
淬灭酶对亚硝化-混合自养脱氮系统的影响
2023-08-16 09:07陈娜张肖静张楠马冰冰张涵杨浩洁张宏忠
化工进展 2023年7期
陈娜,张肖静,张楠,马冰冰,张涵,杨浩洁,张宏忠
(郑州轻工业大学材料与化学工程学院,环境污染治理与生态修复河南省协同创新中心,河南 郑州 450001)
目前脱氮仍是污水处理领域的研究重点,特别是低碳氮比(C/N)条件下的高效脱氮问题仍亟待解决。传统的硝化反硝化工艺因受碳源不足的影响,脱氮效率严重下降[1]。自养脱氮技术作为一种新型脱氮技术,可在低氧环境和无需有机碳源的条件下,实现脱氮[2]。据不完全统计,我国大约有65%以上的污水处理厂存在进水碳源不足的现象,针对于此,基于厌氧氨氧化的自养脱氮技术表现出明显优势[3]。由于自养脱氮系统中缺乏有机质,且系统污泥龄较长,因此系统中在伴随大量微生物凋亡释放出原生质时,易诱导内源反硝化的产生;另一方面在厌氧或缺氧条件下,部分微生物能够吸收挥发性脂肪酸将其转化为内部储能物质,外部碳源不足时可分解胞内聚合物进行内源反硝化[4-7]。因此,本研究建立了亚硝化-混合自养脱氮工艺(厌氧氨氧化和内源反硝化),用于处理低C/N 废水。据报道,亚硝化菌、反硝化菌及厌氧氨氧化菌均具有群体感应性能[8-10],因此,该工艺极有可能受到群体感应的影响。
群体感应是微生物间的通信机制,通过释放信号分子调控微生物的基因表达、胞外聚合物(extracellular polymeric substance,EPS)的 分 泌、生物膜的形成和蛋白酶的合成等[11]。群体淬灭则是群体感应的反过程。群体感应淬灭酶可以分解微生物之间用于交流的信号分子,从而使群体感应减弱或失效。目前研究较多的淬灭酶主要是酰基高丝氨酸内酯(acyl homoserine lactone,AHLs)介导的群体感应淬灭酶,包括AHLs-内酯酶、AHLs-酰基转移酶以及AHLs-氧化还原酶。2009 年,Yeon 等[12]首次将酰基转移酶应用于污水深度处理的膜生物反应器(membrane bioreactor,MBR)中,发现AHL-酰基转移酶能使AHLs类信号分子淬灭,从而防止生物膜的产生,减缓膜污染。随后,Kim 等[13]通过压力作用直接将酰基转移酶固定在膜组件上,发现能够显著抑制生物膜生长,有效缓解了膜堵塞现象。在此背景下,相关研究也不断拓宽,如崔雪燕[9]通过研究酰基转移酶对菌株反硝化的影响,发现20mg/L 的淬灭酶对菌株生长并未造成影响,但对反硝化过程具有明显的促进作用。但这类研究大多关注微生物菌株的变化或者膜污染控制方面,未对反应系统的脱氮性能变化进行深入探究。
因此,本文以亚硝化-混合自养脱氮系统为研究对象,跟踪分析群体感应淬灭酶-AHLs 酰基转移酶对反应体系脱氮性能、EPS、溶解性微生物产物(soluble microbial products,SMP)的分泌、关键酶活和微生物群落的影响,探究淬灭酶对亚硝化-混合自养脱氮工艺的调控作用,以期为低C/N废水的高效低耗处理提供理论支持,并为生物脱氮处理技术提供新的探索途径。
1 材料与方法
1.1 实验设置
本研究接种污泥采用培养良好的MBR 反应器中的自养脱氮活性污泥。该MBR 已经在实验室运行两年以上,在进行本文实验之前,MBR 的水力停留时间为18h,除了每天取样带走的泥量,没有额外排泥,膜组件也保证了所有污泥被截留在反应器中。因此,该MBR 中存在大量死亡微生物残体释放的SMP,进而为内源反硝化提供了碳源[14]。该反应器中氨氮的氧化主要以亚硝化为主,总氮去除以亚硝化-内源反硝化为主,同时含有微量的厌氧氨氧化过程,但并未检测到厌氧氨氧化菌。接种污泥的总氮去除负荷约为0.2kg/(m3·d),混合液悬浮固体(mixed liquor suspended solids,MLSS)的质量浓度为10g/L。
将接种污泥混匀后等分至2个血清瓶中,分别命名为R1 和R2,R1 作为空白对照不加淬灭酶,R2中加入2μmol/L的淬灭酶,其他运行条件完全相同。两个反应器置于摇床,每天于150r/min条件下避光反应18h,整个实验运行30d。每个周期设置如下:进水5min,反应18h,静置0.5h,换水5min,换水比控制在70%,剩余时间静置。具体运行参数见表1。
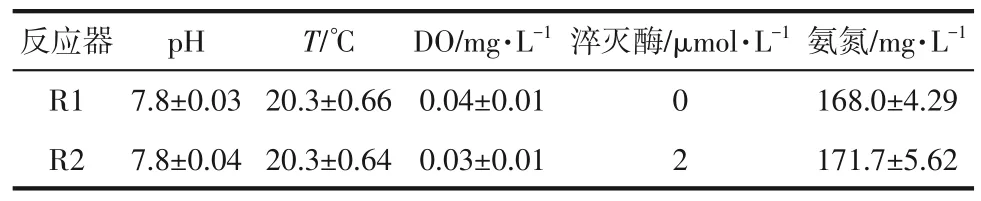
表1 各反应器运行条件和平均进水水质
淬灭酶为酰基转移酶Ⅰ(C30H34Cl2N4O),酶活力为500~1500U/mg,美国Sigma 公司产,淬灭酶在投加时先溶于甲醇中配成浓度为1g/L 的储备溶液,再把溶液加入反应器中。因此空白对照组R1中各周期也添加了相同量的甲醇,以保证实验的单一变量。实验用水采用人工配置的模拟废水,(NH4)2SO4用于提供氨氮,NaHCO3用于提供碱度。各周期始末测定pH、T和DO,并取水样测定反应前后氨氮、亚氮、硝氮的浓度。整个实验结束后,取污泥样品用于测定EPS、SMP、关键酶活力和微生物群落结构。
1.2 水质指标测定方法
两个反应器进出水氨氮、亚氮和硝氮分别采用纳氏试剂分光光度法、N-1-萘基乙二胺分光光度法和紫外分光光度法测定[15],pH、DO 和T采用便携式多参数水质分析仪(德国WTW)测定,MLSS含量采用称重法测定。总氮去除率(total nitrogen removal efficiency,TNRE)、氨氮去除率(ammonia removal efficiency,ARE)分别按照式(1)和式(2)计算。
1.3 EPS和SMP的测定
取5mL 污泥混合液,在8000r/min 下离心15min,上清液用于测定SMP。用pH 为7.0 磷酸缓冲溶液将剩余泥样稀释至原体积,超声3min 后于水浴锅中80℃条件下,将样品加热30min,每隔10min 摇匀1 次。冷却后,在8000r/min 下离心15min,所得上清液用于EPS 的测定,SMP 和EPS中蛋白质和多糖的含量分别采用福林-酚法和蒽酮法测定[16]。
1.4 HAO和Heme-c的测定
取混合均匀的泥水混合液,在4℃、4000r/min条件下离心3min,弃去上清液并用缓冲溶液清洗定容至原体积,再重复上述步骤两次。采用细胞破碎仪进行细胞破碎,随后离心30min,得到的上清液即为粗酶液。羟胺氧化还原酶(hydroxylamine oxidoreductase,HAO)活性的测定如下:先取4mL反应混合液(1mmol/L 铁氰化钾和1mmol/L 盐酸羟胺),再加入适量粗酶液并立即摇匀。随后在25°C下反应15min后,加入2mL HCl 溶液终止反应,室温放置5min 后于400nm 处测吸光度;空白组则先加入终止剂,再加入粗酶液。HAO 活性按照式(3)计算。
式中,A400为400nm处的吸光度;VL为测定液体积,mL;B为稀释倍数;ε为吸光系数,L/(mol·cm);b为比色皿宽度,cm;t为时间,min;V为粗酶液体积,mL;m为测定时的泥重,g。
Heme-c 的测定如下:向离心管中先加入1mL(体积分数为40%的吡啶和200mmol/L的氢氧化钠)混合溶液,再加入适量的粗酶液混匀。将混合液加入到比色皿中,再加入3mg的连二亚硫酸钠,此过程需在1min内于波长500~600nm范围内进行光谱扫描[17]。Heme-c含量按照式(4)进行计算。
式中,A500、A535为500nm 和535nm 处的吸光度的差值;m为测定时加入粗酶液所对应的泥重,g。
1.5 高通量测序
用Qubit2.0 DNA 检测试剂盒(Sangon 公司,中国)检测样品的DNA。采用高通量焦磷酸测序,使用V3-V4 高变区域的通用PCR 引物341F/805R (341F:CCTACGGGNGGCWGCAG;805R:GACTACHVGGG-TATCTAATCC)进行扩增,高通量焦磷酸测序使用MiSeq测序平台进行分析。所得序列与Silva数据库中的微生物进行了比较。
2 结果与讨论
2.1 淬灭酶对自养脱氮性能的影响
图1反映了两个反应器进出水中氮素含量及脱氮性能的变化情况。R1仅加入等量甲醇作为对照,R2 中加入了2μmol/L 的淬灭酶。R1、R2 反应器平均进水氨氮浓度分别为168.0mg/L、171.7mg/L。
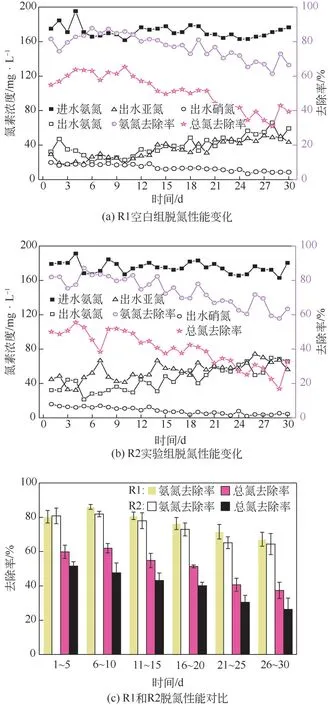
图1 淬灭酶对反应器脱氮性能的影响
在R1 中,以每5 个周期为一个阶段取平均值进行脱氮性能分析,反应器的脱氮性能呈先升高后降低的趋势。其中前两阶段出水氨氮浓度从35.3mg/L 降低至23.2mg/L,ARE 由80.4%升高至86.1%,TNRE由59.9%升高至61.9%。随后反应器各阶段出水氨氮和亚氮浓度不断升高,两者最终稳定在56.0mg/L 和47.5mg/L,出水硝氮含量不断降低,最终降至9.0mg/L;各阶段ARE 由80.3%升高至86.1%,后不断下降至67.2%,TNRE由59.9%升高至61.9%,后不断下降至37.4%。随着反应的进行,反应器中氨氮的升高和亚氮的积累可能是自养脱氮微生物无法适应新环境,导致反应器污泥活性下降。最终ARE 由80.3%下降至67.2%,TNRE 由59.9% 下降至37.4%,两者的下降幅度分别为13.1%和22.5%。根据进出水氮素浓度变化规律、较高的ARE和较低的TNRE,可以推测反应器中大部分的亚氮都由好氧氨氮氧化生成,由于两组反应器DO 均小于0.05mg/L,因此反应器中基本不存在硝化过程。由于两个反应器中都存在亚硝化过程,氨氮和亚氮的共存为厌氧氨氧化提供了反应的底物,因此这表明反应器中存在厌氧氨氧化过程,故反应器中大部分的硝氮主要来源于厌氧氨氧化过程,则反应器中的总氮主要依靠亚硝化-内源反硝化和少量厌氧氨氧化共同去除。
R2 反应器中加入2μmol/L 的淬灭酶,与R1 进行相同分析,反应器脱氮性能呈现不断下降趋势。其中前两个阶段出水氨氮由最初的34.7mg/L降低至31.5mg/L,ARE 由80.8%升高至81.9%。随后反应器各阶段出水氨氮和亚氮浓度不断升高,两者最终稳定在61.4mg/L和66.5mg/L,出水硝氮含量不断降低,最终降至4.2mg/L;各阶段脱氮性能ARE 由80.8%升高至81.9%,后不断下降至64.4%,TNRE由51.5%不断下降至26.4%,两者最终的下降幅度分别为16.4%和25.1%。
结合图1两组反应器各阶段氮素变化和脱氮性能的变化可知,R2 内出现了明显的亚氮积累,各阶段出水亚氮均高出R1 约19.0mg/L。阮心怡等[18]研究淬灭酶对菌株P.aeruginosa反硝化的影响结果表明,群体淬灭对提升菌株反硝化作用具有促进作用,这表明淬灭酶的加入具有促进R2 反硝化作用的可能。值得注意的是,R2各阶段ARE比R1略有降低,且出水硝氮略低于R1,各阶段TNRE 比R1平均低11.1%,这说明淬灭酶在强化亚硝化的同时,厌氧氨氧化过程受到抑制,不能及时消耗氨氮和亚氮,从而导致氨氮去除下降以及亚氮的积累。张肖静等[19]研究群体感应淬灭酶对MBR-CANON系统脱氮的影响,发现淬灭酶抑制了亚硝化和厌氧氨氧化。这一结果与本研究略有不同,可能是两个反应系统内微生物组成的不同以及淬灭酶浓度的较大差异造成的。另一方面,相关研究[9]表明,向原本含有外源信号分子的系统中加入20mg/L 的猪肾酰化酶后,厌氧反硝化作用被显著促进,但实验中各菌株亚氮积累量均较低。本文则主要表现为亚氮的积累,分析原因可能是:上述研究中反应系统含有充足的碳源,而本研究为自养系统,溶解淬灭酶的微量甲醇及内源碳源SMP 等并不足以使硝氮转化为氮气,而是停留在亚氮阶段造成亚氮积累,即短程反硝化得到强化。
2.2 淬灭酶对污泥性能的影响
EPS为微生物的主要代谢产物,其分泌为微生物抵御外界环境的一种保护机制,SMP则主要与微生物的凋亡有关[20]。图2 反映了反应器中EPS 和SMP 含量的变化情况。经过长期运行后,R1 的EPS 中蛋白质和多糖分别为45.2mg/g 和10.2mg/g,R2 的EPS 中蛋白质和多糖则分别为32.1mg/g 和5.2mg/g。与R1 相比,向反应器中投加2μmol/L 淬灭酶后,EPS 总含量由55.4mg/g 降低为37.2mg/g,原因是微生物之间会通过群感效应机制分泌大量的EPS或其他代谢产物,从而形成稳定的生物膜结构以帮助细菌向固体表面附着[21]。淬灭酶则会降解群体感应信号分子,抑制群感效应,从而降低微生物EPS的分泌。另一方面研究表明Metagenome是主要的EPS 产生菌和AHLs 类信号分子的产生菌[22]。值得注意的是,本研究向R2反应器中加入淬灭酶后,Metagenome菌属相对丰度由2.19%减少至1.46%,因此Metagenome菌属的减少直接降低了系统中EPS的含量;另一方面Metagenome菌属的减少限制了AHLs 类信号分子的产生,从而减弱了群体感应,最终表现为EPS的降低。
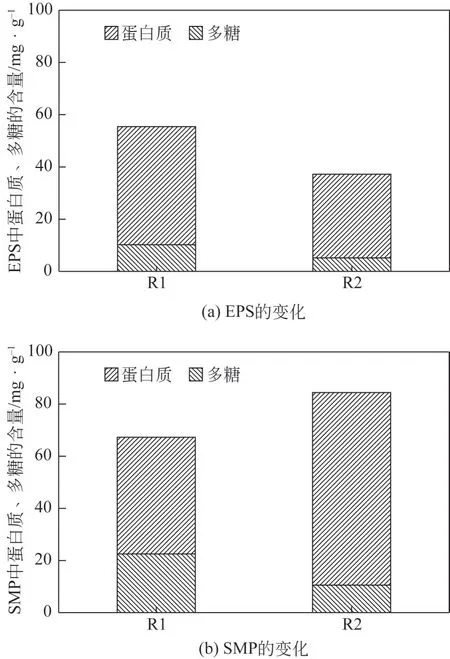
图2 长期作用后两个反应器中EPS和SMP的变化情况
R1 的SMP 中蛋白质和多糖的含量分别为44.8mg/L 和22.5mg/L,R2 的SMP 中蛋白质和多糖的含量分别为73.9mg/L和10.5mg/L,可以看出淬灭酶的加入使得反应器中SMP 的含量由67.3mg/L 升高为84.4mg/L,这表明淬灭酶的加入使得部分微生物凋亡,这与R2 中微生物丰富度和多样性降低的结果相一致。值得注意的是,淬灭酶的加入使得蛋白质含量明显升高、多糖含量明显降低,这表明淬灭酶使得SMP 的组成发生了变化。另一方面,相关研究表明[23],在厌氧和缺乏外源营养物质条件下,反硝化菌可以利用SMP作为碳源进行反硝化。可以看出,R2 反应器中EPS 和SMP 类有机质含量低于R1,可能是淬灭酶诱导了反硝化菌的增殖,从而消耗有机质促进了内源反硝化。但由于外源有机质的缺乏和反硝化菌对内源有机质利用率有限,因此致使短程内源反硝化的发生,从而造成了亚氮积累,最终反应器脱氮性能降低,该结论和2.1 节脱氮过程的分析结果一致。
2.3 淬灭酶对关键酶活的影响
HAO 为生物脱氮过程的关键酶,其在硝化、反硝化以及厌氧氨氧化过程中均发挥着重要作用[24]。一方面亚硝化菌的HAO 能将羟胺进一步氧化成亚硝酸盐,从而完成亚硝化,推动硝化作用;另一方面,异养反硝化菌的HAO 在氧化羟胺时产生一部分电子还原亚硝酸盐,从而促进反硝化过程[25]。如图3(a)所示,在R1中HAO酶活为0.80EU/g,在加入2μmol/L 淬灭酶的R2 中,HAO 酶活增高为0.99EU/g。这说明淬灭酶的加入促进了HAO 酶活,结合2.1节系统脱氮性能和脱氮途径的分析,R2出水氨氮升高、硝氮降低以及亚氮积累,都表明系统亚硝化和短程内源反硝化增强,因此HAO 的增加与上述两种反应过程的强化密切相关。
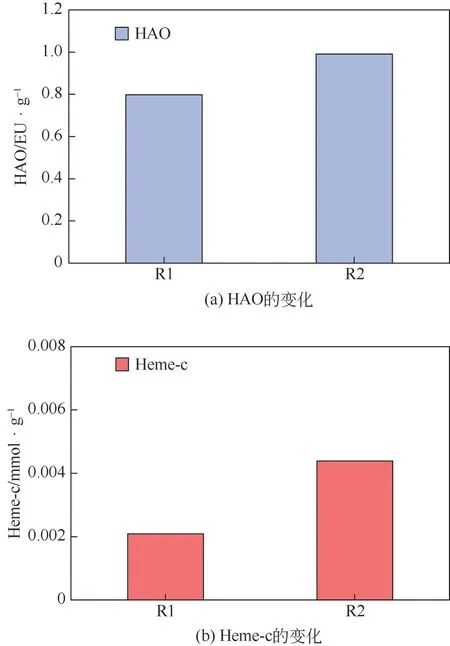
图3 长期作用后两个反应器中HAO和Heme-c的变化情况
Heme-c是厌氧氨氧化菌的特征色素[26]。图3(b)反映了反应器内Heme-c的变化情况,R1中Heme-c的含量为0.002mmol/g,R2 中Heme-c 的含量为0.004mmol/g。相较于R1,淬灭酶的加入提高了Heme-c 的含量。有研究[27]表明Heme-c 含量增加,可能是厌氧氨氧化菌受到抑制后的应激反应,通过分泌更多的Heme-c以应对外部环境冲击。结合R2中厌氧氨氧化受到抑制以及总氮去除下降的结果可知,淬灭酶的加入诱导厌氧氨氧化菌分泌更多的Heme-c 以抵抗其抑制。另一方面,据报道[28],亚氮本身对微生物有毒害作用,出水亚硝酸盐浓度为70mg/L 即可达到半抑制效果。R2 出水亚氮高达66.5mg/L,足以抑制厌氧氨氧化菌的活性,而微生物活性的降低又会反过来促进亚氮的积累。淬灭酶的抑制作用和亚氮的毒害作用相结合,是否会产生联合毒害作用,从而影响脱氮性能,相关机理还有待深入研究。
2.4 淬灭酶对微生物群落结构的影响
对各反应器的活性污泥样品进行高通量测序并分析,结果如图4和表2所示。两组反应器样品的Coverage 均等于0.998,表明数据分析覆盖了绝大多数微生物,保证了数据的可靠性。ACE 和Chao指数越大,表明以OTU 为分类单元的优势物种的丰富度越大。可以发现,空白组R1的ACE和Chao指数高出R2 近1 倍,表明淬灭酶的加入降低了优势微生物的丰度。Shannon 指数与Simpson 指数的变化趋势相反,但反映的规律一致,Shannon 指数越大代表微生物的多样性越高。R1的Shannon指数与Simpson 指数分别为3.8 和0.075,R2 分别为3.4和0.078。这表明两组反应器的微生物多样性相差不大。值得注意的是R1、R2反应器的微生物丰富度存在不容忽视的差异,造成这种差异的主要原因可能是淬灭酶的投加破环了部分微生物的群感效应,从而阻碍了部分微生物的生长繁殖。

图4 属水平微生物群落变化情况
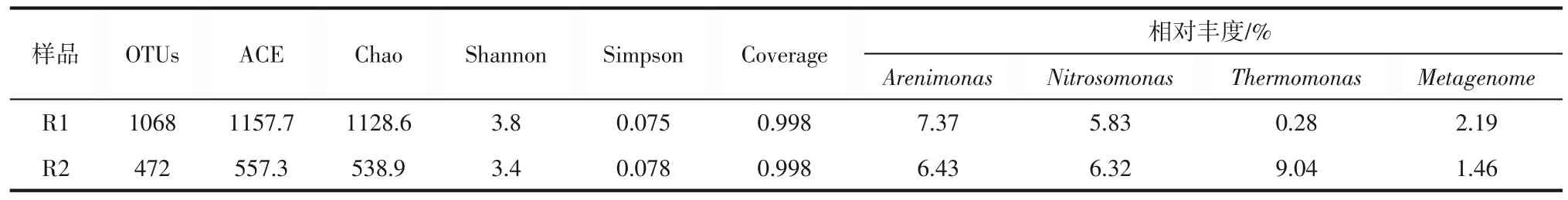
表2 微生物测序多样性指数和相对丰度
图4 反映了两组反应器中的微生物群落组成。如图所示,R1和R2菌属种类未发生变化,但各菌属相对丰度发生了变化。可以看出各反应器中均未检测出厌氧氨氧化菌,主要是因为反应器中厌氧氨氧化菌并不是优势菌属,由于浓度较低未被检出。在R1 中,优势菌属主要为Hyphomicrobium、Arenimonas、Nitrosomonas和Truepera,它们的相对丰度分别为20.56%、7.37%、5.83%和4.33%。其中Hyphomicrobium、Arenimonas和Truepera为主要的反硝化菌属[29-31],Nitrosomonas为优势亚硝化菌属[32]。在加入淬灭酶的R2 中,Nitrosomonas的相对丰度增长至6.32%。这与张肖静等[19]的AHLs 酰基转移酶使脱氮系统亚硝化菌的相对丰度减少的结果略有不同,主要可能是两个反应系统中菌群结构存在较大差异,因此亚硝化菌表现出不同的响应结果。Hyphomicrobium和Truepera反硝化菌属的相对丰度基本不变,分别为20.94% 和4.64%,Arenimonas等反硝化菌属的相对丰度略有减少,为6.43%。Zhang 等[33]研究发现,Arenimonas能有效消耗硝酸盐,从而提高总氮(TN)去除效率,故Arenimonas的减少使得TN 去除率下降。同时相关研究[34]表明,Arenimonas与厌氧氨氧化关系密切,与自养脱氮性能呈正相关,因此Arenimonas相对丰度的降低也从侧面反映厌氧氨氧化受到了抑制。有研究[35]表明,Thermomonas是短程反硝化体系中的优势反硝化菌属,值得注意的是,Thermomonas反硝化菌属的相对丰度由0.28%增长至9.04%,出现了大幅度增长,这再次证明加入淬灭酶后短程反硝化的增强。此外,有研究认为Thermomonas为亚硝酸盐产生的反硝化菌[36],这说明,淬灭酶能够促进硝氮转化成亚氮,这也解释了为什么R2出水亚氮高于R1。
结合上述分析,反应系统主要以亚硝化反应,结合内源反硝化和少量厌氧氨氧化协同脱氮,而淬灭酶的加入促进了系统短程内源反硝化并抑制了厌氧氨氧化过程,最终使R2 中亚氮积累,TNRE 下降。因此,淬灭酶有可能用于两级式自养脱氮工艺(短程反硝化-厌氧氨氧化)短程反硝化单元的强化,进而为后续厌氧氨氧化提供亚氮基质,之后与氨氮废水混合进行处理,最终实现低C/N比废水的高效处理。
3 结论
在以内源反硝化为主要脱氮过程的亚硝化-混合自养脱氮系统中存在厌氧氨氧化过程,但不占优势。2μmol/L 的淬灭酶长期作用于亚硝化-混合自养脱氮系统,导致反应器内亚氮大量积累,出水亚氮达到66.5mg/L,同时总氮去除显著下降,相比未加淬灭酶的对照组降低了11.1%。淬灭酶的加入强化了短程反硝化,但抑制了厌氧氨氧化。淬灭酶抑制了污泥中EPS的分泌,但诱导了SMP的分泌,进而为内源反硝化提供了碳源。Heme-c 的增加有可能是厌氧氨氧化菌抵抗外界不利环境的一种机制。添加2μmol/L 淬灭酶使Nitrosomonas的相对丰度由5.83%增长为6.32%,Thermomonas反硝化菌属的相对丰度由0.28%增长至9.04%。淬灭酶有望用于短程反硝化单元的强化,进而为两级式自养脱氮工艺的厌氧氨氧化单元提供亚氮基质。
猜你喜欢
保健与生活(2022年11期)2022-06-09
生物学通报(2020年10期)2020-08-13
环境保护与循环经济(2017年7期)2018-01-22
上海农业学报(2016年2期)2016-10-27
国际妇产科学杂志(2016年2期)2016-06-16
中国环境科学(2016年3期)2016-02-08
华东理工大学学报(自然科学版)(2015年4期)2015-12-01
动物营养学报(2015年10期)2015-12-01
应用海洋学学报(2014年2期)2014-11-26
云南中医学院学报(2014年3期)2014-07-31
