
玉米抗倒伏相关性状的配合力分析
2023-08-14 05:16:26高花雨任在斌冯慧刘育生吴孝波陈芳刘勇强董绍斌廖增强黄强
江苏农业科学 2023年13期
高花雨 任在斌 冯慧 刘育生 吴孝波 陈芳 刘勇强 董绍斌 廖增强 黄强

摘要:为研究玉米抗倒伏相关性状的遗传基础,了解玉米亲本的抗倒伏相关性状的配合力和遗传率,本研究以5个抗倒伏性能差异的玉米自交系、12个常用及自选自交系按不完全双列杂交(NCⅡ)模式配置的60个组合为供试材料,对第3茎节穿刺强度、第5茎节穿刺强度、株高、穗位高、纤维素含量、半纤维素含量和木质素含量等7个性状进行配合力分析。结果表明,自交系RP125、A185、48-2和KDF的抗倒伏相关性状的一般配合力最优,可用于配置抗倒伏玉米杂交品种的优良自交系或用于遗传改良;60个杂交组合的特殊配合力相对效应值存在一定的差异,表明这些性状也受非加性效应的影响,组合A183×48-2、A185×K169和A189×KDF表现最优;所有抗倒伏相关性状的狭义遗传力均小于50%,这些性状宜于在晚代选择。
关键词:玉米;配合力;抗倒伏;穿刺强度;纤维素;半纤维素;木质素
中图分类号:S513.034 文献标志码:A
文章编号:1002-1302(2023)13-0089-06
玉米倒伏是指玉米茎秆因为各种因素从垂直于地面的生长状态变为倒折状态。玉米茎秆的倒伏不仅会影响玉米的质量,还会影响玉米的产量,导致玉米产量损失5%~25%[1]。原本的種植排列被玉米倒伏的茎秆互相遮挡而打破,使得原本均匀充足的光合效率锐减,从而影响玉米的灌浆[2-3]。茎秆互相遮挡导致田间透气性下降,影响营养输送,病虫害加剧。根据国家标准GB/T 21962—2008《玉米收获机械 技术条件》中规定机械粒收的条件为田间植株倒伏率应低于5%[4],倒伏之后的玉米影响机械化收获,导致收获效率低、产品质量差[3]。玉米茎秆的抗倒伏能力除自然环境影响外,玉米的株高和穗位高低[5-6]、茎秆的机械强度[7-9]、茎秆结构差异[10-11]都会不同程度地影响玉米茎秆的抗倒伏性能。已有的研究还将玉米茎秆穿刺强度作为评价和筛选抗倒品种的依据[12]。选取不同品种,采用不同种植密度的研究发现,倒伏率和纤维素含量与抗折力呈显著正相关,因此,纤维素含量可作为评定玉米茎秆抗倒伏能力的关键指标[13]。甚至有研究认为,茎秆中的纤维素含量对玉米茎秆穿刺强度的大小起决定作用[14]。同时,木质素代谢能力与茎秆的抗倒伏性能也呈正相关[15]。此外,田间的种植条件[16]和水肥管理[17]同样会影响玉米倒伏的发生。因此,对选育的玉米新材料抗倒伏的相关性状进行配合力分析十分有必要。本研究以5个抗倒伏性能差异的自交系、12个常用及自选自交系配制的60个杂交组合为材料,对玉米抗倒伏相关性状茎秆穿刺强度、株高、穗位高、半纤维素含量、纤维素含量、木质素含量进行测量,从不同角度了解玉米倒伏的不同性状特征,分析不同自交系和杂交组合材料的一般配合力(general combining ability,简称GCA)和特殊配合力(special combining ability,简称SCA),为进一步培育强优势抗倒玉米杂交品种提供理论依据。
1 材料与方法
1.1 供试材料
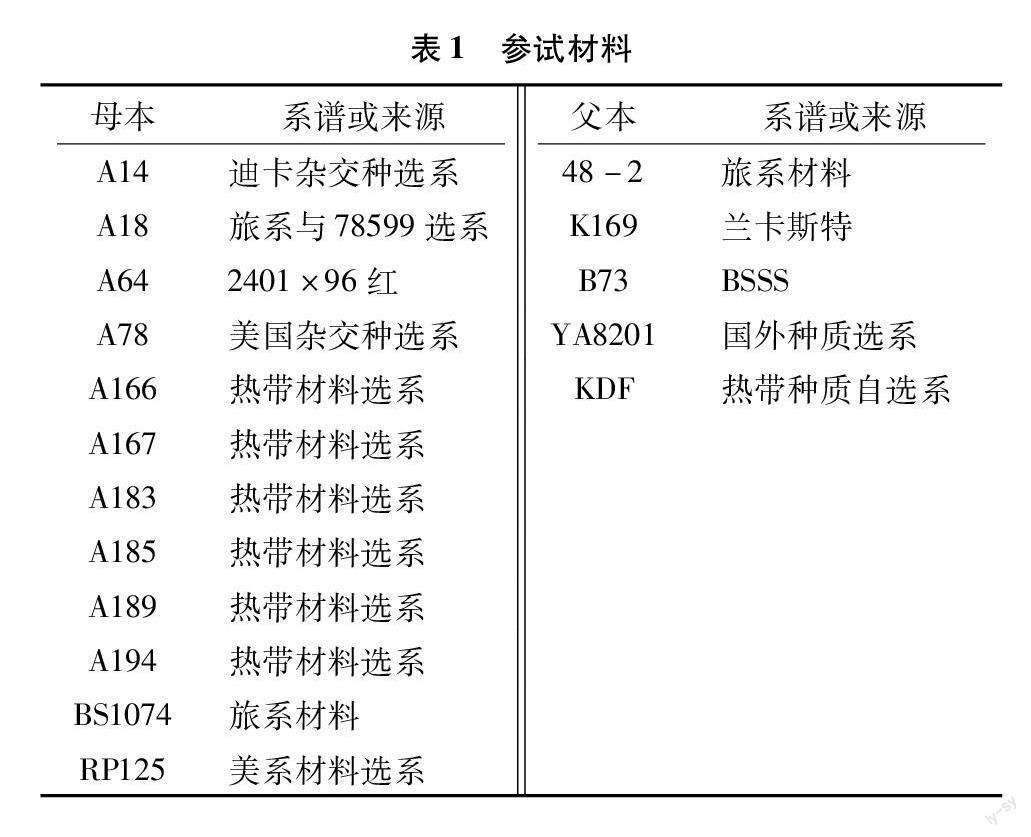
5个抗倒伏性能差异的自交系、12个常用及自选自交系按照不完全双列杂交(NCⅡ)遗传交配设计配制的60个杂交组合(表1),其中自交系A166、A167、A183、A185、A189、A194和KDF为热带材料经过辐照诱变后选系而来。
1.2 试验设计
2017年冬在海南三亚种植亲本并组配F1组合。2018年在四川德阳试验基地开展试验,田间设计为3行区种植小区,每个小区均采用随机区组设计。单株种植,每行14株,3次重复,田间管理同大田生产。小区种植密度为48 000株/hm2[18]。
1.3 性状调查及检测方法
在玉米花期结束后15 d进行株高、穗位高的测定,测定方法为田间生长条件下从泥土处到雄穗顶部和雌穗穗位节的长度。穿刺强度测定采用植物茎秆强度测定仪(石家庄世亚科技有限公司,型号SY-S03,量程5~500 N,分辨率0.1 N,精度±0.5%)对地上第3茎节和第5茎节中部茎秆进行穿刺并记录瞬间穿透茎秆表面韧皮组织的最大力数值[7]。纤维素、半纤维素及木质素含量的测定是取地上部分第3至第5茎节玉米茎秆,杀青后烘干并磨成细粉,具体测定方法参照文献[14]。
1.4 数据分析
以小区均数为单位,对收集的抗倒伏相关各性状进行方差分析;根据方差分析结果按不完全双列杂交设计对组合间差异显著的性状进行配合力分析[19]。数据统计分析由DPS 和Excel 2007软件完成。
2 结果与分析
2.1 抗倒伏相关性状的配合力方差分析
本研究的试验组合有60个,对其表征抗倒伏力的7个性状的数据结果进行方差分析,结果(表2)表明,所有组合间的抗倒伏性状均表现出极显著的遗传差异,同时父本×母本的特殊配合力方差也达到极显著水平。除株高和半纤维素含量2个性状以外,母本第3茎节穿刺强度、第5茎节穿刺强度、穗位高、纤维素含量、木质素含量的一般配合力方差均达到显著水平,尤其是穿刺强度、穗位高、木质素含量达到极显著水平。父本第3茎节穿刺强度、第5茎节穿刺强度、株高、穗位高的一般配合力方差达到显著水平,而纤维素含量、半纤维素含量和木质素含量差异不显著,表明本研究的所有性状均受非加性基因的控制。对各组合的第3茎节穿刺强度、第5茎节穿刺强度、穗位高进行分析发现,这3个性状受到父本和母本基因加性效应的双重影响;此外,母本基因加性效应对各杂交组合的纤维素含量和木质素含量2个性状有较显著的影响,而杂交组合的株高则受到父本基因加性效应的影响。对各组合半纤维素含量的特殊配合力分析显示其方差达极显著水平,但双亲的一般配合力方差并没有表现出差异,说明该性状双亲的互作效应明显。
2.2 抗倒伏相关性状的一般配合力效应分析
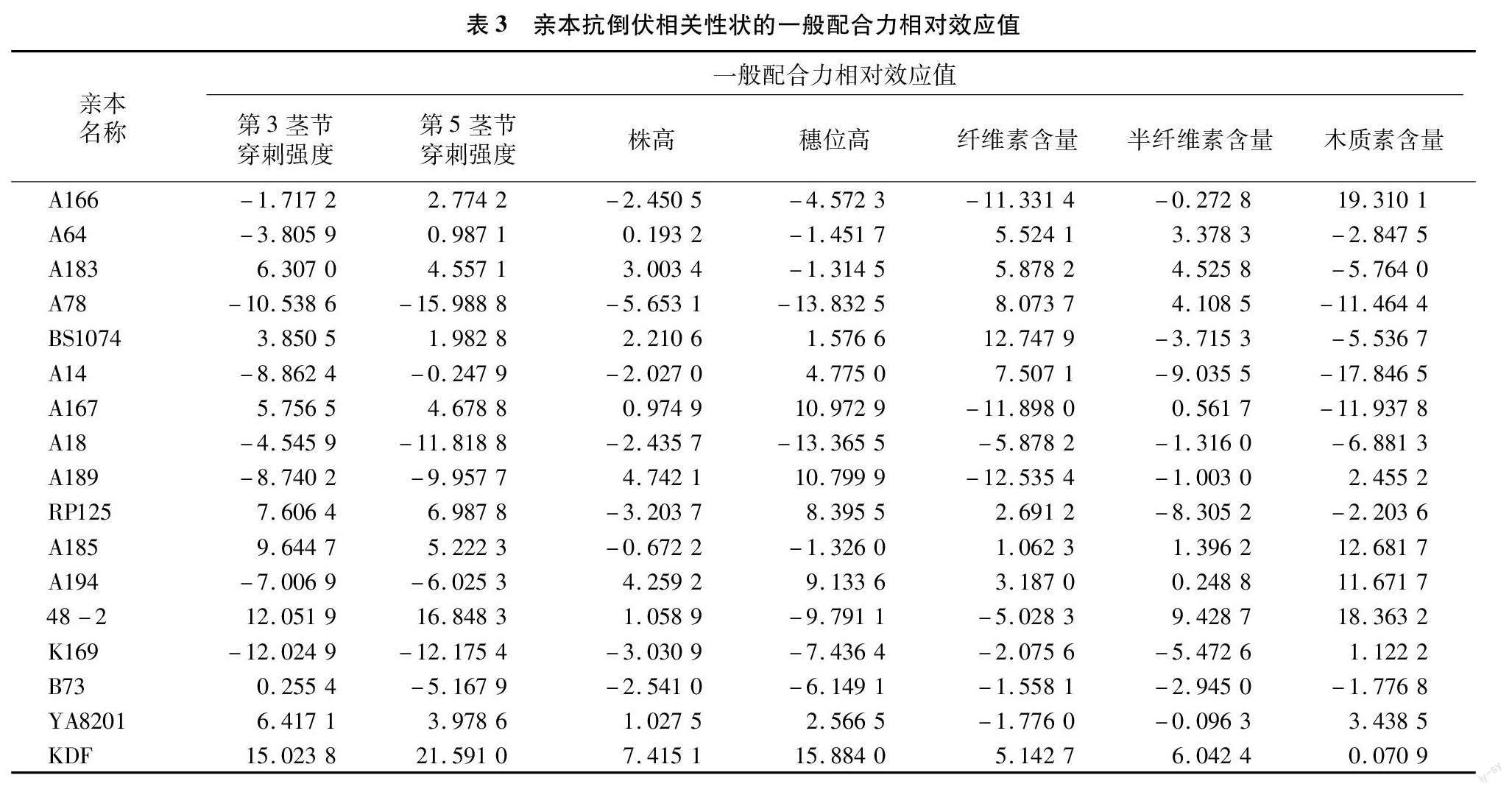
为了分析17个亲本的一般配合力,对其7个性状的相对配合力效应值进行分析,结果(表3)显示,亲本在所测试的7个性状上的一般配合力都存在较大的差异。从母本的各个性状的一般配合力来看,A185和RP125的第3茎节穿刺强度和和第5茎节穿刺强度的一般配合力表现最好,数值最大。自交系BS1074、A78和A14的纤维素含量一般配合力表现较优,A78和A183的半纤维素含量一般配合力表现较优,A166、A185和A194的木质素含量一般配合力表现较优,自交系A185和A194的纤维素含量、半纤维素含量和木质素含量的一般配合力均为正值,表现最优。株高和穗位高2个性状的一般配合力表现较好的自交系有A78、A18和A166,它们的一般配合力效应值均为负值,且最低。综合来看,RP125、A78和A185自交系的抗倒伏相关性状的一般配合力最优。
从5个父本各个性状的一般配合力来看,48-2和KDF的第3茎节穿刺强度和和第5茎节穿刺强度的一般配合力表现最好,数值最大。48-2、K169和B73的穗位高一般配合力表现较好,而K169和B73的株高一般配合力表现最优。仅自交系KDF的纤维素含量一般配合力为正值,48-2和KDF的半纤维素含量一般配合力表现较优,48-2和YA8201的木质素含量一般配合力表现较优,自交系KDF的纤维素含量、半纤维素含量和木质素含量的一般配合力均为正值,表现最优,而48-2也有2个指标为正值,表现同样较好。综合来看,父本自交系中,48-2和KDF的抗倒伏相关性状的一般配合力最优。
2.3 抗倒伏相关性状的特殊配合力效应分析
由表4可知,60个杂交组合的SCA相对效应值存在一定的差异,说明SCA在杂交一代中起到了一定的作用。
第3茎节穿刺强度性状上,SCA相对效应值变幅为-24.135 5~23.717 1,具有正向效应的组合有27个,前5位依次为A166×YA8201、A78×KDF、RP125×B73、A185×KDF、A78×YA8201,负效应前5位组合分别为A189×YA8201、A78×B73、A167×KDF、A183×KDF、A194×YA8201。
第5茎节穿刺强度上,SCA相对效应值变幅为-31.248 7~18.932 7,具有正向效应的组合有31个,前5位依次为A189×B73、A64×B73、A194×YA8201、A167×YA8201、A78×KDF,负效应前5位组合分别为A189×YA8201、A14×B73、A194×K169、A64×48-2、A183×KDF。
纤维素含量上,SCA相对效应值变幅为 -47.232 5~40.155 8,具有正向效应的组合有30个,前5位依次为A18×K169、A185×YA8201、A185×48-2、A78×B73、A18×KDF,负效应前5位组合分别为A185×B73、A18×48-2、RP125×K169、A167×KDF、A183×YA8201。
半纤维素含量上,SCA相对效应值变幅为 -41.157 1~27.130 5,具有正向效应的组合有31个,前5位依次为RP125×YA8201、A166×48-2、A189×YA8201、A183×48-2、A167×K169,负效应前5位组合分别为RP125×48-2、A189×B73、A78×K169、A166×YA8201、A183×KDF。
木质素含量上,SCA相对效应值变幅为 -22.617 0~21.111 1,具有正向效应的组合有35个,前5位依次为A183×48-2、A167×YA8201、A64×YA8201、BS1074×KDF、RP125×KDF,負效应前5位组合分别为A64×KDF、RP125×K169、A189×48-2、A64×B73、A183×YA8201。
株高性状上,SCA相对效应值变幅为-29.068 1~18.630 6,具有负向效应的组合有30个,前5位依次为A78×K169、A189×YA8201、A14×B73、A194×B73、A18×KDF,正效应前5位组合分别为A78×KDF、A189×B73、A14×K169、A189×KDF、A78×YA8201。
穗位高性状上,SCA相对效应值变幅为 -29.667 1~19.453 9,具有负向效应的组合有28个,前5位依次为A194×YA8201、BS1074×K169、A64×K169、A183×B73、A14×YA8201,正效应前5位组合分别为A18×B73、A185×K169、A194×B73、A78×K169、A194×KDF。
综合第3茎节穿刺强度、第5茎节穿刺强度、纤维素含量、半纤维素含量和木质素含量5个性状的SCA相对效应值,发现组合A183×48-2、A185×K169和A189×KDF表现最优,5个性状的SCA相对效应值均为正值;而组合A78×KDF、A185×KDF、A167×YA8201、BS1074×K169、A189×B73、A183×K169、A64×K169、A194×YA8201有4个性状的SCA相对效应值均为正值,也表现优异,可以选出抗倒伏能力强的组合。GCA高的亲本组合所得的后代SCA并不是所有组合中最高的,甚至有部分性状表现出负效应,说明这些性状也受非加性影响;相应地,部分GCA不高的亲本也可以因非加性效应组配出SCA较高的后代。
2.4 双亲在抗倒伏相关性状上的遗传力比较分析
为了评估双亲在抗倒伏性状上的遗传力,对亲本的广义遗传力和狭义遗传力均进行计算分析。茎秆相关性状的遗传力分析结果(表5)显示,狭义遗传率的大小表现为第5茎节穿刺强度>穗位高>第3茎节穿刺强度>木质素含量>株高>半纤维素含量>纤维素含量。所有性状的狭义遗传力均小于50%,说明这些性状不仅受遗传因素影响,可能还受到其他因素的影响。同时所有性状广义遗传力均大于狭义遗传力,说明这些性状受环境影响较大,特别是株高、纤维素含量和半纤维素含量这3个性状的广义遗传力远大于狭义遗传力。基于所有性状的狭义遗传率均小于50%。所以这些抗倒伏相关性状宜于在晚代进行选择。
3 讨论与结论
茎秆力学指标可用来评价茎秆抗倒性的强弱,主要包括穿刺强度、压碎强度和弯折强度,穿刺强度反映的是玉米茎秆皮层的性状[20]。穿刺强度不仅能够反映玉米茎秆抗病菌侵害和抗钻心虫破坏的能力[21-22],也与茎秆的抗倒伏能力呈正相关[23]。值得注意的是,研究表明玉米茎秆的第3茎节穿刺强度与倒伏率是极显著相关的[7,24]。本研究对地上第3茎节和第5茎节茎秆的穿刺强度进行了测定和分析,发现母本自交系A185和RP125的第3茎节和第5茎节穿刺强度的一般配合力表现最好,父本48-2和KDF的第3茎节和第5茎节穿刺强度的一般配合力表现最好,因此可将自选系自交系A185、RP125、48-2 和KDF作为配置抗倒伏杂交组合(高穿刺强度)的优良自交系和改良抗性的优质来源。
樊海潮等的研究结果不仅证实了穿刺强度与倒伏率具有紧密的关系,还证实了株高和穗位高与倒伏率呈极显著正相关[25];同时也有研究表明,矮秆品种之所以能在高密度种植条件下高产稳产,得益于其较好的抗倒伏性能[5]。在本研究中,自交系A78、A18、A166的株高、穗位高一般配合力表现最好,可在配置低株高、低穗位高的杂交组合中发挥作用。
纤维素、半纤维素和木质素等其他细胞壁成分相互交联形成结构复杂的细胞外基质增强了植物组织和细胞的机械强度,它们的含量多少决定了玉米茎秆强度的高低[26-28]。本研究中,自交系A185、A194、KDF和48-2的反映植物茎秆机械强度的细胞壁成分相关性状的一般配合力表现最优,其中A185、A194和KDF的一般配合力表现均为正值,这些自交系可作为配置玉米茎秆高纤维素、半纤维素和木质素含量组合的优良自交系或用于遗传改良。综合来看,本研究中自交系RP125、A185、48-2和KDF的抗倒伏相关性状的一般配合力最优,但自交系KDF的株高、穗位高一般配合力相对效应值均较高,这可能是该自交系选自热带种质,含有热带血缘较多的原因。
在本研究中60个杂交组合的SCA相对效应值存在一定的差异,说明SCA在杂交一代中起到了一定的作用,这与前人的其他农艺性状的研究结果[29]相似。所有性状的狭义遗传力均小于50%,暗示这些抗倒伏相关性状宜于在晚代进行选择。
本研究选取的几个倒伏相关性状表现在不同的自交系中各不相同,自交系A167的穿刺强度一般配合力均较优,但是其纤维素和木质素含量一般配合力均为负值。由于抗倒伏相关性状是相互影响的,在种植密度一定的情况下,玉米的穗位高和株高越小,玉米各株之间对彼此光合作用的影响越小,光能的利用越充分,越有利于玉米合成半纤维素和木质素,穿刺强度和抗倒伏能力越强[5,30]。与其他农艺性状和产量性状的研究一样,在玉米抗倒伏自交系和杂交组合的选育中,在选择这些性状的高GCA自交系作为亲本时,也要注重高SCA的选择,能够增加选择高抗倒伏的杂交组合的机会[31-32]。
参考文献:
[1]汪黎明,姚国旗,穆春华,等. 玉米抗倒性的遗传研究进展[J]. 玉米科学,2011,19(4):1-4.
[2]李得孝,康 宏,员海燕. 作物抗倒伏性研究方法[J]. 陕西农业科学(自然科学版),2001,47(7):20-22.
[3]薛 军,王克如,谢瑞芝,等. 玉米生长后期倒伏研究进展[J]. 中国农业科学,2018,51(10):1845-1854.
[4]中國机械工业联合会. 玉米收获机械 技术条件:GB/T 21962—2008[S].北京:中国标准出版社,2008.
[5]任佰朝,李利利,董树亭,等. 种植密度对不同株高夏玉米品种茎秆性状与抗倒伏能力的影响[J]. 作物学报,2016,42(12):1864-1872.
[6]潘志远. 不同株高玉米植株性状和抗倒伏特性及杂种优势研究[D]. 石河子:石河子大学,2021:24-25.
[7]丰 光,刘志芳,李妍妍,等. 玉米茎秆耐穿刺强度的倒伏遗传研究[J]. 作物学报,2009,35(11):2133-2138.
[8]马德志,于乔乔,孙玉珺,等. 黑龙江省三、四积温带玉米新品种抗倒伏性比较研究[J]. 西南农业学报,2019,32(8):1692-1700.
[9]Robertson D,Smith S,Gardunia B,et al. An improved method for accurate phenotyping of corn stalk strength[J]. Crop Science,2014,54(5):2038-2044.
[10]任安然,武丽芬,关红辉,等. 不同种植密度下玉米茎秆纤维性状和抗倒性相关分析[J]. 植物遗传资源学报,2017,18(4):653-664.
[11]关红辉,刘文斯,郭晋杰,等. 不同杂种优势群玉米茎秆纤维品质及配合力分析[J]. 植物遗传资源学报,2018,19(5):925-936.
[12]童淑媛,杜震宇,李春杰,等. 三江平原不同玉米品种茎秆特性比较与抗倒伏品种筛选[J]. 分子植物育种,2020,18(17):5860-5868.
[13]郑孟静,翟立超,申海平,等. 种植密度对河北夏玉米生理成熟后茎秆抗倒能力及产量的影响[J]. 核农学报,2022,36(7):1434-1445.
[14]赵英善. 玉米茎秆结构性化合物变化与抗倒伏强度关系的研究[D]. 石河子:石河子大学,2015:22-23,33-35.
[15]耿文杰,李 宾,任佰朝,等. 种植密度和喷施乙烯利对夏玉米木质素代谢和抗倒伏性能的调控[J]. 中国农业科学,2022,55(2):307-319.
[16]于玮淇,王芙臣,杜伟嘉,等. 种植密度与化学调控对春玉米茎秆性状及抗倒伏能力的影响[J]. 玉米科学,2022,30(5):71-79.
[17]张嘉莉. 硅磷配施对玉米茎秆形态特征及抗倒能力和产量的影响[D]. 雅安:四川农业大学,2018:40-41.
[18]肖 梅,董绍斌,任在斌,等. 玉米茎秆第三和第五茎节穿刺强度遗传模型选择[J]. 四川农业科技,2020(7):51-54.
[19]宋旭东,黄小兰,张振良,等. 10个糯玉米自交系穗部性状配合力及其遗传特性分析[J]. 南方农业学报,2020,51(10):2447-2453.
[20]赵 雪,周顺利. 玉米抗茎倒伏能力相关性状与评价研究进展[J]. 作物学报,2022,48(1):15-26.
[21]Butro n A,Malvar R A,Revilla P,et al. Rind puncture resistance in maize:inheritance and relationship with resistance to pink stem borer attack[J]. Plant Breeding,2002,121(5):378-382.
[22]Martin S A,Darrah L L,Hibbard B E.Divergent selection for rind penetrometer resistance and its effects on European corn borer damage and stalk traits in corn[J]. Crop Science,2004,44(3):711-717.
[23]豐 光,景希强,李妍妍,等. 玉米茎秆性状与倒伏性的相关和通径分析[J]. 华北农学报,2010,25(增刊1):72-74.
[24]谷利敏,乔江方,张美微,等. 种植密度对不同耐密夏玉米品种茎秆性状与抗倒伏能力的影响[J]. 玉米科学,2017,25(5):91-97.
[25]樊海潮,顾万荣,尉菊萍,等. 植物生长调节剂增强玉米抗倒伏能力的机制[J]. 江苏农业学报,2017,33(2):253-262.
[26]Xue J,Zhao Y S,Gou L,et al. How high plant density of maize affects basal internode development and strength formation[J]. Crop Science,2016,56:3295-3306.
[27]Chen Y X,Chen J,Zhang Y F,et al. Effect of harvest date on shearing force of maize stems[J]. Livestock Science,2007,111(1/2):33-44.
[28]Appenzeller L,Doblin M,Barreiro R,et al. Cellulose synthesis in maize:isolation and expression analysis of the cellulose synthase (CesA) gene family[J]. Cellulose,2004,11(3):287-299.
[29]刘 倩,张国豪,车万均,等. 杂交水稻重要亲本农艺性状配合力遗传力分析[J]. 广东农业科学,2020,47(1):1-8.
[30]徐 幸. 种植密度对不同株高玉米品种茎秆抗倒伏性能及产量的影响[D]. 长春:吉林农业大学,2019:34-36.
[31]莫润秀,黄开健,黄爱花,等. 20个CIMMYT耐低氮玉米自交系主要性状的配合力分析[J]. 西南农业学报,2019,32(12):2732-2739.
[32]郭向阳,邬 成,陈泽辉,等. 玉米Suwan-Lancaster和Tuxpeno-Reid改良系的产量及相关性状配合力分析[J]. 西南农业学报,2016,29(12):2796-2799.
猜你喜欢
农业科技通讯(2023年1期)2023-02-12 07:09:28
纺织科技进展(2021年3期)2021-06-09 08:07:14
陶瓷学报(2021年1期)2021-04-13 01:33:02
上海包装(2019年8期)2019-11-11 12:16:14
猪业科学(2018年5期)2018-07-17 05:56:14
天津造纸(2016年1期)2017-01-15 14:03:29
河北农业科学(2016年2期)2016-09-09 01:41:31
中国造纸学报(2015年1期)2015-12-16 19:35:25
中国塑料(2014年4期)2014-10-17 03:00:45
应用化工(2014年11期)2014-08-16 15:59:13
