
授粉期高温胁迫下雄穗大小对玉米干物质积累及产量的影响
2023-08-15 11:19:22穆心愿吕姗姗卢良涛刘天学李树岩薛昌颖王宏伟赵霞夏来坤唐保军
中国农业科学 2023年15期
穆心愿,吕姗姗,卢良涛,刘天学,李树岩,薛昌颖,王宏伟,赵霞,夏来坤,唐保军
授粉期高温胁迫下雄穗大小对玉米干物质积累及产量的影响

1河南农业大学农学院,郑州 450046;2河南省农业科学院粮食作物研究所/河南省玉米绿色精准生产国际联合实验室,郑州 450002;3中国气象局/河南省农业气象保障与应用技术重点实验室,郑州 450003;4南阳市宛城区信访局,河南南阳 473000
【目的】高温是制约夏玉米高产稳产的重要气象因子之一,通过设置授粉期高温胁迫和雄穗变小处理,探明高温胁迫下雄穗大小对玉米产量形成的影响,为玉米抗逆栽培及耐高温育种提供参考。【方法】以2个玉米品种浚单20(XD20)和农华101(NH101)为试材,于2020—2021年玉米抽雄至散粉结束的10 d内采用人工模拟增温试验,同时,设置抽雄期雄穗变小处理(剪除60%雄穗分枝数),探究授粉期高温胁迫下雄穗大小对玉米干物质积累分配和产量形成的影响。【结果】2年结果表明,授粉期高温胁迫对玉米雄穗长度、分枝数、小穗数和雌雄开花动态影响较小,但导致玉米干物质积累能力及向穗部分配比例下降,影响雌穗生长发育,造成穗轴长度和穗轴粗显著降低,穗粒数显著减少,花后物质积累量及向籽粒分配比例下降,进而产量显著降低。高温胁迫后,NH101穗长的下降幅度小于XD20,但行粒数、穗粒数及花后物质积累向籽粒分配比例的下降幅度高于XD20,导致NH101产量降幅超过XD20,XD20和NH101产量降幅分别为12.32%和25.00%,可见XD20比NH101更耐高温。雄穗变小处理使XD20和NH101的雄穗分枝数和小穗数分别显著降低58.57%、42.91%和57.30%、41.34%,但对雌雄开花动态无显著影响。2个温度条件下,雄穗变小处理均能促进雌穗生长,增加穗粒数,促进花后物质向籽粒积累,进而提高产量,其中,高温条件下的XD20增产幅度最大。在常温条件下,与正常雄穗处理相比,雄穗变小处理下,XD20和NH101的产量分别平均增加2.76%和4.37%,而在高温条件下,分别增加12.47%和5.75%。【结论】授粉期高温胁迫对雄穗生长发育影响较小,但导致雌穗生长发育受到不可逆损伤,穗粒数减少,制约了花后光合同化物向籽粒分配,产量显著下降。高温条件下,适当减少雄穗分枝数可促进雌穗生长发育,增加穗粒数,促进花后物质向籽粒积累,提高产量,且大雄穗型品种浚单20增产幅度高于小雄穗型品种农华101。
玉米;授粉期高温;雄穗大小;干物质积累;籽粒产量
0 引言
【研究意义】玉米是中国三大主粮之首,在保障国家粮食安全中具有重要地位。近年来,全球气候变暖导致的极端天气愈加频繁[1],对玉米生产带来不利影响[2-3]。玉米花期包括抽雄期、吐丝期和散粉期,是玉米籽粒形成最关键的时期,亦是受高温胁迫危害最大的生长发育阶段[4]。因此,研究玉米植株响应花期高温胁迫的形态机理,为玉米抗逆栽培及耐高温育种提供实践依据和理论指导,对保障中国粮食安全具有重要的现实意义。【前人研究进展】玉米虽是喜温作物,但在一段时间内气温超过临界阈值(一般为连续3 d以上日最高气温≥35 ℃[5]),可能对植株生长发育造成不可逆的损伤[6-7]。高温胁迫对玉米生长和产量的影响也因生育阶段而异[4]。在营养生长阶段,高温胁迫会抑制植株生长和生殖器官发育,造成玉米减产[8-9];花期高温胁迫导致雌雄生殖器官发育不良,影响授粉结实,穗粒数大幅减少致使严重减产,且此阶段高温胁迫导致的玉米产量损失最大[10-11]。玉米是雌雄同株异花植物,其雌雄穗分化发育具有特殊规律,且易受环境影响[12]。研究表明,在孕穗阶段与开花散粉过程中,高温胁迫对雌雄穗发育均有影响,且雄穗受影响的程度要大于雌穗[13]。这可能与在同样高温胁迫条件下雌穗部位因叶片遮荫而温度较雄穗部位低[14],以及雌穗能够产生热激蛋白而雄穗花粉不能[15]等有关。多数研究表明,花期前后高温胁迫导致玉米雄穗分枝变小、数量减少,小花退化,总小花数和花粉量骤减,花粉活力降低[16-20];缩短雄穗散粉持续期,延缓雌穗花丝吐出,增加雌雄开花间隔期,造成花期不遇[4, 21-22];花丝绒毛数量减少,花丝活力降低[20, 22-23];这些因素均会导致雌穗授粉受精困难,降低受精率,且受害程度随温度升高和持续时间延长而加剧[16, 18]。同时,高温胁迫也影响到同化物积累与转运,即使玉米完成受精,也可能会因为物质生产不足与转运不畅而导致雌穗生长受限,败育率增加[24-25]。玉米的高温耐性存在显著的基因型差异[26]。综合前人研究结果表明,耐热性强的玉米品种往往具有较高的雄穗分枝数、花粉量、花粉活力和较短的雌雄开花间隔期[16-17, 21, 27-28]。从生产经验上来说,普遍认为雄穗分枝短、数量少且颖壳不饱满的小雄穗型玉米品种,通常花粉量较少、散粉周期短、抗高温胁迫能力较弱,而雄穗分枝数较多、颖壳饱满、花粉量多的大雄穗型品种更耐高温[28]。但玉米雄穗长在植株顶端,具有顶端优势,而且发育比雌穗早,在营养供应上明显比雌穗优越,二者间存在着竞争[29]。研究表明,雄穗分枝数与产量呈负相关[30]。在玉米群体中,雄穗的花粉供应量远超过雌穗正常授粉的实际需求量[29, 31]。因此,选育适度较小雄穗的品种已成为玉米育种的趋势。【本研究切入点】高温胁迫会造成玉米雄穗生长发育异常,导致花粉量和花粉活力大幅下降,且供试材料中雄穗分枝多的大雄穗型品种往往比雄穗分枝少的小雄穗型品种更耐高温,但雄穗大小是否与品种耐热性之间有密切关系仍不清楚。【拟解决的关键问题】本研究通过设置玉米授粉期高温胁迫和雄穗变小处理,研究高温胁迫与雄穗大小对玉米品种的雌雄穗农艺性状、开花动态、花后物质积累、产量及其构成的互作效应,探究玉米品种雄穗大小与高温耐性之间的关系,以期为未来气候变暖背景下玉米抗逆栽培及耐高温育种提供实践依据与理论参考。
1 材料与方法
1.1 试验设计
试验于2020—2021年在河南现代农业研究开发基地作物抗逆中心(河南原阳,35°01′N,113°42′E,海拔63.4 m)进行。该地区属于暖温带大陆性季风气候,试验地土壤类型为潮土,0—20 cm土层土壤有机质含量8.95 g·kg-1、全氮含量1.23 g·kg-1、碱解氮含量68.46 mg·kg-1、速效磷含量71.11 mg·kg-1、速效钾含量214.56 mg·kg-1。供试品种为农华101(NH101)和浚单20(XD20),其中,农华101的雄穗属于长轴少分枝类型,浚单20的雄穗属于短轴多分枝类型。试验采用池栽方式进行,试验池子规格为3.25 m(长)×2.40 m(宽)×2.00 m(深),每个池子作为1个小区,种植密度67 500株/hm2,每小区种植4行,行距60 cm。肥料采用玉米专用复合肥(氮磷钾养分含量比例为28﹕15﹕5),按750 kg·hm-2计算每小区施肥量,全部肥料播种时一次性作基肥施入。
试验设置品种、温度处理和雄穗处理等3个因素,采用再裂区试验设计,2个品种为主区,2个温度处理为副区,2个雄穗处理为副副区。高温处理(high temperature,HT)于玉米抽雄期开始,至散粉结束恢复大田温度,处理持续10 d,参照穆心愿等[21]方法采用方钢和PO塑料膜(厚度0.1 mm,透光率95%以上)搭建的增温棚进行高温处理;常温处理(normal temperature,NT)的小区气候条件始终与大田环境保持一致。每个温度下设置2个雄穗处理,雄穗变小处理(removal tassel branch,RB)于抽雄期(80%以上植株雄穗抽出5—10 cm)选择长势一致且具有代表性的植株进行雄穗修剪,即从雄穗主轴上均匀剪除60%的雄穗分枝数,并用碘伏给剪刀和雄穗伤口消毒,同时去除长势较弱和雄穗抽出过大的植株;正常雄穗处理(normal tassel branch,NB)是保持雄穗原样,不进行任何处理(图1)。试验中为防止小区间传粉,用100目纱网将每个小区四周围起来,高度2 m,上端高于雄穗0.5 m。每个品种下形成4个处理组合,即常温处理+正常雄穗处理(NT+NB)、常温处理+雄穗变小处理(NT+RB)、高温处理+正常雄穗处理(HT+NB)、高温处理+雄穗变小处理(HT+RB),每个处理重复3次,共24个小区。
于2020年6月6日播种,8月1日高温处理开始,8月10日高温处理结束,9月28日收获;于2021年6月11日播种,8月10日高温处理开始,8月19日高温处理结束,9月28日收获。在HT和NT处理的小区雄穗部位均放置有温度自动记录仪,记录间隔为20 min(图2)。图2显示,HT与NT处理的夜间温度无明显差异,但日间温度差异较大,且在12:00—14:00温度最高时差异最大;2020年,HT处理一天中>35 ℃和>38℃的时数约为8和5 h,NT处理分别为6和0 h;2021年,HT处理一天中>35 ℃和>38 ℃的时数约为8和4 h,NT处理分别为1和0 h;在高温处理期间,HT处理2020年的日平均气温、日最高气温和日最低气温分别较NT处理高0.1—2.8 ℃、0.1—4.9 ℃和0.2—0.7 ℃,2021年分别比NT处理高1.1—3.9 ℃、1.7—6.4 ℃和0.3—1.2 ℃。

图1 正常雄穗(NB)和雄穗变小处理(RB)示意图

图2 常温处理(NT)和高温处理(HT)条件下日平均温度和处理期间温度的变化
1.2 测定项目与方法
1.2.1 玉米雄穗农艺性状测定 于抽雄后第4天,每处理选5株长势一致且具有代表性植株的雄穗,测量雄穗主轴和分枝的长度,统计雄穗分枝数和雄穗小穗数。
1.2.2 玉米开花动态测定 在玉米12叶展期,每个处理标定长势一致且具有代表性的植株10株,于抽雄期开始,记录抽雄、吐丝、散粉开始和散粉结束时期。然后计算散粉开始日期至吐丝日期的间隔天数,即开花吐丝间隔期;散粉开始到散粉结束的持续天数,即散粉持续期。
1.2.3 玉米雌穗农艺性状测定 于高温处理结束当天,每小区选取长势一致具有代表性的植株3株,摘取雌穗带回实验室,测量雌穗长度、苞叶层数、苞叶长度、穗轴长度和穗轴粗。
1.2.4 干物质积累与分配 分别于高温处理结束当天和成熟期,每小区选长势一致具有代表性的植株3株,高温处理结束当天将植株分为茎叶和雌穗,成熟期将植株分为茎叶、穗轴和籽粒,置烘箱内105 ℃杀青30 min,然后75 ℃烘干称重,并计算各器官干物质量和分配比例。
1.2.5 产量及其构成因素 于玉米成熟期,每小区收获中间2行考种,测量穗长、穗粗、穗行数、行粒数、穗粒数和百粒重,风干后脱粒称重、测含水量,并计算小区产量。
1.3 数据处理与统计分析
采用Microsoft Excel 2019整理数据和制作表格,用SPSS 24.0软件中的多因素方差分析(Multi-way ANOVA)评价各年度的品种、温度、雄穗处理及交互作用对干物质积累和产量性状的影响,用Duncan’s test分别对2个供试品种的各测定指标进行处理间差异性检验,用Origin Pro 2022b软件作图。
2 结果
2.1 授粉期高温胁迫和雄穗变小处理对玉米雄穗农艺性状的影响
通过对处理后玉米雄穗的农艺性状进行统计分析(图3),XD20的雄穗是短轴多分枝类型,主轴长度平均为20.50 cm,分枝数平均为23.67个,而NH101属于长轴少分枝类型,主轴长度平均为27.67 cm,分枝数平均为7.50个。从2年数据分析,HT处理的4个雄穗农艺性状与NT处理无显著差异;与NB处理相比,RB处理显著降低了2个品种雄穗分枝数和小穗数。2020年,与NB处理相比,在RB处理下,XD20的雄穗分枝数和小穗数分别减少59.71%和48.29%,NH101分别减少57.81%和37.74%;2021年,与NB处理相比,在RB处理下,XD20的雄穗分枝数和小穗数分别减少57.43%和30.09%,NH101则分别减少56.82%和46.39%。
2.2 授粉期高温胁迫和雄穗变小处理对玉米开花动态的影响
通过对处理后玉米开花动态进行统计分析(图4),2个品种的开花动态无显著差异;HT处理的抽雄期、吐丝期、散粉期等与NT处理无显著差异,但HT较NT处理缩短了2个品种的散粉持续期;2个品种在RB处理下的抽雄期、吐丝期、散粉期、散粉结束期、雌雄开花间隔期和散粉持续期与NB处理无显著差异。总之,2020年XD20和NH101在HT处理下的散粉持续期较NT处理分别平均缩短了1.2和1.2 d,2021年分别平均缩短了0.3和0.3 d。
2.3 授粉期高温胁迫和雄穗变小处理对玉米雌穗农艺性状的影响
2020和2021年于高温处理结束当天,测定每个处理的雌穗长度、苞叶层数、苞叶长度、穗轴长度和穗轴粗等雌穗农艺性状(图5)。2个品种的雌穗长度、苞叶层数和苞叶长度均表现为HT与NT处理间差异不显著,穗轴长度和穗轴粗则表现为HT显著低于NT处理。2020年和2021年,与NT处理相比,HT处理下,XD20的穗轴长和穗轴粗分别降低8.96%、3.75%和9.91%、7.79%,NH101则分别降低4.70%、6.28%和4.10%、12.12%。从2年数据分析,2个温度条件下,RB处理提高了2个品种的穗轴长度和穗轴粗,但与NB处理差异不显著;与NB处理相比,在RB处理下,XD20和NH101的穗轴长和穗轴粗分别增加4.83%、2.82%和4.77%、2.05%。
2.4 授粉期高温胁迫和雄穗变小处理对玉米干物质积累与分配的影响
2.4.1 高温处理结束当天干物质积累与分配 由表1可知,HT处理导致XD20的茎叶、雌穗和单株干物质量均显著降低,而NH101的雌穗干物质量显著降低,茎叶干物质量略有增加,单株干物质量略有减少,但差异均不显著。HT处理下,在2020和2021年,XD20的茎叶、雌穗和单株干物质量分别下降6.45%、30.06%、12.47%和10.24%、29.85%、15.30%,NH101的茎叶干物质量分别增加1.06%和1.09%,雌穗和单株干物质量分别下降24.51%、5.53%和10.77%、1.89%。从2年数据综合分析,2个温度条件下,RB处理均能提高2个品种的茎叶、雌穗和单株干物质量,但与NB处理差异不显著;与NB处理相比,RB处理下,XD20的茎叶、雌穗和单株干物质量增幅在NT条件下分别为0.79%、3.62%和1.51%,而HT条件下则为5.61%、6.07%和5.78%;NH101的茎叶、雌穗和单株干物质量增幅在NT条件下分别为0.08%、0.88%和0.28%,而HT条件下则为1.35%、3.12%和1.74%。

平均值±标准误差。同一品种不同小写字母表示不同处理在P<0.05水平差异显著。下同
此外,HT处理后2个品种的干物质向雌穗分配比例有所降低,2年结果一致。2020和2021年,XD20的HT处理的干物质向雌穗分配比例分别降低20.09%和17.17%,NH101的HT处理的干物质向雌穗分配比例分别降低20.10%和9.05%。从2年数据综合分析,RB处理后2个品种的干物质向雌穗分配比例略有增加;在NT条件下,经RB处理后,XD20和NH101的干物质向雌穗分配比例分别增加2.08%和0.59%,而HT条件下则分别增加0.33%和1.36%。
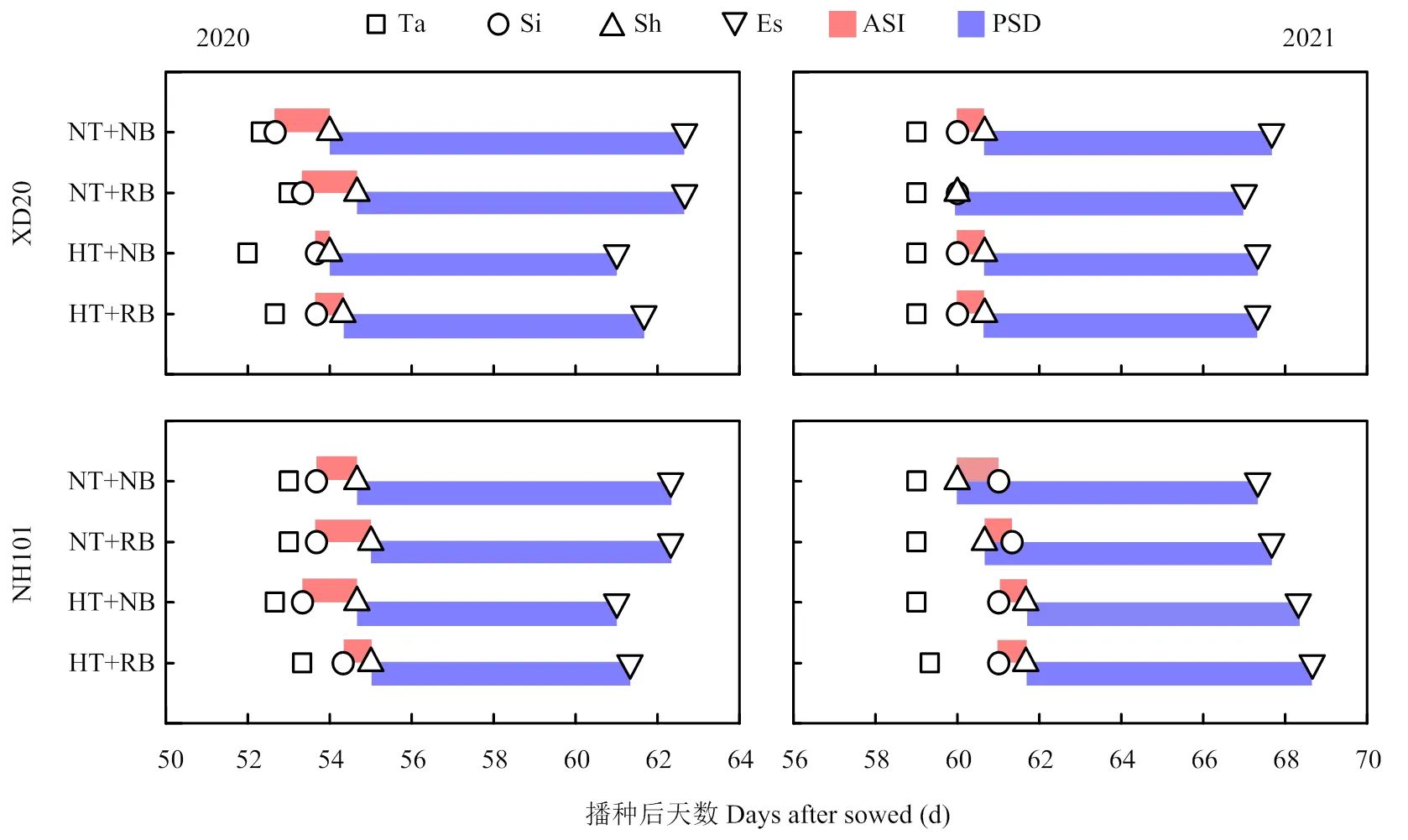
Ta:抽雄期;Si:吐丝期;Sh:散粉期;Es:散粉结束期;ASI:雌雄开花间隔期;PSD:散粉持续期
2.4.2 成熟期干物质积累与分配 由表2可知,HT处理导致2个品种成熟期穗轴、籽粒和单株干物质量显著降低,2年结果一致。2020和2021年,HT处理下,XD20的穗轴、籽粒和单株干物质量分别下降13.47%、16.74%、10.13%和24.14%、20.67%、17.58%,NH101则分别下降4.52%、26.24%、12.87%和22.74%、24.19%、14.99%。从2年数据综合分析,RB处理后籽粒干物质量显著提高,茎叶、穗轴和单株干物质量也有所增加,但与NB处理差异不显著。NT条件下,经RB处理后,XD20和NH101的籽粒和单株干物质量分别提高7.42%、6.62%和7.25%、6.32%,而HT条件下则分别增加19.49%、16.10%和5.58%、5.43%。
HT处理导致2个品种成熟期干物质向籽粒分配比例降低,NH101的降幅高于XD20,2年结果一致。2020和2021年,在HT处理下,XD20和NH101的干物质向籽粒分配比例分别降低7.60%、15.33%和3.76%、10.81%。从2年数据综合分析,RB处理后2个品种成熟期干物质向籽粒分配比例有所增加;在NT条件下,经RB处理后,XD20和NH101的干物质向籽粒分配比例分别增加0.79%和0.82%,而HT条件下则分别增加2.75%和0.11%。
2.5 授粉期高温胁迫和雄穗变小处理对玉米产量性状的影响
通过对处理后玉米产量性状的分析(表3),发现HT处理导致玉米产量显著下降,NH101的降幅高于XD20,2年结果一致。2020年和2021年,HT处理下,XD20的产量分别降低11.84%和12.80%,NH101则分别降低27.00%和20.05%。从2年数据综合分析,RB处理后玉米产量显著提高;NT条件下,经RB处理后,XD20和NH101的产量分别提高2.76%和4.37%,而HT条件下则分别提高12.47%和5.74%。
玉米产量构成因素中,HT处理后玉米穗长、穗粗、行粒数和穗粒数显著降低,但2个品种的降幅存在差异,其中,XD20穗长的降幅大于NH101,而穗粒数的降幅小于NH101。以穗粒数为例,在2020和2021年,与NT处理相比,HT处理下,XD20的穗粒数降幅分别为14.86%和15.71%,而NH101则分别为26.84%和21.19%。从2年数据综合分析,RB处理显著影响穗长、行粒数和穗粒数,均表现为RB>NB。以穗粒数为例,与NB处理相比,RB处理下,XD20和NH101在NT条件下的穗粒数增幅分别为2.67%和3.64%,而HT条件下则为6.36%和3.46%。
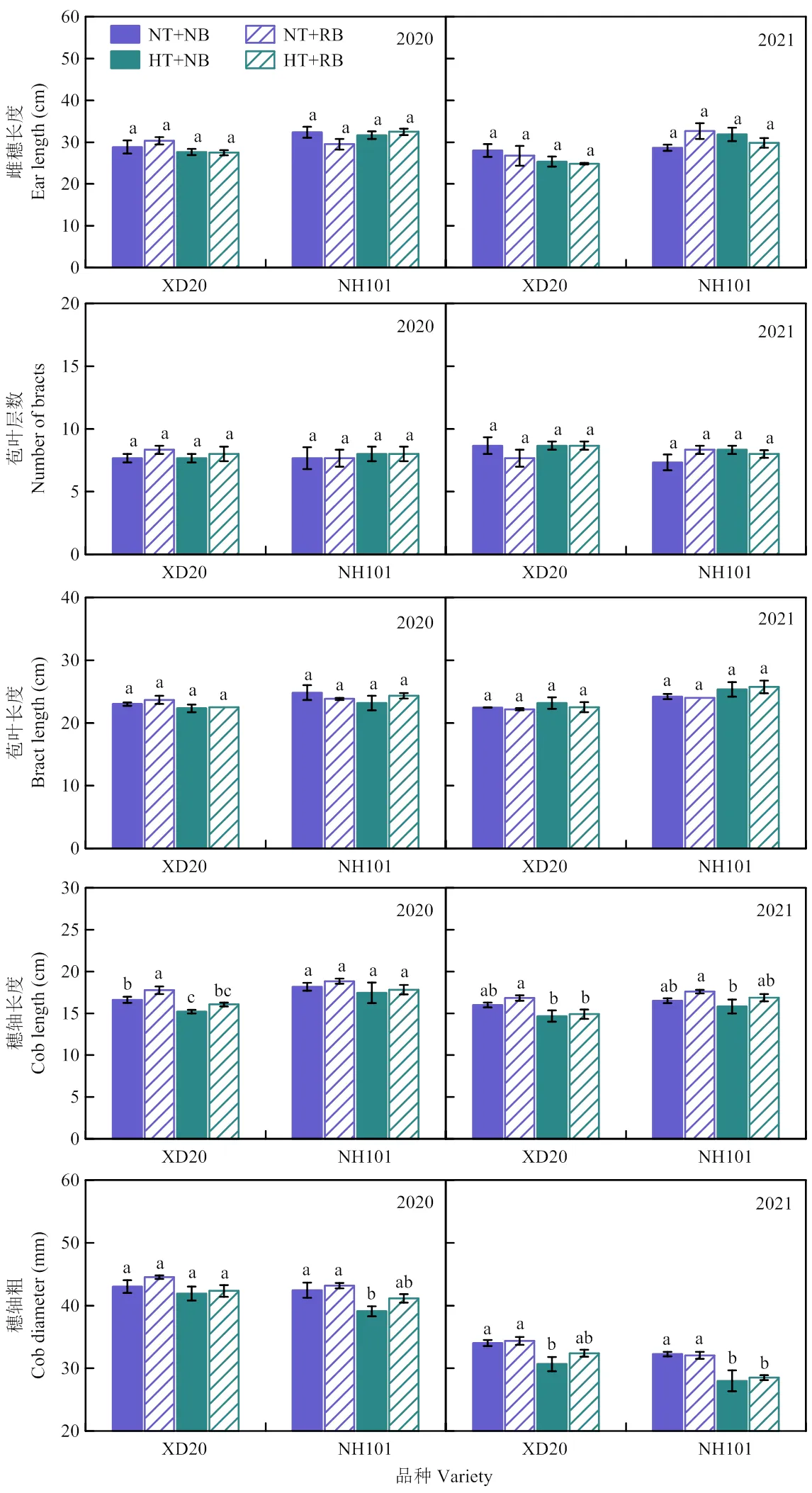
图5 授粉期高温和雄穗变小处理对玉米雌穗农艺性状的影响

表1 授粉期高温胁迫和雄穗变小处理对高温处理结束当天干物质积累与分配的影响
平均值±标准误差。同一年份同一品种不同小写字母表示在0.05水平上差异显著。*和**分别表示在0.05和0.01水平上差异显著,NS表示差异不显著。下同
Mean ± Standard error. Values within the same variety followed by different lowercase letters in the same year are significantly different at 0.05 level. * and ** indicate significant difference at 0.05 and 0.01 levels, respectively, NS indicates no significant difference. The same as below

表2 授粉期高温胁迫和雄穗变小处理对成熟期干物质积累与分配的影响
3 讨论
3.1 授粉期高温胁迫对玉米雌雄穗形态特征、干物质积累分配及产量的影响
玉米是雌雄同株异花授粉作物,雌雄穗协调发育是保证雌穗正常受精结实及产量形成的关键因素。前人研究表明,第9片叶展开期至吐丝期高温胁迫会抑制玉米雌雄穗发育,导致雄穗分枝数减少,雌穗吐丝时间推迟,开花吐丝间隔期(anthesis-silking interval,ASI)拉长[22]。抽雄后高温胁迫亦会造成玉米雄穗分枝数、主轴小花数和分枝小花密度大幅下降,散粉持续期缩短,散粉量减少[17]。花期前后持续高温胁迫对雄穗分枝数和雌雄穗总小花数影响较小[21],但会使雄穗散粉提前,散粉持续期缩短,ASI拉长[13, 21]。但也有学者研究表明,全生育期升温1.5—5 ℃对玉米吐丝和开花的持续时间或高峰日期没有显著影响,因此,对ASI亦无显著影响[32]。本研究授粉期高温胁迫对雄穗长度、分枝数和小穗数以及开花动态(如抽雄期、吐丝期、散粉期)无显著影响,但缩短了雄穗散粉持续期。这可能是因为拔节期至抽雄期是雌雄穗协调分化发育的关键时期,此阶段高温胁迫对玉米雌雄穗生长发育影响较大,而抽雄后是玉米雌雄穗爆发式生长阶段,此时的高温胁迫可能对雄穗生长及开花动态影响较小。与前人研究对比,试验的不同结果可能与高温处理时期、程度、持续时间及试验品种不同有关[4, 28]。
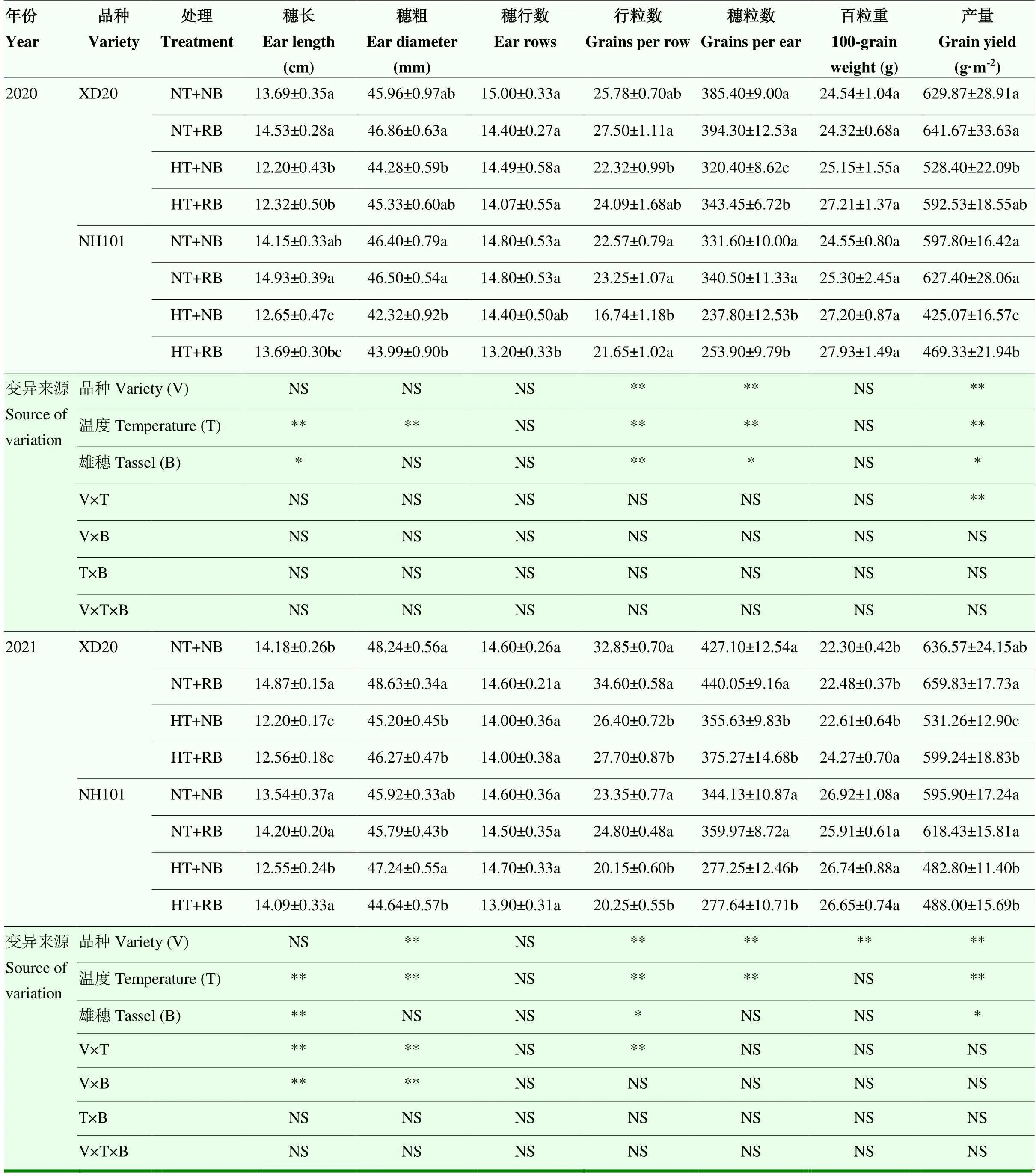
表3 授粉期高温胁迫和雄穗变小处理对玉米产量和产量构成的影响
干物质生产与分配是玉米获得产量的重要物质基础。前人研究表明,高温胁迫使玉米叶片净光合速率降低,导致同化物供应不足,且向茎叶等营养器官中的分配比例增加,向穗(籽粒)中的转运积累减少[21-22]。本研究授粉期经受高温胁迫10 d后,玉米干物质积累量降低,且向穗部分配比例减少,不能满足穗发育所需光合同化物,进而玉米穗分化受阻,穗轴长度和穗轴粗下降,导致穗粒数减少,进而籽粒库容减小,以至于成熟期干物质向籽粒的分配比例大幅下降,影响玉米产量。这与前人研究结果一致,即高温胁迫限制雌穗发育,穗粒数减少,影响了花后光合物质向籽粒的转运[4, 22]。玉米穗部性状是构成产量的重要因素,逆境胁迫影响产量构成要素的不同要件,进而影响产量。本研究表明,授粉期高温胁迫后玉米穗长、穗粗、行粒数、穗粒数和产量显著降低,而穗行数和百粒重受影响较小,因行粒数减少导致的穗粒数下降是授粉期高温胁迫造成产量大幅减少的重要原因之一。本研究与前人研究结果一致[4, 17, 24]。
玉米授粉期经受高温胁迫10 d后,虽然浚单20植株总干重和穗部干重的降幅高于农华101,且穗轴长度的下降幅度也超过农华101,但穗轴粗的降幅小于农华101;至成熟期,浚单20穗长降幅较农华101高,而行粒数、穗粒数和产量的降幅较农华101低。前期研究结果也表明,在高温胁迫条件下,浚单20会缩短果穗长度,维持较高的授粉结实率,尽可能增加穗粒数,而农华101会尽量保持果穗长度,但授粉结实率较低,极易出现秃尖、果穗花粒现象,穗粒数下降幅度较大[21]。2个供试品种雌穗发育对高温胁迫的不同响应机制可能是浚单20比农华101更耐高温的原因之一。
3.2 雄穗大小对玉米雌雄穗形态特征、干物质积累分配及产量的影响
玉米是雌雄同株异花植物,作为其生殖器官,雌雄穗在发育上存在着竞争,雄穗具有顶端优势,在营养供应上比雌穗优越[33]。研究表明,雄穗分枝数与产量呈负相关[30],而去雄可减少雄穗对雌穗营养物质的竞争,促进雌穗发育,提高玉米产量[34-36]。如在遮荫胁迫下,去雄可促进干物质向雌穗分配,提高产量[37]。本研究条件下,雄穗变小处理对玉米雄穗主轴长度和开花动态无显著影响,只是使雄穗分枝数和小穗数分别减少57.94%和42.13%。这是因为本研究只是均匀地剪除了一部分雄穗分枝,对整棵植株伤害微小,并不足以改变植株开花动态。此外,雄穗变小处理后,授粉期间雄穗的能量消耗减小,干物质向穗部分配比例增加,促进了雌穗发育,穗长、行粒数和穗粒数增加,进而产量提高。
3.3 授粉期高温胁迫下雄穗大小对玉米产量形成的影响
前人研究认为,高温胁迫会导致玉米雄穗分枝变小、数量减少,小花退化,总小花数和花粉量骤减,花粉活力降低,影响授粉结实,造成结实率下降、产量降低[13, 17-18, 24]。与之不同,本研究在大幅降低雄穗分枝数和小穗数(减少雄穗分枝数近60%和雄穗小穗数40%左右)的条件下,大雄穗型品种浚单20和小雄穗型品种农华101在授粉期高温胁迫下的穗粒数和籽粒产量表现不降反升,说明存在雄穗花粉冗余现象,大雄穗型品种浚单20比小雄穗型品种农华101更明显。这可能是因为浚单20雄穗较大,开花授粉期间雄穗对光合同化物的消耗较大,同时高温胁迫导致植株光合性能下降,光合同化物供应不足,而去除部分雄穗分枝可减少雄穗能量消耗,改善雄穗对雌穗的营养竞争,促进雌穗发育,增加穗粒数和粒重,进而提高玉米产量。相关研究也表明,在玉米群体中,雄穗的花粉供应量远超过雌穗正常授粉的实际需求量,雌雄穗同步发育比花粉量对玉米结实率的影响要大,在雌穗花丝完全吐出的情况下,一天内100粒/cm2的散粉量就能使90%以上的花丝完成授粉[29, 31]。由此可见,在不影响正常授粉的情况下,适当减少雄穗分枝数对于授粉期高温胁迫下玉米产量下降具有一定程度的缓解效应。
4 结论
授粉期高温胁迫对玉米雄穗长度、分枝数、小穗数和雌雄开花动态影响较小,主要造成植株干物质积累量降低且向穗部分配比例减少,影响雌穗生长发育,导致穗粒数减少,使成熟期干物质向籽粒的分配比例降低,产量显著下降。高温胁迫导致的穗粒数减少是产量降低的主要原因,高温条件下农华101的穗粒数和产量降幅均高于浚单20,可见农华101对高温胁迫的响应更敏感。雄穗变小处理显著降低玉米雄穗分枝数和小穗数,减少了授粉期间雄穗对养分的消耗,增加了光合同化物积累及向穗部分配,促进雌穗生长发育,增加了穗粒数,促进花后物质向籽粒积累,提高产量。高温胁迫下,雄穗变小处理对大雄穗型品种浚单20的产量增幅高于小雄穗型品种农华101。
[1] ABBASS K, QASIM M Z, SONG H M, MURSHED M, MAHMOOD H, YOUNIS I. A review of the global climate change impacts, adaptation, and sustainable mitigation measures. Environmental Science and Pollution Research, 2022, 29(28): 42539-42559.
[2] 陈怀亮, 李树岩. 气候变暖背景下河南省夏玉米花期高温灾害风险预估. 中国生态农业学报(中英文), 2020, 28(3): 337-348.
CHEN H L, LI S Y. Prediction of high temperature disaster risks during summer maize flowering under future climate warming background in Henan Province. Chinese Journal of Eco-Agriculture, 2020, 28(3): 337-348. (in Chinese)
[3] 陆伟婷, 于欢, 曹胜男, 陈长青. 近20年黄淮海地区气候变暖对夏玉米生育进程及产量的影响. 中国农业科学, 2015, 48(16): 3132-3145.
LU W T, YU H, CAO S N, CHEN C Q. Effects of climate warming on growth process and yield of summer maize in Huang-Huai-Hai plain in last 20 years. Scientia Agricultura Sinica, 2015, 48(16): 3132-3245. (in Chinese)
[4] LI T, ZHANG X P, LIU Q, LIU J, CHEN Y Q, SUI P. Yield penalty of maize (L.) under heat stress in different growth stages: A review. Journal of Integrative Agriculture, 2022, 21(9): 2465-2476.
[5] 和骅芸, 胡琦, 潘学标, 马雪晴, 胡莉婷, 王晓晨, 何奇瑾. 气候变化背景下华北平原夏玉米花期高温热害特征及适宜播期分析. 中国农业气象, 2020, 41(1): 1-15.
HE H Y, HU Q, PAN X B, MA X Q, HU L T, WANG X C, HE Q J. Characteristics of heat damage during flowering period of summer maize and suitable sowing date in North China plain under climate change. Chinese Journal of Agrometeorology, 2020, 41(1): 1-15. (in Chinese)
[6] WAHID A, GELANI S, ASHRAF M, FOOLAD M R. Heat tolerance in plants: An overview. Environmental and Experimental Botany, 2007, 61(3): 199-223.
[7] CAIRNS J E, SONDER K, ZAIDI P H, VERHULST N, MAHUKU G, BABU R, NAIR S K, DAS B, GOVAERTS B, VINAYAN M T, RASHID Z, NOOR J J, DEVI P, SAN VICENTE F, PRASANNA B M. Maize production in a changing climate. Advances in Agronomy, 2012, 114: 1-58.
[8] LIZASO J I, RUIZ-RAMOS M, RODRÍGUEZ L, GABALDON- LEAL C, OLIVEIRA J A, LORITE I J, SÁNCHEZ D, GARCÍA E, RODRÍGUEZ A. Impact of high temperatures in maize: Phenology and yield components. Field Crops Research, 2018, 216: 129-140.
[9] CICCHINO M, RATTALINO EDREIRA J I, OTEGUI M E. Heat stress during late vegetative growth of maize: Effects on phenology and assessment of optimum temperature. Crop Science, 2010, 50(4): 1431-1437.
[10] LOHANI N, SINGH M B, BHALLA P L. High temperature susceptibility of sexual reproduction in crop plants. Journal of Experimental Botany, 2020, 71(2): 555-568.
[11] ZINN K E, TUNC-OZDEMIR M, HARPER J F. Temperature stress and plant sexual reproduction: uncovering the weakest links. Journal of Experimental Botany, 2010, 61(7): 1959-1968.
[12] 冀华, 李宏, 张树伟. 玉米雌雄穗发育及其与产量的关系. 山西农业科学, 2011, 39(7): 754-755, 774.
JI H, LI H, ZHANG S W. Differentiation and growth of the male and female ears and the relationship with yield in maize. Journal of Shanxi Agricultural Sciences, 2011, 39(7): 754-755, 774. (in Chinese)
[13] WANG Y Y, TAO H B, TIAN B J, SHENG D C, XU C C, ZHOU H M, HUANG S B, WANG P. Flowering dynamics, pollen, and pistil contribution to grain yield in response to high temperature during maize flowering. Environmental and Experimental Botany, 2019, 158: 80-88.
[14] 蔡丰乐, 马昕, 王帅丽, 卢良涛, 邵瑞鑫, 李鸿萍, 赵亚丽, 穆心愿, 赵霞, 李树岩, 刘天学. 氮素对高温胁迫下玉米籽粒发育的调控效应. 华北农学报, 2022, 37(3): 119-127.
CAI F L, MA X, WANG S L, LU L T, SHAO R X, LI H P, ZHAO Y L, MU X Y, ZHAO X, LI S Y, LIU T X. Effect regulating of nitrogen on grain development of maize under heat stress. Acta Agriculturae Boreali-Sinica, 2022, 37(3): 119-127. (in Chinese)
[15] DUPUIS I, DUMAS C. Influence of temperature stress onfertilization and heat shock protein synthesis in maize (L.) reproductive tissues. Plant Physiology, 1990, 94(2): 665-670.
[16] WANG Y Y, LIU X L, HOU X F, SHENG D C, DONG X, GAO Y B, WANG P, HUANG S B. Maximum lethal temperature for flowering and seed set in maize with contrasting male and female flower sensitivities. Journal of Agronomy and Crop Science, 2021, 207(4): 679-689.
[17] 闫振华, 刘东尧, 贾绪存, 杨琴, 陈艺博, 董朋飞, 王群. 花期高温干旱对玉米雄穗发育、生理特性和产量影响. 中国农业科学, 2021, 54(17): 3592-3608.
YAN Z H, LIU D Y, JIA X C, YANG Q, CHEN Y B, DONG P F, WANG Q. Maize tassel development, physiological traits and yield under heat and drought stress during flowering stage. Scientia Agricultura Sinica, 2021, 54(17): 3592-3608. (in Chinese)
[18] WANG Y Y, SHENG D C, ZHANG P, DONG X, YAN Y, HOU X F, WANG P, HUANG S B. High temperature sensitivity of kernel formation in different short periods around silking in maize. Environmental and Experimental Botany, 2021, 183: 104343.
[19] 侯昕芳, 王媛媛, 黄收兵, 董昕, 陶洪斌, 王璞. 花期前后高温对玉米花粉发育及结实率的影响. 中国农业大学学报, 2020, 25(3): 10-16.
HOU X F, WANG Y Y, HUANG S B, DONG X, TAO H B, WANG P. Effects of high temperature during flowering on pollen development and seed setting rate of maize (L.). Journal of China Agricultural University, 2020, 25(3): 10-16. (in Chinese)
[20] 于康珂, 孙宁宁, 詹静, 顾海靖, 刘刚, 顾海靖, 潘利文, 刘天学. 高温胁迫对不同热敏型玉米品种雌雄穗生理特性的影响. 玉米科学, 2017, 25(4): 84-91.
YU K K, SUN N N, ZHAN J, LIU G, GU H J, LIU G, PAN L W, LIU T X. Effect of high temperature stress on physiological characteristics of tassel and ear in different maize varieties. Journal of Maize Sciences, 2017, 25(4): 84-91. (in Chinese)
[21] 穆心愿, 马智艳, 张兰薰, 付景, 刘天学, 丁勇, 夏来坤, 张凤启, 张君, 齐建双. 不同耐/感玉米品种的叶片光合荧光特性, 授粉结实和产量构成因素对花期高温的反应. 中国生态农业学报(中英文), 2022, 30(1): 57-71.
MU X Y, MA Z Y, ZHANG L X, FU J, LIU T X, DING Y, XIA L K, ZHANG F Q, ZHANG J, QI J S, ZHAO X, TANG B J. Responses of photosynthetic fluorescence characteristics, pollination, and yield components of maize cultivars to high temperature during flowering. Chinese Journal of Eco-Agriculture, 2022, 30(1): 57-71. (in Chinese)
[22] 高英波, 张慧, 单晶, 薛艳芳, 钱欣, 代红翠, 刘开昌, 李宗新. 吐丝前高温胁迫对不同耐热型夏玉米产量及穗发育特征的影响. 中国农业科学, 2020, 53(19): 3954-3963.
GAO Y B, ZHANG H, SHAN J, XUE Y F, QIAN X, DAI H C, LIU K C, LI Z X. Effects of pre-silking high temperature stress on yield and ear development characteristics of different heat-resistant summer maize cultivars. Scientia Agricultura Sinica, 2020, 53(19): 3954-3963. (in Chinese)
[23] 徐美玲. 温度对玉米花丝生活力的影响. 浙江农业科学, 2002, 1(3): 120-122.
XU M L. Impacts of temperature on vitality of pistils in maize. Journal of Zhejiang Agricultural Sciences, 2002, 1(3): 120-122. (in Chinese)
[24] LIU X W, WANG X L, WANG X Y, GAO J, LUO N, MENG Q F, WANG P. Dissecting the critical stage in the response of maize kernel set to individual and combined drought and heat stress around flowering. Environmental and Experimental Botany, 2020, 179: 104213.
[25] NIU S D, DU X, WEI D J, LIU S S, Tang Q, BIAN D H, ZHANG Y R, CUI Y H, GAO Z. Heat stress after pollination reduces kernel number in maize by insufficient assimilates. Frontiers in Genetics, 2021, 12: 728166.
[26] 高英波, 张慧, 王竹, 薄丽秀, 武智民, 薛艳芳, 钱欣, 代红翠, 韩小伟, 李宗新. 夏玉米品种花期耐热性鉴定与评价. 山东农业科学, 2019, 51(6): 43-48.
GAO Y B, ZHANG H, WANG Z, BO L X, WU Z M, XUE Y F, QIAN X, DAI H C, HAN X W, LI Z X. Identification and evaluation of heat tolerance of summer maize varieties during flowering stage. Shandong Agricultural Sciences, 2019, 51(6): 43-48. (in Chinese)
[27] 赵霞, 穆心愿, 马智艳, 刘天学, 齐红志, 丁勇, 张凤启, 张君, 赵发欣, 邢健伟, 吴东洪, 唐保军. 不同玉米杂交种对花期高温、干旱复合胁迫的响应. 河南农业科学, 2017, 46(8): 32-37.
ZHAO X, MU X Y, MA Z Y, LIU T X, QI H Z, DING Y, ZHANG F Q, ZHANG J, ZHAO F X, XING J W, WU D H, TANG B J. Response of different maize hybrids to high temperature and drought stresses at flowering stage. Journal of Henan Agricultural Sciences, 2017, 46(8): 32-37. (in Chinese)
[28] 任寒, 刘鹏, 董树亭, 张吉旺, 赵斌. 高温胁迫影响玉米生长发育的生理机制研究进展. 玉米科学, 2019, 27(5): 109-115.
REN H, LIU P, DONG S T, ZHANG J W, ZHAO B. Research advancements of effect of high temperature stress on growth and development of maize. Journal of Maize Sciences, 2019, 27(5): 109-115. (in Chinese)
[29] 岳玉兰, 朱敏, 于雷, 刘春光. 玉米雄穗对产量影响研究进展. 玉米科学, 2010, 18(4): 150-152.
YUE Y L, ZHU M, YU L, LIU C G. Research progress on the impact of maize tassel on yield. Journal of Maize Sciences, 2010, 18(4): 150-152. (in Chinese)
[30] GERALDI I, FILHO J B M, VENCOVSKY R. Estimates of genetic parameters for tosses characters in maize (L.) and breeding perspectives. Maydica, 1985, 30: 1-14.
[31] LIZASO J I, WESTGATE M E, BATCHELOR W D, FONSECA A. Predicting potential kernel set in maize from simple flowering characteristics. Crop Science, 2003, 43(3): 892-903.
[32] SHIM D, LEE K J, LEE B W. Response of phenology- and yield-related traits of maize to elevated temperature in a temperate region. The Crop Journal, 2017, 5(4): 305-316.
[33] 徐洪文, 宋凤斌, 童淑媛. 玉米雌雄穗发生发展及生理特性研究进展. 广东农业科学, 2012, 39(3): 22-24.
XU H W, SONG F B, TONG S Y. Research progress on development and physiological characteristics of ear and tassel in maize. Guangdong Agricultural Sciences, 2012, 39(3): 22-24. (in Chinese)
[34] 杨德光, 陈庚, 吴宝欣, 金新月, 宋翔雨, 郝天乐, 李威. 去雄携带顶叶对玉米光合特性、干物质积累及产量的影响. 西北农业学报, 2022, 31(1): 25-33.
YANG D G, CHEN G, WU B X, JIN X Y, SONG X Y, HAO T L, LI W. Effect of detasseling with top leaf on photosynthetic characteristics, dry matter accumulation and yield of maize. Acta Agriculturae Boreali-occidentalis Sinica, 2022, 31(1): 25-33. (in Chinese)
[35] 王磊, 杜雄, 崔彦宏, 党红凯, 李科江, 贺振营, 于秀艳. 冗余器官去除对高产夏玉米产量形成的影响. 华北农学报, 2015, 30(4): 132-138.
WANG L, DU X, CUI Y H, DANG H K, LI K J, HE Z Y, YU X Y. Effects of removing redundant organs on yield formation of high-yielding summer maize. Acta Agriculturae Boreali-Sinica, 2015, 30(4): 132-138. (in Chinese)
[36] 宋世宗, 李继平, 李文举, 李芳, 蒋付伟, 刘爱青. 玉米授粉后去雄与去叶对穗部性状及产量影响的研究. 湖南农业科学, 2011(9): 33-35.
SONG S Z, LI J P, LI W J, LI F, JIANG F W, LIU A Q. Impacts of emasculation and defoliation after maize pollination on tassel characteristics and yield. Hunan Agricultural Sciences, 2011(9): 33-35. (in Chinese)
[37] GAO Z, SUN L, REN J H, LIANG X G, SHEN S, LIN S, ZHAO X, CHEN X M, WU G, ZHOU S L. Detasseling increases kernel number in maize under shade stress. Agricultural and Forest Meteorology, 2020, 280: 107811.
Effects of tassel sizes on post-flowering dry matter accumulation and yield of different maize varieties under high temperature stress during pollination
MU XinYuan1,2, Lü ShanShan1, LU LiangTao1, LIU TianXue1, LI ShuYan3, XUE ChangYing3, WANG HongWei4, ZHAO Xia2, XIA LaiKun2, TANG BaoJun2
1College of Agronomy, Henan Agricultural University, Zhengzhou 450046;2Cereal Institute, Henan Academy of Agricultural Sciences/ Henan International Joint Laboratory on Maize Precision Production, Zhengzhou 450002;3China Meteorological Administration/Henan Key Laboratory of Agrometeorological Support and Applied Technique, Zhengzhou 450003;4Nanyang Wancheng District Letters and Calls Bureau, Nanyang 473000, Henan
【Objective】High temperature stress is one of the most critical meteorological disaster factors that restrict the high and stable yield of maize. This study explored the effect of tassel sizes on yield of different maize varieties under high temperature stress during pollination, so as to provide the theoretical basis and reference for stress-resistant cultivation and high temperature resistance breeding of maize. 【Method】This study was conducted by plot experiment in a greenhouse with two maize varietiesas the experimental materials, Xundan 20 (XD20) and Nonghua 101 (NH101), from 2020 to 2021.The effect of tassel sizes on dry matter accumulation, distribution and yield of maize under high temperature stress during pollination was investigated by setting the tassel branch removal treatment at tasseling stage.【Result】The results of two years showed that high temperature stress during pollination had little effect on tassel length, branch number, spikelet number and flowering dynamics of male and female. However, high temperature stress resulted in the decrease of dry matter accumulation capacity and distribution ratio to ear, which affected the growth and development of ear, resulting in the significant decrease of cob length and diameter, the significant decrease of grain number per ear, the decrease of proportion of matter accumulation to grain after anthesis, and the significant decrease of yield. Under high temperature stress, the decrease of ear length of NH101 was less than that of XD20, but the decrease of grain number per row, grain number per ear and the proportion of matter accumulation to grain after anthesis was higher than that of XD20, resulting in the decrease of yield of NH101 more than that of XD20, and the yield of XD20 and NH101 decreased by 12.32% and 25.00% respectively. XD20 is more resistant to heat than NH101. The tassel branch removaltreatment significantly reduced the number of tassel branch and spikelet of XD20 and NH101 by 58.57%, 42.91% and 57.30%, 41.34%, respectively, but had little effect on the flowering dynamics of male and female. Under the two temperature conditions, thetassel branch removal treatment promoted the growth of ear, increased the grain number per ear, increased the proportion of matter accumulation to grain after anthesis, and thus increased the yield. Among them, the yield increase of XD20 under high temperature conditions was the largest. Compared with normal tassel branch treatment, tassel branch removal treatment increased the yield of XD20 and NH101 by 2.76% and 4.37% under normal temperature conditions, while increased by 12.47% and 5.75% under high temperature conditions, respectively. 【Conclusion】High temperature stress during pollination has little effect on the growth and development of tassel, but it caused irreversible damage to the growth and development of ear, reduced the number of grains per ear, limited the distribution of photosynthate to grains after anthesis, and significantly reduced the yield. Under high temperature conditions, properly reducing the number of branches in tassel can promote the growth and development of ear, increase the number of grains per ear, promote the accumulation of matter to grains after anthesis, and increase the yield. In addition, the yield increase of the large tassel-type variety XD20 was higher than that of the small tassel-type variety NH101.
maize; high temperature during pollination; tassel size; dry matter mass; grain yield

10.3864/j.issn.0578-1752.2023.15.004
2022-12-02;
2023-03-02
中国气象局·河南省农业气象保障与应用技术重点开放实验室项目(AMF202108)、河南省农业科学院科技创新团队项目(2023TD37)、河南省农业科学院粮食作物研究所自主立项项目(LZZC202203)、河南省玉米产业技术体系建设专项(HARS-22-02-G2)
穆心愿,E-mail:muxinyuan@163.com。通信作者刘天学,E-mail:tianxueliu2005@163.com。通信作者夏来坤,E-mail:xialaikun@126.com
(责任编辑 杨鑫浩,李莉)
猜你喜欢
中国农业大学学报(2022年4期)2022-05-26 02:52:44
中国种业(2022年4期)2022-04-19 04:08:50
农业工程学报(2021年14期)2021-10-12 10:52:38
语文周报·教研版(2021年28期)2021-08-19 02:14:42
作物学报(2021年7期)2021-04-27 01:04:12
优雅(2017年10期)2017-10-16 14:57:29
数学理论与应用(2017年2期)2017-06-27 07:38:56
数学理论与应用(2017年2期)2017-06-27 07:38:54
广西植物(2017年10期)2017-05-30 08:22:32
环境与生活(2016年9期)2016-10-08 05:23:20
