
嗜线虫肠杆菌抑制大蜡螟幼虫的免疫反应
2023-08-10 08:07廖春丽杨校飞李美换黄亚鹏马安琪陈丙艳陈孟娇李冰冰
河南城建学院学报 2023年3期
廖春丽,黄 冉,杨校飞,李美换,张 凯,黄亚鹏,马安琪,陈丙艳,陈孟娇,李冰冰
(1.河南城建学院 生命科学与工程学院,河南 平顶山 467036;2.河南城建学院 河南省健康食品工程技术研究中心,河南 平顶山 467036;3.河南城建学院 河南省水体污染防治与修复重点实验室,河南 平顶山 467036;4.河南久星生物科技研究院有限公司,河南 平顶山 467036)
利用大蜡螟(Galleria mellonella)诱集到致病性高的昆虫病原线虫(Heterorhabditissp.PDSj2),从其肠道内分离出1株具有较高杀虫活性的共生菌株NK(鉴定为新属阴沟肠杆菌)[1]。文献[2]发现共生菌高效的杀虫活性与其作用机制密不可分,其产生的各种毒素蛋白和免疫抑制因子会破坏昆虫的免疫系统,最终将其杀死,另外共生菌侵入宿主后快速繁殖也会大量消耗宿主血淋巴中的能源物质。
血淋巴中含有昆虫生理活动所必需的一切物质(如蛋白质、糖等生物大分子)。生物大分子含量的异常变化会直接影响昆虫的正常生理功能,导致昆虫生长发育受阻,甚至死亡。丁晓帆等[3]研究表明,昆虫病原线虫会明显改变大蜡螟的血淋巴总量及血淋巴蛋白含量、酯酶活性。杨君等[4]研究表明,注射嗜线虫致病杆菌血腔毒素后,大蜡螟体内羧酸酯酶和乙酰胆碱酯酶活力都明显升高,而酚氧化酶活力受到抑制。肖猛[8]研究表明,昆虫病原线虫能明显改变寄主昆虫血淋巴中蛋白含量、血糖含量和酯酶活性。
昆虫采用精确复杂的负调控机制调节Imd信号通路免疫系统,负调控因子同时受Imd信号通路所调控[5]。负调控因子主要是一类具有酰胺酶活性的PGRPs家族蛋白(PGRP-LB、PGRP-SB1、PGRP-SB2、PGRP-SC1a、PGRP-SC1b、PGRP-SC2)[6-7]。已有研究表明昆虫肠道Imd信号通路在昆虫肠道的体液免疫防御中起着不可或缺的作用。姚志超[8]在研究中发现,Imd信号通路在橘小实蝇系统免疫反应中发挥重要作用,大肠杆菌DH5α的系统侵入能够迅速激活Imd信号通路的免疫响应且在Imd信号通路中存在负调控作用。肖蓉[9]在对胞质型肽聚糖识别蛋白等的研究中发现肠道抗菌肽区域化表达也是一种调控Imd信号通路的重要机制。
广东海事局局长陈毕伍表示,广东毗邻东南亚,水网发达,人们水上日常出行、旅游等活动频繁,广东珠江口海域和琼州海峡涉客渡运近10年来保持零事故、零死亡纪录,得益于广东致力建立安全、便捷、高效、绿色的现代综合交通运输体系和共商共建共享共治的安全社会治理体系。愿与各国分享“广东经验”,共同为亚太地区的渡运安全和人民群众安全出行贡献力量。
虽然关于线虫对大蜡螟等多种昆虫的致病机理研究取得了一些进展,但是昆虫病原线虫共生菌侵入对大蜡螟生理及病理影响的研究很少。本文对嗜线虫肠杆菌侵入过程中大蜡螟幼虫酶活性、能源物质含量及基因表达量的变化进行研究,初步探讨嗜线虫肠杆菌的杀虫机理。
把教书当“饭碗”,混口饭吃。认为在中小学思品、政治课是“副科”,应付一下上级、应付一下学生、应付一下中考就行了,对于党的十八大明确指出的“把立德树人作为教育的根本任务”[2]置若罔闻,教师“立德树人”的主导作用少见踪影。
利用已有的精细化学技术、软件开发技术、涂层技术、图像技术,爱克发印艺制定了将技术转化为为客户创造价值的可持续解决方案的战略目标,布局相关市场和业务群。据爱克发感光器材(深圳)有限公司产品与市场总监葛玉军介绍,市场方面,爱克发印艺在坚守既有的平版胶印和宽幅打印市场之外,持续发力包括智能包装、安全印刷和工业印刷在内的新市场;业务群方面,则按印前(商业和报业印前产品、安全与防伪印刷产品),喷墨(设备、油墨、介质),印刷耗材(胶印油墨、车房化学品和光油、橡皮布),服务(印刷专家服务、云端软件服务)四大业务群划分。上述布局涵盖印前到印刷、硬件到软件、产品到服务,贯穿整个产业链。
1 材料与方法
1.1 材料
1.1.1 嗜线虫肠杆菌NK
将其分离出后,保存在实验室待用。
本文以31个省份为研究对象,对1978~2017年我国要素禀赋重心和经济重心耦合趋势进行研究,从而更好地刻画了我国要素禀赋分布格局的动态演进过程。结果显示,改革开放以来我国劳动力重心较为稳定,资本重心表现出阶段性移动特征,劳均资本重心在不同时期空间上呈相对稳定的渐进式变化。
(1)血淋巴酚氧化酶(PO)活性和酯酶活性测定。
采用考马斯亮蓝法G-250法测定血淋巴蛋白含量,即每毫升血淋巴中蛋白的质量(单位为mg/mL)。
(3)发酵培养基:蛋白胨2.1%,葡萄糖0.6%,MgSO40.13%,Na2SO40.15%,(NH4)2SO40.23%,KH2PO40.072%,K2HPO40.098%;pH调至7.2~7.4。
1.1.3 供试昆虫
证明 (1) 由于A是完全确定性自动机,所以A不会影响监控器M中的概率计算.另外,如果H中某一状态不包含在M的组合状态中,则该状态与监控属性无关,也不会影响监控器M中的概率计算.所以,
分别选取若干只大蜡螟3龄幼虫(Galleria mellonella larvae)作为实验组和对照组。实验组用含有嗜线虫肠杆菌的饲料(10 mg/g)喂养;对照组用普通饲料喂养。饲养温度为25~30 ℃,湿度为75%,避光。
1.1.4 饲料
引物为北京生工公司合成(引物序列见表1)。荧光定量试剂盒选用北京聚合美生物科技有限公司的2x M5 HiPer SYBR Premix Es Taq。将反转录cDNA作为模板,按照荧光定量反应体系(qPCR mix 5 μL,cDNA 1 μL,primer F 0.3 μL,primer R 0.3 μL,ddH2O 3.4 μL,共10 μL),在LightCycler480荧光定量仪器反应,反应程序设定为:95 ℃ 2 min; 95 ℃ 5 s; 60 ℃ 30~34 s; 45个循环。结果分析采用2-ΔΔCT法。
随机选取每个时间点的实验组和对照组大蜡螟幼虫各30头,在无菌操作条件下麻醉,准备解剖。
谈到改革开放以来公司的发展壮大,山河智能工会主席张爱民如数家珍。得益于国家加大对基础设施建设的投入,公司快速发展。目前,公司已跻身全球工程机械企业500强、世界挖掘机企业50强、世界支线飞机租赁前3强,产品畅销国内外,出口100多个国家和地区。
1.2 方法
1.2.1 嗜线虫肠杆菌菌体细胞制备
将活化的初生型嗜线虫肠杆菌NK接种到LB液体培养基中,恒温(28 ℃)、振荡(180 r/min)培养24 h,得到种子菌液。将种子菌液按照6%的接种量接种到发酵培养基中,恒温(28 ℃)、振荡(180 r/min)发酵培养28 h,收集发酵液并在4 ℃、10 000 r/min的条件下离心10 min,把菌体细胞按照10 mg/g的比例添加到人工饲料中,放置于冰箱(4 ℃)中保存备用。
1.2.2 大蜡螟幼虫的血淋巴酚氧化酶和酯酶活性、蛋白质及总糖含量的测定
每隔12 h取一次实验组和对照组的血样(血淋巴),将血样放入eppendorf管内并加少许苯基硫脲以防止氧化,冻贮待用。
(1)NA培养基:牛肉膏 0.3%,蛋白胨0.5%,营养琼脂2%,pH 7.2~7.4。
PO活性测定参照文献[10]的方法,在紫外分光光度计上测定其在495 nm下的吸光值变化,其活性大小定义为5 min内的吸光值变化。羧酸酯酶活性测定采用陈长琨[11]的方法。
细究下去,我发现当他们在早年只能获得一份打扫办公室卫生的工作时,他们只会全力以赴,不去挑剔工作的好坏,他们不仅干得很快乐,而且对周围人非常关爱和尊重,总是以积极正面的态度处理人际关系和工作环境,结果自然处处和谐融洽,成功也便孕育其中了。
(2)血淋巴蛋白含量测定。
(2)LB培养基:蛋白胨10%,酵母粉5%,氯化钠10%,pH 7.2~7.4。
(3)血淋巴总糖含量测定。
采用陈毓荃[12]蒽酮比色法测定大蜡螟幼虫血淋巴总糖含量,即每毫升血淋巴中葡萄糖的质量(单位为mg/mL)。
1.2.3 大蜡螟幼虫中肠制备
首先将解剖大蜡螟幼虫所用的镊子、刀等物品用DEPC无酶水浸泡24 h,风干,锡纸包裹高压灭菌20 min。随机选取每个时间点的实验组和对照组大蜡螟幼虫各30头。无菌操作条件下,在超净工作台上铺一层锡箔纸(用酒精棉擦拭一遍,再喷上一层核酸保护剂),把上述灭菌的物品放在上面,然后将麻醉后的大蜡螟幼虫放入乙醇中浸泡数秒,并用无菌生理盐水涮洗后,用手术刀快速取出肠道组织,立刻放入无 RNase 离心管中,拧紧盖子,放入液氮中冷冻10 min,取出放入冰箱(-80 ℃)待用。
1.2.4 大蜡螟幼虫中肠Duox基因实时荧光定量PCR
每一个人的内心深处都希望得到别人的肯定与赞美,更何况幼儿,他们最希望教师、家长的赞许,教师一个赞许的眼神,一句真诚的表扬可能会让他们兴奋一整天。幼儿在混班游戏活动中,特别希望在别的小朋友面前得到肯定,正如叶圣陶老先生所说,骄傲的架子只能在同伴面前摆。教师要抓住每个幼儿的闪光点适时进行表扬,给他们一个赞,一朵小红花,一颗泡泡糖,他们在游戏中会更加卖力,就连中午吃饭睡觉都会格外乖巧。
(1)RNA的提取及检测。
采用Trizol法,用小分子核酸测定仪(Nanodrop-2000,赛默飞)检测RNA的浓度和纯度。
1.1.2 嗜线虫肠杆菌培养基
(2)反转录。
采用反转录试剂盒(ReverTra Ace® qPCR RT Master Mix,TOYOBO)进行反转录。
从世界范围来看,过去30多年的水市场浪潮,并未改变水分配和管理政府主导的格局。目前运作良好的水权市场只是局限于美国、澳大利亚等少数发达国家的部分地区。一些国家特别是发展中国家,尽管开展了水权市场的探索,但是由于水权法规不健全、制度执行不力及客观条件制约,水权市场的运作面临很多问题,政府需要大量干预和介入,水权市场因而具有“准市场”的特征。至于水务市场化的改革,普遍被认为并不成功,进入新世纪以来日渐式微。目前全世界85%以上的水设施在政府控制之下。
(3)qRT-PCR 反应。
(1)中国对 “一带一路”沿线国家机械运输设备产品出口的种类比较丰富,且在各国的分布差异在逐步缩小。从图1中可以看到,2007~2017年,扩展边际的核密度图峰值均接近于1,基本在 (0.9~1)之间,这说明中国在 “一带一路”沿线国家的出口种类比较齐全。2007~2012年,核密度图的峰值越来越高且曲线位置微微右移,说明中国对 “一带一路”沿线国家的出口产品种类差距越来越小;由于2012~2016年机械行业市场低迷,所以核密度图的峰值波动下降,意味着各国的扩展边际差距稍有变化,但变化幅度并不大。2017年核密度图的峰值稍有升高,机械行业市场稍有回暖。
1.1.5 大蜡螟幼虫的处理

表1 引物序列
1.3 数据分析
使用Excel 2018和SPASS 13.0分析软件进行数据分析,采用t测验方法对嗜线虫肠杆菌处理组和对照组之间的能源物质含量、酶活性和PGRP-LB基因表达量进行差异性检验。
2 实验结果
2.1 大蜡螟幼虫机体表观变化
在饲喂含有嗜线虫肠杆菌的饲料后,大蜡螟幼虫机体迅速发生免疫反应(见图1)。在12 h时机体普遍出现斑点,大蜡螟幼虫活力下降;在36 h时有些大蜡螟幼虫的机体稍微发黑;在60 h时机体出现黑化现象,有些大蜡螟幼虫死亡;在84 h时所有大蜡螟幼虫死亡,大多数大蜡螟幼虫通体发黑。

图1 饲喂过程中大蜡螟幼虫机体表观变化
2.2 大蜡螟幼虫血淋巴酚氧化酶和羧酸酯酶活性变化
大蜡螟幼虫血淋巴酚氧化酶和羧酸酯酶活性变化分别如图2、图3所示。由图2可知,对照组的血淋巴酚氧化酶活性基本没有变化,实验组的酚氧化酶活性呈现“急速上升—急速下降”的趋势,在36 h时酚氧化酶活性达到最高,然后出现明显的下降。由图3可知,对照组的血淋巴羧酸酯酶活性基本没有变化;而实验组的羧酸酯酶活性呈现“缓慢上升—缓慢下降”的趋势,在48 h时羧酸酯酶活性达到最高,然后出现明显的下降。
(1)信息技术安全弱。技术风险是互联网金融相比传统金融尤为突出的一种风险。由于其载体是互联网,因此本身在互联网行业就十分显著的技术风险便传导至互联网金融理财领域。互联网金融企业的用户信息全部存放在互联网平台上,安全极难得到保障,一旦被黑客攻破,数据信息便可被任意修改或删除,从而造成无法估量的损失。造成互联网金融技术风险的主要原因是互联网金融平台由于资金短缺,无法做到设备的物理隔离、数据信息的备份和不同机房间的无缝切换。
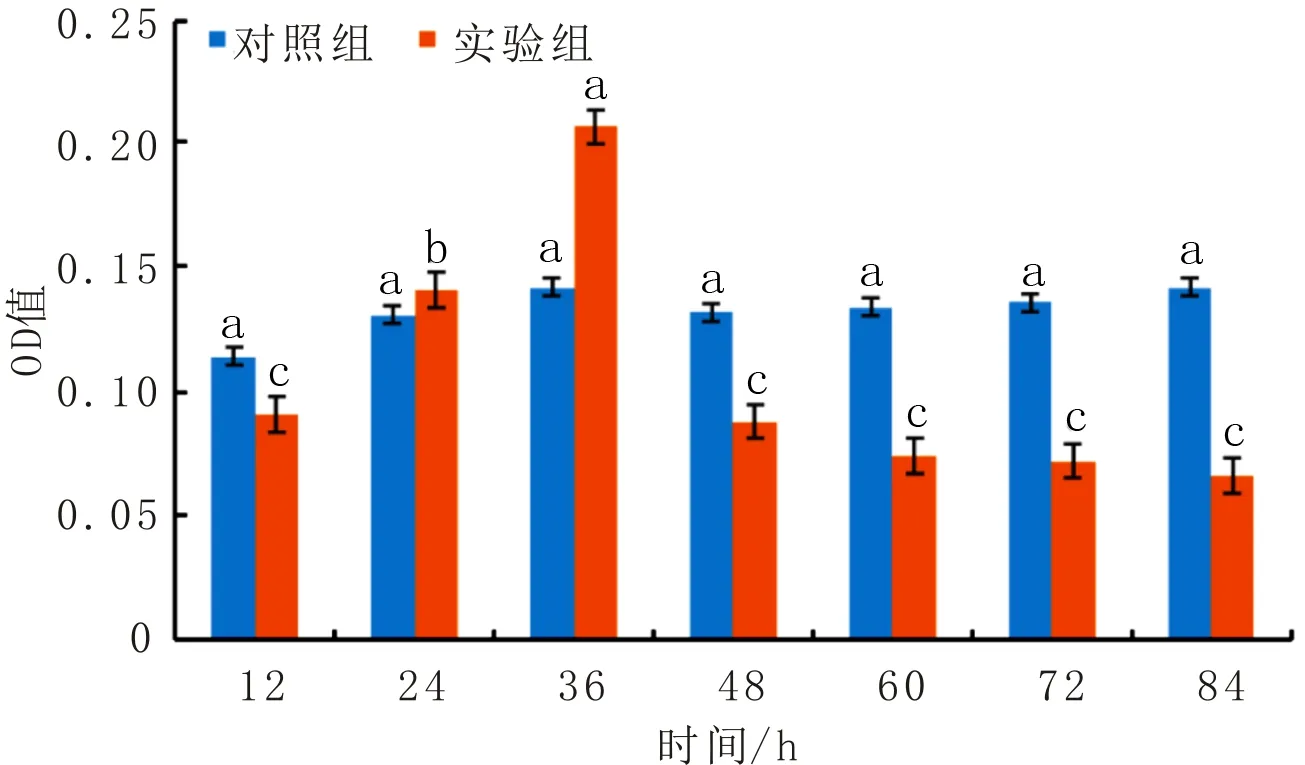
图2 大蜡螟幼虫血淋巴酚氧化酶活性变化

图3 大蜡螟幼虫血淋巴羧酸酯酶活性变化
2.3 大蜡螟幼虫血淋巴血糖和蛋白含量变化
大蜡螟幼虫血淋巴血糖和蛋白含量变化分别如图4、图5所示。由图4可知,对照组的血淋巴血糖含量在84 h内基本不变,但是实验组血淋巴血糖含量急剧下降。由图5可知,对照组的血淋巴蛋白含量呈缓慢上升趋势,但在72 h时又下降到最初水平,实验组的血淋巴蛋白含量明显高于对照组,而且持续增加。

图4 大蜡螟幼虫血淋巴血糖含量变化
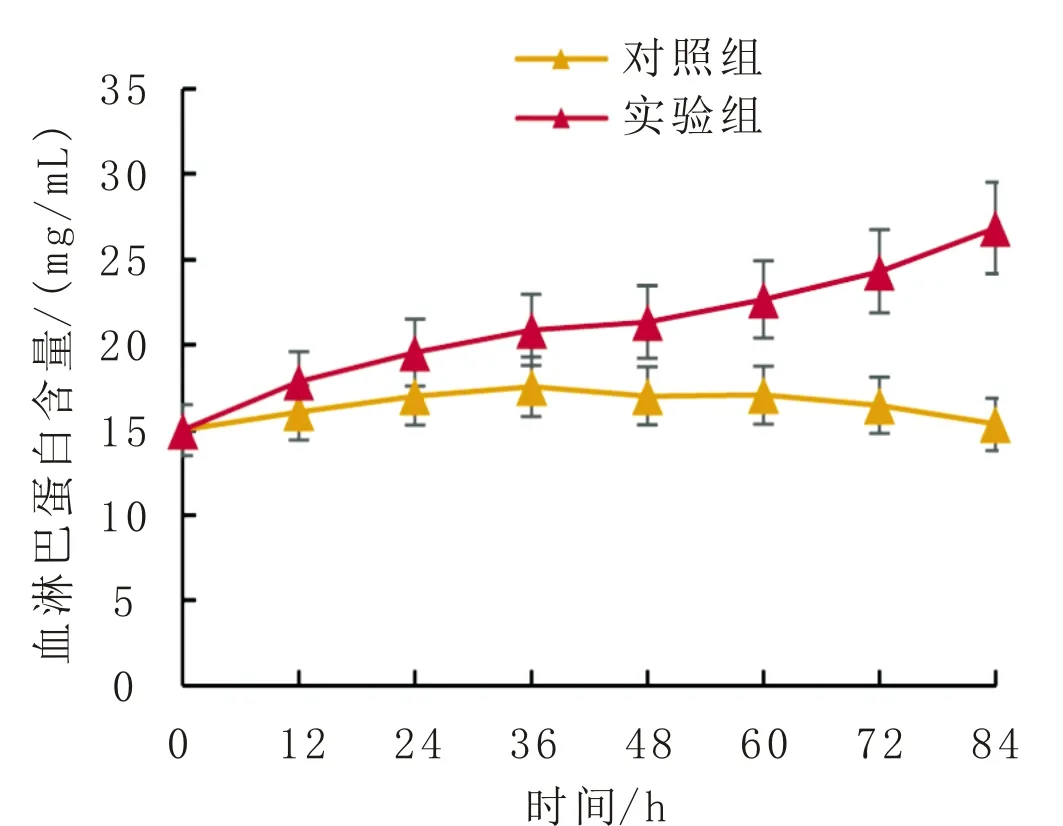
图5 大蜡螟幼虫血淋巴蛋白含量变化
2.4 PGRP-LB基因的荧光定量PCR
(1)引物合成PCR检测。
例1~例6“不”修饰谓语,在例7中,“不”位于句末,构成反复问句,此种用法《周》共17见,《齐》中无此用法。
目的基因引物和内参基因引物PCR电泳图如图6所示。由图6可知,目的基因PGRP-LB为208 bp,内参基因α-Tubulin为184 bp,与marker对比大小基本符合,目的基因和内参基因电泳条带清晰明亮,可以用于荧光定量PCR。
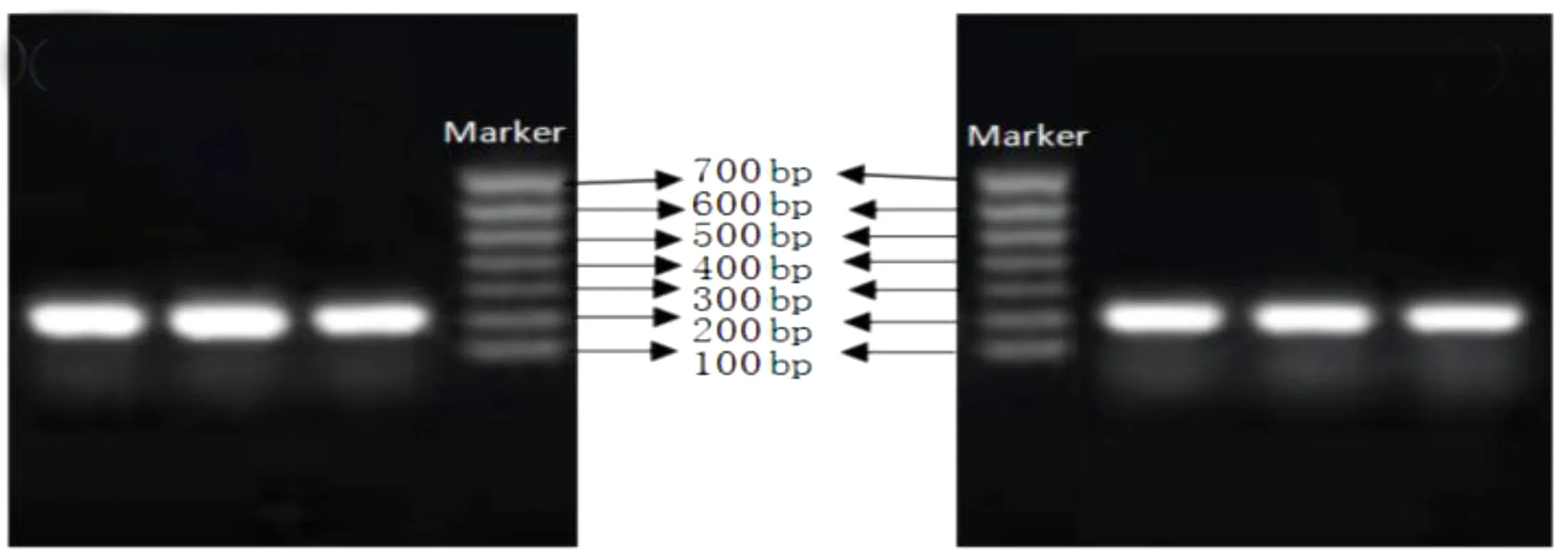
a 目的基因 b内参基因
(2)荧光定量PCR检测。
大蜡螟幼虫肠道PGRP-LB基因表达如图7所示。在图7 a中,大蜡螟幼虫在取食含有嗜线虫肠杆菌饲料后,其肠道内PGRP-LB基因的表达量在18 h时显著上升,与对照组相比上升了6.07倍,在18 h后其肠道内PGRP-LB基因的表达量开始下降。在图7 b中,在24 h以后停止饲喂含有嗜线虫肠杆菌饲料,虽然大蜡螟幼虫的肠道内PGRP-LB基因的表达量迅速下降,但仍然低于正常含量。在36 h后,在大蜡螟幼虫的Imd信号通路的免疫调节下,其肠道内PGRP-LB基因的含量开始恢复正常。推测大蜡螟幼虫具有强大的自身免疫调节能力,同时嗜线虫肠杆菌的侵入会强烈影响PGRP-LB基因的表达。
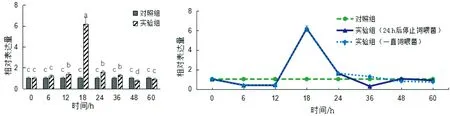
a 嗜线虫肠杆菌侵入后 b 嗜线虫肠杆菌停止侵入后
3 结论与讨论
昆虫一般通过产生黑色素来抵御外来物的侵入[13]。酚氧化酶(PO)是昆虫体内黑色素合成的关键酶,在昆虫免疫反应中起关键性作用[14]。本文中,随着嗜线虫肠杆菌的侵入,大蜡螟幼虫的体表颜色呈现从黑色斑点到通体发黑的变化,血淋巴中酚氧化酶活性呈现“急速上升—达到峰值—急速下降”的变化,血淋巴中酯酶活性也呈现“上升—达到峰值—下降”的变化,酯酶活性峰值到达时间晚于酚氧化酶活性峰值时间。发生这种变化可能是由于嗜线虫肠杆菌侵入大蜡螟幼虫体内之后,引起免疫反应,免疫系统被激活导致酶活性升高,但是随着肠杆菌不断侵入,多酚氧化酶级联反应产生的有毒物质大量堆积以及肠杆菌对大蜡螟幼虫免疫系统的攻击,导致其免疫功能下降,多酚氧化酶活性降低;同时,随着肠杆菌不断侵入,大蜡螟幼虫的活动能力降低,取食变得困难,需要分解一些贮存的能源物质来满足自身代谢的需求;另一方面大蜡螟幼虫免疫系统启动后,合成免疫蛋白等可能进一步消耗了其体内储备的营养物质,致使酯酶活性峰值到达时间晚于酚氧化酶活性峰值时间。
肖猛[15]发现受到斯氏线虫感染的家蚕,其血淋巴中蛋白质含量在12 h后明显快速下降。高志华等[16]发现嗜菌异小杆线虫H.bacteriophoraE-6-7感染雌性血蜱12 h后血蜱血淋巴蛋白含量升高,在48 h后又显著降低。丁晓帆等[3]用不同病原线虫感染大蜡螟24 h后,对照大蜡螟的血糖含量基本不变,而处理大蜡螟的血糖则因线虫品系的不同而有不同程度的降低。本文中,嗜线虫肠杆菌的侵入使大蜡螟幼虫体内血淋巴血糖含量迅速下降,这种变化可能是由于嗜线虫肠杆菌侵入大蜡螟幼虫体内后,使大蜡螟幼虫的糖类合成代谢受阻,因而血淋巴总糖含量下降。嗜线虫肠杆菌的侵入使大蜡螟幼虫体内血淋巴蛋白含量初期也缓慢增加,推测是在血淋巴中产生一些与免疫机制相关的大分子酶类,使蛋白含量有所提升,后期血淋巴蛋白含量继续升高,这是由于嗜线虫肠杆菌大量繁殖,造成大蜡螟体液免疫系统失效,其组织被破坏,释放出更多蛋白。
姚志超[8]研究发现橘小实蝇取食大肠杆菌 DH5α后,PGRP-LB基因表达量在1 h和6 h时分别上调了2.82倍和1.36倍,在1 h时变化比较迅速。本文中,大蜡螟幼虫在取食含有嗜线虫肠杆菌10 mg/g浓度的人工饲料后,肠道内PGRP-LB基因的表达量在18 h时显著上升,比空白组上调6.07倍,相比姚志超的研究结果,本文中大蜡螟对于嗜线虫肠杆菌的侵入,反应较为缓慢,但反应更加强烈。总之,大蜡螟受到嗜线虫肠杆菌的侵入后,体内免疫防御启动,Imd信号通路开始发挥作用,前期PGRP-LB基因表达量显著上升,后期虽然PGRP-LB基因表达量下降,但是免疫防御无法进行恢复,最终造成大蜡螟死亡。
有研究发现,在昆虫的体液免疫中存在负调控现象,防止免疫过激对机体造成损害。本文实验在嗜线虫肠杆菌感染大蜡螟幼虫24 h后,停止饲喂嗜线虫肠杆菌,PGRP-LB基因含量在24~36 h继续降低,推测是由于为防止免疫过激对机体造成损害,负调控机制启动,PGRP-LB基因减少(后续可以采用基因沉默进行验证),之后PGRP-LB基因含量恢复正常,而且大蜡螟幼虫不再出现大量死亡现象。说明在恰当的时间停止嗜线虫肠杆菌对大蜡螟的侵入,通过自身免疫调节,大蜡螟幼虫体内稳态可以恢复正常。
猜你喜欢
世界最新医学信息文摘(2021年12期)2021-06-09
原子与分子物理学报(2021年1期)2021-03-29
知识窗(2019年9期)2019-10-09
蜜蜂杂志(2019年12期)2019-06-16
天然产物研究与开发(2018年8期)2018-09-10
天然产物研究与开发(2018年4期)2018-05-07
广西林业科学(2016年4期)2016-03-16
听力学及言语疾病杂志(2015年5期)2015-12-24
哈尔滨医药(2015年3期)2015-12-01
中国蜂业(2014年6期)2014-05-25
