
贺兰山蒙古扁桃灌丛根际土壤AM 真菌群落多样性特征研究
2023-08-10 06:15侯晖颜培轩谢沁宓赵宏亮庞丹波陈林李学斌胡杨梁咏亮倪细炉
生态环境学报 2023年5期
侯晖,颜培轩,谢沁宓,赵宏亮,庞丹波,4,陈林,4,李学斌,胡杨,4,梁咏亮,倪细炉,4*
1.宁夏大学生态环境学院/西北土地退化与生态恢复国家重点实验室培育基地,宁夏 银川 750021;2.宁夏大学/西北退化生态系统恢复与重建教育部重点实验室,宁夏 银川 750021;3.宁夏大学农学院,宁夏 银川 750021;4.宁夏贺兰山森林生态系统定位观测研究站,宁夏 银川 750021;5.宁夏贺兰山国家级自然保护区管理局,宁夏 银川 750021
丛枝菌根真菌(arbuscular mycorrhizal fungi,AMF)是一种存在于根际土壤中,能与陆地上大多数维管植物形成互利互惠的关系的菌根真菌(Smith et al.,2010),除十字花科、蓼科、莎草科、苋科、石竹科等10 余科植物外,广泛存在于草地、灌木等生态系统,是脆弱和退化生态系统植被恢复过程和维持生态系统功能的重要组成部分(王晓龙等,2016)。
AMF 通过根外菌丝扩大根系的吸收面积,可以有效地改善土壤结构和保持水土的功能,还能减少土壤中的养分淋失,提高植物对土壤水分和碳、氮、磷、钾等养分的吸收作用(李芳等,2016;王邵军等,2019),在干旱、贫瘠的土壤中显著增强宿主植物抗逆性(王琚钢等,2014),通过促进植被群落中的亚优势种、稀有种或少见种的生长来增强草地植被多样性(Krüger et al.,2017)。植被群落的丰富度越高,AMF 的多样性越高。土壤、气候和地理因子等因素对AMF 群落分布也有影响(Klichowska et al.,2019)。
贺兰山生态系统是西北干旱半干旱地区重要的生态屏障,其植被为贺兰山生态系统安全和区域生态系统服务提供了重要保障。宁夏贺兰山自然保护区内有着丰富的耐旱灌木和草本植物,是中低海拔主要的植被类型,成为维系荒漠地表景观及系统稳定性的重要基础。其中蔷薇科李属的蒙古扁桃(Prunusmongolica)和禾本科针茅属的短花针茅(Stipabreviflora)为低山区优势种和建群种,是重要的防风固沙植物之一,两者能在营养贫瘠的干旱荒漠土壤环境中共存,形成典型的蒙古扁桃-短花针茅群落。研究表明,AMF 能与蒙古扁桃和短花针茅形成良好的共生关系(王琚钢等,2011;格根宝乐尔,2013);灌木可作为护生植物(nurse plant)保护近邻植物,改善土壤养分,在灌木周围形成“沃岛”效应,进而形成有利于微生物生存的生境(Varela et al.,2017;Schöb et al.,2018);但目前,该区域共生的蒙古扁桃灌丛根际土壤养分及其AMF 群落结构特征尚不清楚,因此,本研究以宁夏贺兰山自然保护区低海拔地区蒙古扁桃灌丛典型的蒙古扁桃-短花针茅群落为研究对象,设置去除蒙古扁桃或短花针茅形成的单一植被样地作为对照,探明蒙古扁桃灌丛群落与单一植被间的根际土壤养分及AMF 群落结构变化特征,揭示其根际土壤AMF 群落多样性及其影响因子,为贺兰山干旱半干旱区生态系统功能的养护和稳定以及植被的修复提供理论参考。
1 材料与方法
1.1 实验区概况
实验点位于宁夏回族自治区银川市贺兰山东坡响水沟小流域,处于银川平原和阿拉善高原之间(地处38°27′-39°30′N,105°41′-106°41′E 之间),属典型大陆性气候,具有山地气候特征。年均气温−0.8 ℃,年均降水量420 mm,年均蒸发量2 000 mm,其降水主要集中在6-8 月,占全年降水量的60%-80%。研究区主要植物种有蒙古扁桃(Prunus mongolica)、小叶金露梅(Potentillaparvifolia)、狭叶锦鸡儿(Caraganastenophylla)、短花针茅(Stipa breviflora)、戈壁针茅(Stipatianschanicavar.gobica)和蒙古冰草(Agropyronmongolicum)等。
1.2 试验设计
于2021 年春季4 月,在贺兰山东坡海拔约1 700 m 处,选择15 个植被长势均一的蒙古扁桃-短花针茅(G×S)群落为设置样地,样地内无其他植物生长。样地面积为5 m×5 m,设置3 个处理:(1)5 个样地去除植被中的短花针茅植株(S);(2)5 个样地去除植被中的蒙古扁桃植株(G);(3)5个样地保持蒙古扁桃-短花针茅群落体系(蒙古扁桃:G×S−S,短花针茅:G×S−G)。2021 年7 月,利用工具和五点采样法将样地内的植株根系尽可能保持完整地挖出,以抖土法取附着在根际的土壤,混合均匀后,放入灭菌袋中,土样一部分存于−80 ℃冰箱保存,用于土壤AMF 群落多样性的测定;一部分自然风干后过筛,用于土壤理化性质测定(表1)。

表1 样地基本信息Table 1 Basic information of the plot
1.3 测定项目及方法
土壤理化性质测定参考《土壤农化分析》(鲍士旦,2000),土壤pH 采用酸度计电位法测定,水土质量比为5꞉1;土壤有机质(organic matter,OM)采用重铬酸钾氧化外加热法测定;土壤全氮(total nitrogen,TN)采用半微量凯氏法测定;土壤碱解氮(alkali-hydrolyzable nitrogen,AN)采用碱解扩散法测定;土壤全磷(total phosphorus,TP)采用HClO4-H2SO4消煮,钼锑抗比色法测定;土壤速效磷(available phosphorus,AP)采用NaHCO3浸提-锑抗比色法测定;土壤速效钾(available potassium,AK)采用NH4OAc 浸提-火焰光度法测定。
土壤AMF 群落多样性测定,由上海美吉生物医药科技有限公司完成。采用AMV4-5NF/AMDGR引物(Sato et al.,2005),序列为:前引物5′-AAGCTCGTAGTTGAATTTCG-3′,后引物 5′-CCCAACTATCCCTATTAATCAT-3′,PCR 扩增在ABI GeneAmp® 9700 扩增仪上进行;扩增体系:5×反应缓冲液4 μL,dNTP(2.5 mmol·L−1),前、后引物(5 μmol·L−1)各0.8 μL,DNA 聚合酶0.4 μL,BSA 0.2 μL,DNA 模板10 ng,超纯水(ddH2O)补足至20 μL。扩增程序:95 ℃预变性3 min,95 ℃变性30 s,55 ℃退火30 s,72 ℃延伸45 s,共25个循环;72 ℃延伸10 min,10 ℃保存。采用Illumina MiSeq 平台对AMF 群落DNA 片段进行双端测序,按97%的序列相似度进行OTU 划分;以RDP classifier 贝叶斯算法对97%相似水平的OTU 代表序列进行分类学分析,通过maarjam20220506/AM物种数据库进行对比,获取分类学信息。
1.4 数据分析
采用Excel 2019 进行数据整理,以SPSS 26 进行单因素方差分析(one-way ANOVA)与邓肯检验(Duncan’s test),分析不同植被类型间土壤理化性质的差异性(P<0.05),单尾显著性检验不同变量间的皮尔逊相关性。土壤AMF 群落α 多样性、β 多样性利用公司运算平台分析,采用非度量多维尺度分析( non-metric multidimensional scaling analysis,NMDS)和相似度分析检验(ANOSIM),分析AMF群落间的差异,基于999 个排列的ANOSIM 计算与NMDS 相关的r值和P值;以Canoco 5 进行冗余分析(redundancy analysis,RDA);以Origin 2021进行绘图。图表中数据为平均值±标准差。
2 结果与分析
2.1 蒙古扁桃灌丛根际土壤理化性质
G×S−G 和G×S−S 的根际土壤pH、有机质、全氮、速效磷和速效钾含量最大,根际土壤碱解氮和全磷含量仅次于G,两者间除pH 有显著性差异外(P<0.05),其余均无显著性差异,说明共生下两种植物根际土壤理化性质没有差异。S 下的土壤养分含量最小。有机质在G×S−G 与S 间呈显著性差异(P<0.05);碱解氮在G 与其他3 种处理间,S 与G×S−G 和G×S−S 间呈显著性差异(P<0.05);全磷在G 和S 间呈显著性差异(P<0.05);速效磷在S与其他3 种处理间呈显著性差异(P<0.05);速效钾在G 与S 和G×S−G 间,S 与G×S−G 和G×S−S 间呈显著性差异(P<0.05)(表2)。
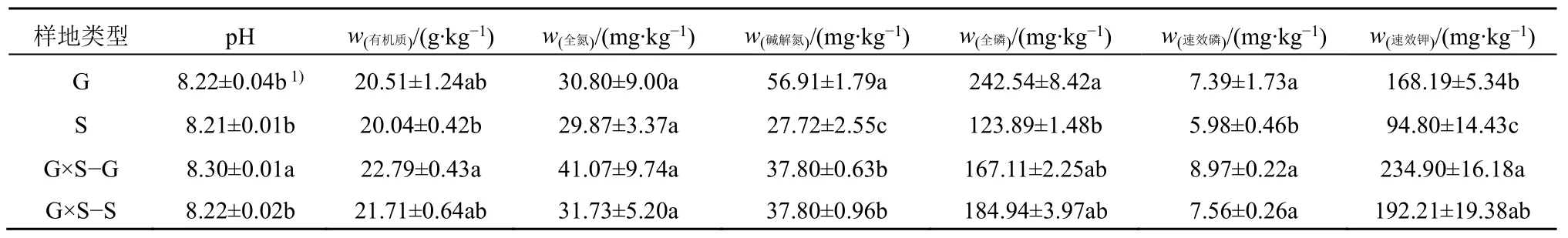
表2 根际土壤基本理化性质Table 2 Rhizosphere soil physical and chemical properties
2.2 蒙古扁桃灌丛根际土壤AMF 群落α 多样性
4 种处理下共获得OTU174 个(图1),其中G×S−G 有154 个,G×S−S 有152 个,G 有139 个,S 有143 个,4 种处理下共有OTU 数目115 个,G、G×S−G 和S 共有1 个,G、G×S−G 和G×S−S 共有14 个,G、S 和G×S−S 共有2 个,G×S−G、S 和G×S−S 共有8 个,G 和G×S−G 共有1 个,G 和S共有2 个,G 和G×S−S 共有1,G×S−G 和S 共有5 个,G×S−G 和G×S−S 共有8 个,S 和G×S−S 共有2 个,而G×S−G、G×S-S、G 和S 各自特有OTU分别为2、2、3 和8 个。
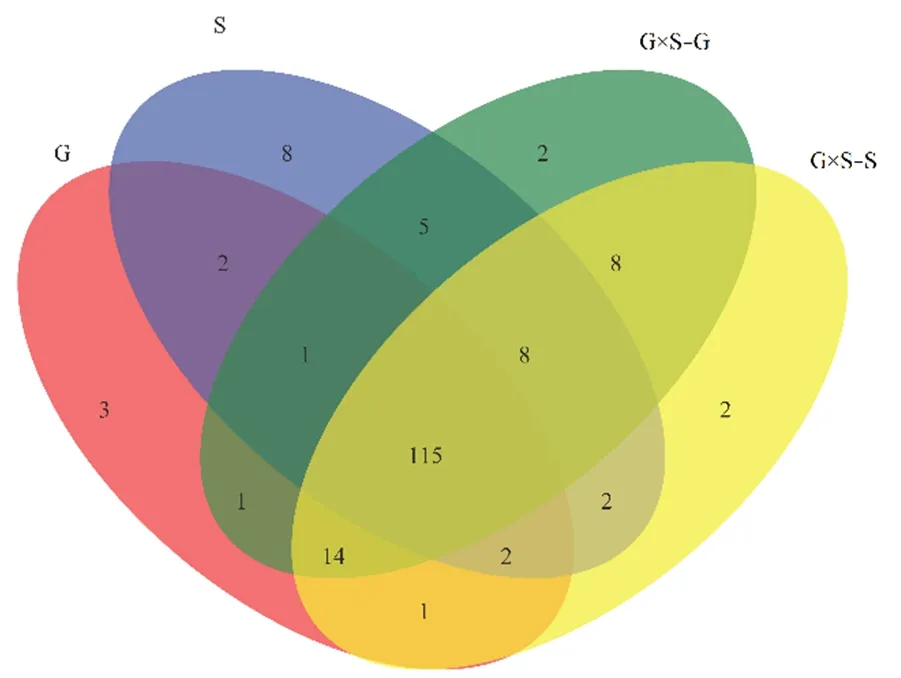
图1 4 种处理下根际土壤AMF OTU 分布维恩图Figure 1 Venn diagram of rhizosphere soil AMF OTU distribution under four treatments
4 种处理下根际土壤AMF 群落的物种多样性和丰富度产生了不同影响(图2)。G×S−G 和G×S−S下AMF 群落的Shannon 指数、Simpson 指数最大,两者无显著性差异,ACE 指数和Chao1 指数G×S−G最大,G×S−S 次之,S 最小。Shannon 指数在S 与其他3 种处理间呈显著性差异(P<0.05);Simpson指数在S 和G×S−G 间呈显著性差异(P<0.05);ACE 指数和Chao1 指数在G×S−G 和其他3 种处理间呈显著性差异(P<0.05),ACE 指数在S 和G×S−S间,Chao1 指数在G、S 与G×S−S 间呈显著性差异(P<0.05)。
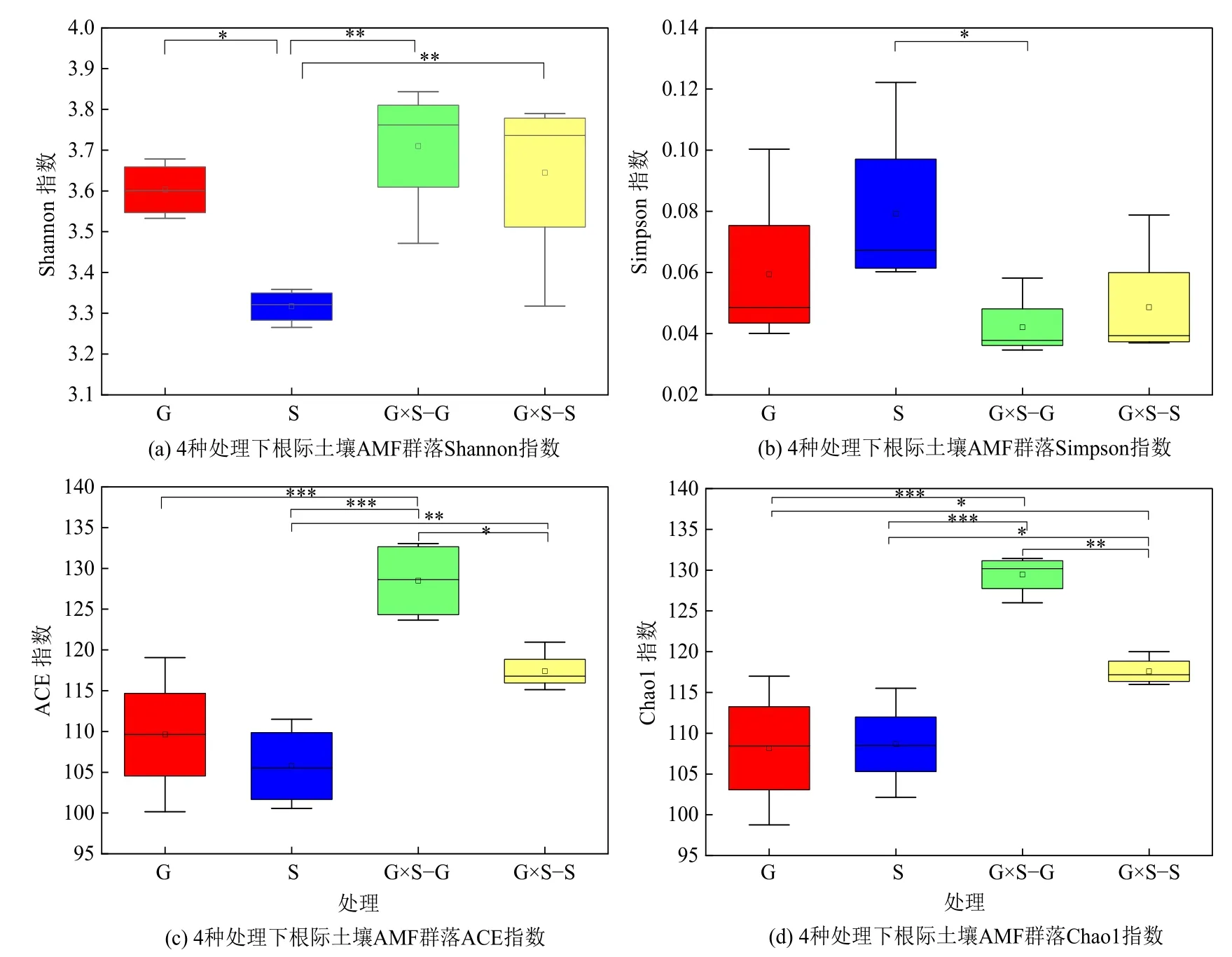
图2 4 种处理下根际土壤AMF 群落α 多样性指数箱型图Figure 2 Box plot of AMF communities α diversity index of rhizosphere soil under four treatments
2.3 蒙古扁桃灌丛根际土壤AMF 分类学组成和分布
通过maarjam20220506/AM 物种数据库进行对比,4 种处理下AMF 隶属于1 门4 纲6 目10 科10属63 种,分别为球囊霉属(Glomus)、类球囊霉属(Paraglomus)、原囊霉目待定属(unclassified_o_Archaeosporales)、原囊霉属(Archaeospora)、两性球囊霉属(Ambispora)、近明球囊霉属(Claroideoglomus)、球囊霉目待定属(unclassified_o_Glomerales)、球囊菌门待定属(unclassified_p_Glomeromycota)、多孢囊霉属(Diversispora)、球囊菌纲待定属(unclassified_c_Glomeromycetes)。在属水平上计算各处理不同属的比例,G×S−G、G×S−S、G 和S4 种处理下球囊霉属占比最大,分别为99.36%、98.97%、99.36%和99.36%,平均占比为99.26%,其余9 属总计占比分别为0.64%、1.03%、0.64%和0.64%,占比由大到小分别为近明球囊霉属、多孢囊霉属、球囊菌纲待定属、球囊菌门待定属、球囊霉目待定属、原囊霉属、类球囊霉属、两性球囊霉属和原囊霉目待定属(图3)。通过物种差异分析表明,仅在S 下发现两性球囊霉属,G×S−G下发现原囊霉目待定属,且仅两性球囊霉属在4 种处理间呈显著性差异(P<0.05)(图4)。
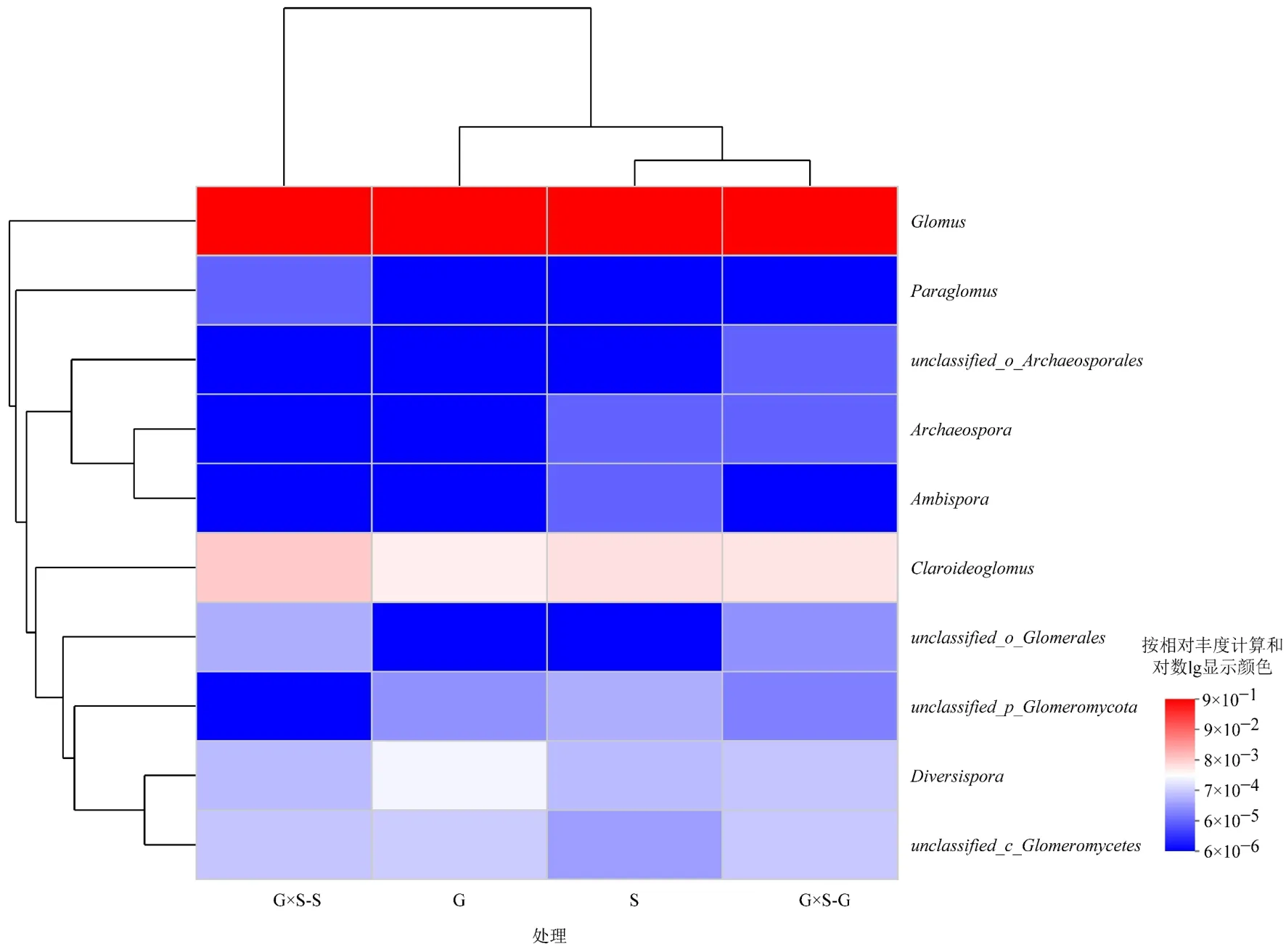
图3 4 种处理下AMF 属水平分类学组成和分布Figure 3 Taxonomic composition and distribution of AMF genera at the level of four treatments
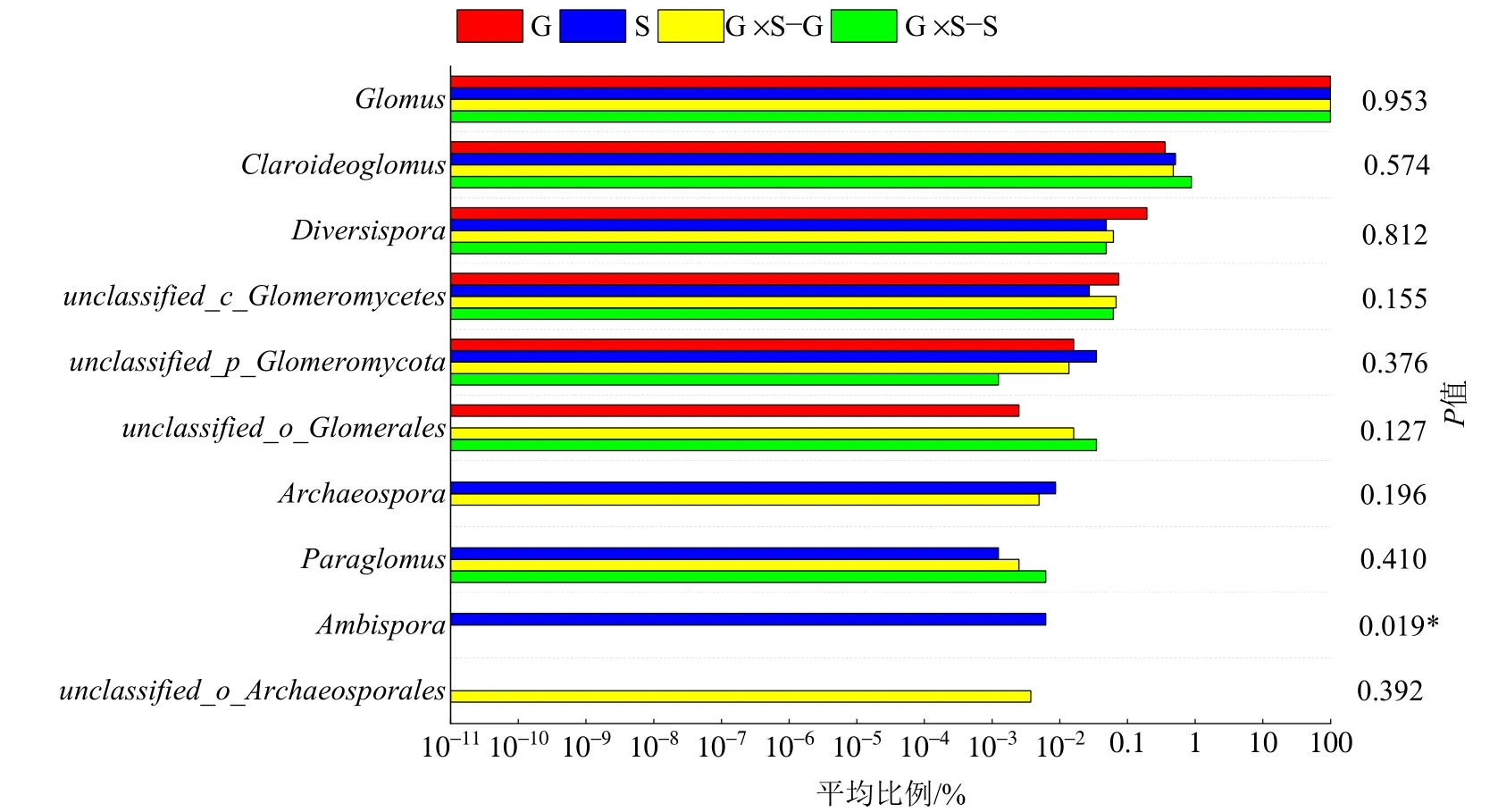
图4 4 种处理下AMF 物种差异显著性检验Figure 4 Significance test for differences in AMF species under the four treatments
基于OTU 水平和bray_curtis 距离计算的根际土壤 AMF 群落结构的非度量多维尺度分析(NMDS)结果表明(图5),S 组内相对分散,说明样本组内重复间相似性较差,其余组内相对较为集中,说明样本组内重复间相似性较好;4 组样本组间重合区域较大,说明物种组成相似性较高。对999个排列组合的相似性(ANOSIM)分析发现(stress=0.14,r=0.15,P=0.02),4 种处理下根际土壤AMF 群落多样性在OTU 水平下具有差异性。
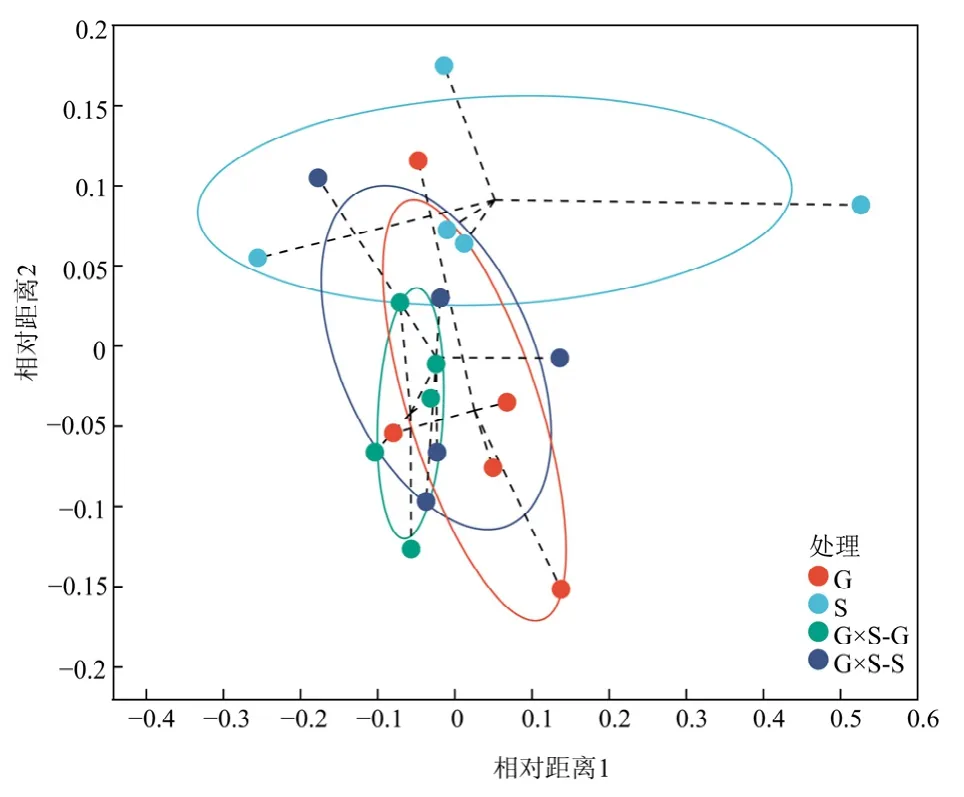
图5 4 种处理下土壤AMF β 多样性的非度量多维尺度分析Figure 5 Non-metric multidimensional scaling analysis(NMDS) of the soil AMF β diversity under four treatments
2.4 蒙古扁桃灌丛根际土壤AMF 多样性、属水平分布与土壤理化性质间的相关分析
将占比小于 0.01%的物种合并为其它属(Others),通过Person 相关性分析得到如下结果(表3),土壤pH 与Shannon 指数呈显著正相关,与Simpson 指数呈显著负相关;土壤有机质与ACE指数、Chao1 指数和其他属呈显著正相关;土壤全氮与Shannon 指数呈极显著负相关,与Simpson 指数呈极显著正相关;土壤碱解氮与Shannon 指数呈显著正相关,与Simpson 指数呈显著负相关;土壤全磷与球囊霉属呈显著正相关,与近明球囊霉属呈显著负相关;土壤速效磷与Shannon 指数、ACE 指数和Chao1 指数呈显著正相关,与球囊霉属呈显著负相关;土壤速效钾与Shannon 指数呈极显著正相关,与ACE 指数、Chao1 指数和近明球囊霉属呈显著正相关,与Simpson 指数和球囊霉属呈极显著负相关。

表3 AMF 群落多样性、属水平相对丰度与土壤环境因子间的皮尔逊相关性分析Table 3 Pearson’s correlation analysis of AMF community diversity, relative abundance based on the genus level and soil physicochemical properties
Shannon 指数、ACE 指数、Chao1 指数、近明球囊霉属和其他属与土壤速效钾、有机质和速效磷呈正相关;土壤碱解氮、pH 与近明球囊霉属和Shannon 指数、ACE 指数和Chao1 指数呈正相关;土壤全氮与Simpson 指数、球囊霉属和其他属呈正相关;土壤全磷与球囊霉属呈正相关。其中速效钾(解释量19.4%,P=0.006)对AMF 群落α 多样性和物种丰度影响显著。第一排序轴解释了AMF 群落组成24.77%的变异,第二排序轴解释了AMF 群落组成8.29%的变异。第一排序轴对蒙古扁桃灌丛根际土壤AMF 群落多样性及丰富度的变化解释最多(图6)。

图6 AMF 多样性与土壤理化性状间的冗余分析Figure 6 Redundancy analysis between AMF diversity and the soil physicochemical properties
3 讨论
3.1 蒙古扁桃灌丛根际土壤养分及AMF 群落结构特征
本研究表明,蒙古扁桃灌丛根际土壤养分含量大于蒙古扁桃和短花针茅单一植被类型的根际土壤养分含量且差异显著,这与邱江梅等(2020)的研究结果部分相同。由于灌丛的地上植物丰富,其根系和凋落物相对较多,相比单一植被能缓解降水对地表的冲刷,减少了土壤养分淋失;灌丛下土壤养分更为丰富,导致AMF 群落在灌丛下富集,符合灌丛“沃岛”效应,说明灌丛更有利于土壤AMF的生存,反映出灌丛对贺兰山低海拔地区灌丛地下生态系统的重要意义。
本试验共得到174 个丛枝菌根真菌OTUs,隶属于1 门、4 纲、6 目、10 科、10 属、63 种,球囊霉属是蒙古扁桃灌丛根际土壤AMF 的优势属,这与王占军等(2020)在宁夏荒漠草原、干草原和格根宝乐尔(2013)在内蒙古短花针茅荒漠草原地区得到球囊霉属是优势属的结论一致,也和研究发现蔷薇科灌木土壤下球囊霉属是优势属的结果一致(杨秀丽等,2010)。在高海拔地区土壤、沙漠、农田土壤等类型得到的研究结果也是球囊霉属为优势属(Qin et al.,2015),说明球囊霉属能与蒙古扁桃灌丛形成良好的共生关系,并且提高植物对土壤养分的吸收。本试验的两性球囊霉属仅在灌木分布且差异显著,说明更适合在低养分土壤生存;近明球囊霉属、多孢囊霉属、球囊菌纲待定属和球囊菌门待定属在4 种处理下均有分布且无显著性差异,说明其生态位较广;原囊霉属和类球囊霉属仅在灌木和灌丛下分布,说明更适合与灌丛共生。本研究说明了人为扰动对根际土壤AMF 群落的变化产生了一定影响,但对球囊霉属等物种的影响较小;但本研究中未检测出无梗囊霉属(Acaulospora)物种,与王琚钢等(2011)在内蒙古西部的蒙古扁桃样地调查的研究结果不一致,可能是地域差异导致土壤因素不同,造成了研究结果的差异。因为土壤AMF的球囊霉属和无梗囊霉属是出现频率较高的AMF属,其包括的AMF 物种数较多,扩散、定殖能力较强以及适应性较广(Menezes et al.,2016)。本试验检测出了球囊霉属且相对丰度均占98%以上,分析认为,由于贺兰山处于干旱半干旱地区,导致低海拔地区土壤发育差,养分在有限的情况下,微生物间存在竞争关系,球囊霉属物种能利用自身的产孢效率快速占据土壤环境(Xue et al.,2015),与寄主植物根系能更快形成菌根,建立起互利共生关系,提高了寄主植物从土壤中吸收养分的能力;寄主植物又促进了对球囊霉属的能量分配,从而导致土壤AMF 的其他物种的相对丰度降低。采用的AMV4-5NF/AMDGR 这一对扩增引物对球囊霉科AMF 物种有扩增偏好的特点,从而使球囊霉属的AMF 比其他科、属更容易被扩增出来而被检测到(Van Geel et al.,2014)。
3.2 蒙古扁桃灌丛根际土壤AMF 群落结构影响因素
土壤AMF 群落的组成、分布及多样性受土壤因素(土壤类型、理化性质),植被类型的影响(胡嘉丽,2019),岩性和坡向也可改变土壤性质,从而对土壤AMF 群落产生影响(柴宇星,2018;张晓晓等,2019);随着植物多样性的增加,土壤AMF群落多样性也呈升高趋势(Mellado-Vazquez et al.,2016)。但有些研究表明,驱动植物和AMF 群落变化的原因是非生物环境因子(Zobel et al.,2014)。本试验所选样地处于同一海拔梯度,空间位置相对集中,气候条件、土壤因素、岩性、坡向相对一致,所以相对非生物条件而言,不同的植被类型可能是影响根际土壤AMF群落结构和多样性的主要原因。通过对根际土壤理化性质和AMF 群落α 多样性及物种相对丰度进行相关性分析表明,土壤速效钾与除了其他属外均为显著性相关,其他理化性质也和AMF 群落呈一定显著相关性,与前人的研究结果基本一致(盛敏等,2011;任爱天等,2014),说明土壤速效钾是贺兰山低海拔地区蒙古扁桃灌丛根际土壤AMF 群落α 多样性和物种相对丰度的主要影响因子,土壤氮、磷是次要影响因子。土壤pH 不同,AMF 组成可能也不同,pH 能直接影响菌根的形成,也影响AMF 的产孢率,其中球囊霉属有着较宽的pH 耐受幅度,在pH 5-9 的土壤中出现较多,本试验区土壤pH 范围在8.21-8.30 之间,所以球囊霉属相对丰度占比较高,但未发现无梗囊霉属物种,这也验证了无梗囊霉属喜好pH 小于7 的偏酸性土壤的特性。土壤的高有机碳含量有利于促进AMF 的生长,土壤有机质与丰富度指数和其他属相对丰度呈显著正相关,但土壤速效磷含量过高往往会抑制AMF 的生长、发育和功能。本试验样地土壤速效磷含量最高为8.97 mg·kg−1,为低磷土壤环境,适合AMF 群落生长,所以AMF 群落α 多样性指数与土壤速效磷呈显著正相关,但球囊霉属与其呈显著负相关,可能因为球囊霉属相对丰度占比过高。球囊霉属和近明球囊霉属相对丰度与土壤全磷呈显著相关性,这与马琨等(2019)的研究结果不一致,可能因为其农田土壤全磷含量为0.83 g·kg−1,远高于本试验区的土壤全磷含量,抑制了AMF 的生长。土壤全氮和碱解氮与AMF 多样性指数呈显著相关,这与周光荣(2020)在梵净山的研究结果不一致,与王占军等(2020)在宁夏草原的研究结果较为一致,可能因为研究的地域差异导致环境因素不同而导致结果不同。
本研究RDA 表明,土壤速效钾和AMF 群落α多样性指数和属相对丰度呈极显著相关(P<0.01),这和赵蕾等(2019)在喀斯特地区的研究结果一致,由此认为土壤速效钾是影响土壤AMF 群落结构和多样性的重要因素。后续研究可通过对土壤AMF孢子、AMF 分泌的球囊霉素相关蛋白和根系侵染率以及地上微环境进行检测,进一步论证蒙古扁桃灌丛根际土壤AMF群落特征和外界因素之间的关系。
4 结论
(1)宁夏贺兰山国家级自然保护区低海拔地区蒙古扁桃灌丛下根际土壤AMF 隶属于1 门、4 纲、6 目、10 科、10 属、63 种,优势属为球囊霉属。
(2)蒙古扁桃灌丛两种植物根际土壤AMF 群落α 多样性和根际土壤理化性质高于蒙古扁桃和短花针茅单一植被,符合灌丛“沃岛”效应。
(3)AMF 群落结构及α 多样性受根际土壤理化性质和生物因素的显著影响,其中土壤速效钾是主要影响因子,氮磷次之。
猜你喜欢
水土保持学报(2022年5期)2022-10-10
西北农林科技大学学报(自然科学版)(2022年3期)2022-04-01
中国土壤与肥料(2021年5期)2021-12-02
草业科学(2019年5期)2019-06-04
中成药(2018年6期)2018-07-11
天然产物研究与开发(2018年3期)2018-05-07
中国蔬菜(2016年8期)2017-01-15
天津师范大学学报(自然科学版)(2016年4期)2016-12-14
甘肃林业(2016年3期)2016-11-07
国外医药(抗生素分册)(2016年2期)2016-07-12
