展 QTL 定位、分子标记辅助选择改良低危害烟草品种(系)提供了科学依据。此外,8个性状的最佳遗传模型在连续3个世代中均无微效多基因存在,进一步表明该8个性状的主基因起主导作用,非遗传(环境)因素的影响较小,因而可在育种早期世代实现目标性状的分离和选择,科学、高效地加快具有较低有害成分释放量的单株/系的培育进程。
4 结论
结果表明,烟草重组自交系(RIL)群体8个主流烟气有害成分性状在连续3个世代均存在较广泛变异,且呈正态或近似正态分布,属典型的数量性状。采用主基因+多基因 SEA-DH(DH or RIL )联合分析方法,可知8个性状均受4对主基因遗传模型控制,其中, CO 、HCN 、NNK、NH3、B[a]P和 CRO 等6个性状受4对具有部分等加性1的主基因遗传模型控制(4MG-EEA);PHE 和 TAR 则受4对具有部分等加性2的主基因遗传模型控制(4MG-EEEA)。8个烟草主流烟气有害成分性状的遗传变异主要由遗传因素决定(主基因遗传率平均为89.01%),其中, CO 、HCN 、NH3和 TAR 的主基因遗传率高达97.16%、98.04%、99.01%和96.77%,这些具有极高遗传率的主基因/QTL 存在将有助于进一步开展烟草低有害成分性状的分子标记辅助选择和 QTL 定位研究。
参考文献
[1] 王元英,周健. 中美主要烟草品种亲源分析与烟草育种[J].中国烟草学报,1995,3(2):11-22.
WANG Y Y, ZHOU J. Parentage analysis of major tobacco varieties and tobacco breeding in America and China[J]. Acta TabacariaSinica, 1995, 3(2):11-22.
[2] 常爱霞,贾兴华,冯全福,等.我国主要烤烟品种的亲源系谱分析及育种工作建议[J].中国烟草科学,2013,34(1):1-6.
CHANG A X, JIA X H, FENG Q F, et al. Parentage analysis of Chinese Flue-cured tobacco varieties and breeding suggestion[J]. Chinese Tobacco Science, 2013, 34(1):1-6.
[3] 谢剑平,刘惠民,朱茂祥,等.卷烟烟气危害性指数研究[J].烟草科技,2009(2):5-15.
XIE J P, LIU H M, ZHU M X, et al. Development of a novel hazard index of main stream cigarette smoke and its application on risk evaluation of cigarette products[J]. Tobacco Science & Technology, 2009(2):5-15.
[4] 王珂清,秦艳华,吴洋,等.加热卷烟烟气中氨释放特性研究[J].中国烟草科学,2021,42(6):74-78.
WANG K Q, QIN Y H, WU Y, et. Study on the characteristics of ammonia release in heat-not-burn cigarette smoke[J]. Chinese Tobacco Science, 2021, 42(6):74-78.
[5] 王林,周平,贺佩,等.糖类物资对烟草香气品质的影响研究进展[J].中国烟草科学,2021,42(6):92-98.
WANG L, ZHOU P, HE P, et al. Research progress on the influence of carbohydrates on tobacco aroma quality[J]. Chinese Tobacco Science, 2021, 42(6):92-98.
[6] 赵云川,廖晓祥,陈冉,等.微波膨胀梗丝对卷烟7种烟气有害成分释放量及危害性指数的影响[J].烟草科技,2015,48(11):53-58.
ZHAO Y C, LIAO X X, CHEN R, et al. Influences of microwave expanded cut stems on deliveries of seven harmful components in mainstream smoke and hazard index of cigarette[J]. Tobacco Science & Technology, 2015, 48(11):53-58.
[7] 闫宁,刘加红,杜咏梅,等.我国不同产区烤烟烟叶主流烟气主要有害成分分析[J].中国烟草科学,2017,38(1):85-90.
YAN N, LIU J H, DU Y M, et al. Analysis on main harmful components in mainstream smoke of flue-cured tobacco leaves form different areas in China[J]. Chinese Tobacco Science, 2017, 38(1):85-90.
[8] 夏国聪,马丽娜,黄红仪,等. 国内外不同品牌卷烟样品危害性指数比较[J].烟草科技,2012(6):37-40.
XIA G C, MA L N, HUANG H Y, et al. Comparison of hazard indexes of some domestic and imported cigarettes[J]. Tobacco Science & Technology, 2012(6):37-40.
[9] 蓋钧镒,章元明,王建康.植物数量性状遗传体系[M].北京:科学出版社,2003:96-102.
GAI J Y, ZHANG Y M, WANG J K. Genetic system of quantitative traits in plants[M]. Beijing: Science Press, 2003:96-102.
[10] 章元明,盖钧镒,王永军.利用 P1、P2和 DH 或 RIL 群体联合分离分析的拓展[J].遗传,2001(5):467-470.
ZHANG Y M, GAI J Y, WANG Y J. An expansion of joint segregation analysis of quantitative trait for using P1, P2 and DH or RIL populations[J]. Hereditas, 2001(5):467-470.
[11] 王靖天,张亚雯,杜应雯,等.数量性状主基因+多基因混合遗传分析 R 软件包 SEA v2.0[J].作物学报,2022,48(6):1416-1424.
WANG J T, ZHANG Y W, DU Y W, et al. SEA v2.0: an R software package for mixed major genes plus polygenes inheritance analysis of quantitative traits[J]. Acta AgronomicaSinica, 2022, 48(6):1416-1424.
[12] 鞠馥竹,張洪博,闫宁,等.烟草西柏三烯二醇含量的遗传分析[J].中国烟草科学,2021,42(1):1-6.
JU F Z, ZHANG H B, YAN N, et al. Genetic analysis of cembratrien-diols content in tobacco[J]. Chinese Tobacco Science, 2021, 42(1):1-6.
[13] 牛文利,巫升鑫,余文,等.烟草抗青枯病突变体153-K 的抗性遗传及与农艺性状的关系[J].中国烟草科学,2021,42(2):1-7.
NIU W L, WU S X, YU W, et al. Analysis of resistance inheritance of 153-K to bacterial wilt and its correlation with agronomic characters in tobacco[J]. Chinese Tobacco Science, 2021, 4(2):1-7.
[14] 蒋勋,杨全柳,刘国祥,等.高烟碱烤烟种质资源再鉴定及遗传多样性分析[J].中国烟草科学,2021,42(4):1-8.
JIANG X, YANG Q L, LIU G X, et al. Reidentification and genetic diversity analysis of high nicotine flue-cured tobacco germplasm[J]. Chinese Tobacco Science, 2021, 42(4):1-8.
[15] 朱惠琴,张宪银,薛庆中.烟草两个 DH 群体农艺性状的遗传分析[J].浙江大学学报(农业与生命科学版),2004,30(5):477-481.
ZHU H Q, ZHANG X Y, XUE Q Z. Genetic analysis of agronomic traits of two doubled haploid populations in tobacco[J]. Journal of Zhejiang University (Agric.& Life Sci.), 2004, 30(5):477-481.
[16] 王日新,任民,张兴伟,等.普通烟草栽培种内株高性状主基因加多基因遗传分析[J].中国烟草科学,2009,30(2):15-20.
WANG R X, REN M, ZHANG X W, et al. Genetic analysis of plant height using mixed major gene plus polygenes inheritance model in culture spawn of Nicotiana Tabacum L. [J]. Chinese Tobacco Science, 2009, 30(2):15-20.
[17] 王毅,程君奇,蔡长春,等. 白肋烟主要农艺性状的杂种优势及其遗传分析[J].中国烟草科学,2009,30(3):28-32.
WANG Y, CHENG J Q, CAI C C, et al. The heterosis and genetic analysis of main agronomic traits of burley tobacco[J]. Chinese Tobacco Science, 2009, 30(2):15-20.
[18] 张兴伟,王志德,任民,等.烤烟几个重要植物学性状的遗传分析[J].中国烟草科学,2012,33(5):1-8.
ZHANG X W, WANG Z D, REN M, et al. Genetic analysis of several important botanic traits in flue-cured tobacco[J]. Chinese Tobacco Science, 2012, 33(5):1-8.
[19] 李海洋,李荣华,赖瑞强,等.大叶密合×长脖黄重组自交系群体主要农艺性状遗传分析[J].中国烟草科学,2017,38(5):39-44.
LI H Y, LI R H, LAI R Q, et al. Genetic analysis of important agronomic traits in recombinant inbred lines of ‘DayemiheבChangbohuang[J]. Chinese Tobacco Science, 2017, 38(5):39-44.
[20] 倪超,徐秀红,张兴伟,等.烤烟品种易烤性相关性状的主基因+多基因遗传分析[J].中国烟草科学,2011,32(1):1-4.
NI C, XU X H, ZHANG X W, et al. Genetic analysis of easy curing potential in flue-cured tobacco with the mixed major-gene plus polygene inheritance model[J]. Chinese Tobacco Science, 2011, 32(1):1-4.
[21] 郝贤伟,徐秀红,许家来,等.烤烟耐烤性的遗传效应[J]. 中国农业科學,2012,45(23):4939-4946.
HAO X W, XU X H, XU J L, et al. Genetic effects of holding curing potential in flue-cured tobacco[J]. Scientia Agricultura Sinica, 2012, 45(23):4939-4946.
[22] 高加明,王志德,张兴伟,等.香料烟青枯病抗性基因的遗传分析[J].中国烟草科学,2010,31(1):1-4.
GAO J M, WANG Z D, ZHANG X W, et al. Genetic analysis on resistance to bacterial wilt in oriental tobacco[J]. Chinese Tobacco Sciences, 2010, 31(1):1-4.
[23] 张振臣,袁清华,马柱文,等.烟草品种 GDSY-1 的青枯病抗性与遗传分析[J].中国烟草科学,2017,38(4):9-16.
ZHANG Z C, YUAN Q H, MA Z W, et al. Inheritance of resistance to bacterial wilt in Chinese domestic tobacco cultivar GDSY-1[J]. Chinese Tobacco Science, 2017, 38(4):9-16.
[24] 郭璇,闫杏杏,蒋彩虹,等.雪茄烟 Beinhart1000-1对黑胫病0号生理小种的抗性遗传分析[J].中国烟草科学,2017,38(2):56-62.
GUO X, YAN X X, JIANG C H, et al. Genetic analysis of Beinhart1000-1 resistance to black shank in tobacco[J]. Chinese Tobacco Science, 2017, 38(2):56-62.
[25] 蔡长春,张俊杰,黄文昌,等.利用 DH 群体分析白肋烟烟碱含量的遗传规律[J].中国烟草学报,2009,15(4):55-60.
CAI C C, ZHANG J J, HUANG W C, et al. Genetic analysis of nicotine content in DH burley tobacco population[J]. Acta TabacariaSinica, 2009, 15(4):55-60.
[26] 巫升鑫,潘建菁,陈顺辉,等.烤烟若干农艺性状的杂种优势及其遗传分析[J].中国烟草学报,2001,7(4):17-22.
WU S X, PAN J Q, CHEN S H, et al. The analysis of heterosis and inheritance of some agronomic characters of flue-cured tobacco[J]. Acta TabacariaSinica, 2001, 7(4):17-22.
