
CO2浓度升高下水稻株高、茎蘖与SPAD 动态响应及其模拟
2023-08-10 06:15杨凯杨靖睿曹培培吕春华孙文娟于凌飞邓希
生态环境学报 2023年5期
杨凯,杨靖睿,曹培培,吕春华,孙文娟,于凌飞,邓希
1.中国科学院植物研究所/植被与环境变化国家重点实验室,北京 100093;2.中国科学院大学,北京 100049;3.中山大学大气科学学院,广东珠海 519000
自工业革命以来,大气CO2浓度迅速升高,到2021 年,全球平均CO2浓度已达到414 μmol·mol−1,约为工业革命前CO2水平的1.5 倍(https://www.esrl.noaa.gov/gmd/ccgg/trends/global.html)。中等典型浓度路径情景下(RCP4.5 与RCP6.0),全球CO2浓度在本世纪末将达到538-670 μmol·mol−1(IPCC,2022)。中国是一个农业大国,水稻产量占全国粮食总产量的32%(http://www.stats.gov.cn/)。65%的中国人以大米为主食,预计到2030 年中国的水稻需求量将增加20%(Peng et al.,2009)。气候变化对水稻生产具有直接的影响,研究表明:温度升高可造成水稻减产,但CO2施肥效应可能对该损失具有补偿作用(Lobell et al.,2011;黄耀,2017;方精云等,2018),从而在一定程度上缓解气候变暖对粮食安全造成的威胁。
株高作为水稻直观的形态指标,与产量密切相关(严定春等,2008);茎蘖的增长与消亡直接影响水稻的成穗率,进而决定产量构成中的主要因素——有效穗数(王萌萌等,2016)。CO2浓度升高条件下,CO2的施肥效应主要作用在生育前期,导致水稻在生育前期消耗过多的氮,引起生育后期氮的供给不足,用于光合作用的叶绿素含量降低,从而使生殖生长阶段获得较少的碳吸收(Sakai et al.,2001;Sakai et al.,2006;Wu et al.,2018)。因此,用于表征叶绿素相对含量的SPAD 可作为生育后期功能叶片衰老与氮素营养的参考指标(边立丽等,2022)。大气CO2浓度升高一般对农作物的株高与分蘖具有正向效应(Kim et al.,2003;Hu et al.,2022;黄建晔等,2002;杨连新等,2007),但对叶绿素含量的影响仍无定论(蒋跃林等,2005;Koti et al.,2007;周宁等,2017)。值得注意的是,上述研究大多集中在某一生育期或时间点上,但在CO2浓度升高下,对株高、分蘖与SPAD 动态变化的研究尚不足。
通过田间试验数据建立作物生长模型,是研究气候变化情景下作物生长特征的重要方法(Tubiello et al.,2007)。生育期是田间试验观测的重要指标,它影响着作物模型在不同生长发育阶段的参数化。水稻是喜高温的短日照作物,完成某一阶段生长发育所需要的有效积温相对稳定。因此,以有效积温(growing degree day,GDD)建立作物生长模型能有效表征作物生长发育随时间和温度变化的特点(Lecoeur et al.,1998;苏李君等,2020;陈杨等,2022)。常规条件下,前人已开展了作物株高、分蘖与SPAD 生长动态模拟的研究(黄耀等,1994;王声锋等,2010;杨沈斌等,2016;赵考诚等,2021),但在CO2升高条件下,基于连续观测的模型模拟还较为少见(Grant et al.,2001;孙成明,2006a;Tubiello et al.,2002)。
本文通过两年试验探究水稻株高、分蘖与SPAD 生长动态对CO2升高的响应,并利用模型模拟的方法对动态特征进行定量描述,从而为未来CO2升高条件下,水稻生长发育和产量形成的预测提供科学依据,这也对未来水稻的栽培、管理与生产具有一定的理论和实践意义。
1 材料与方法
1.1 试验地概况
试验区域位于江苏省南京市南京信息工程大学农业气象生态实验站(32.21°N,118.71°E)。该区域属于亚热带湿润气候,年平均气温15.6 ℃,年平均降水量约1 100 mm,平均日照时数超过1 900 h,无霜期为237 d。供试土壤为潴育型水稻土,灰马肝土属,土壤质地为粉砂壤土,0-20 cm 耕层土壤中砂土含量为9.5%,粉土含量为85.2%,pH 为6.1,容重1.51 g·cm−3,土壤有机碳9.52 g·kg−1,全氮1.18 g·kg−1,全磷0.85 g·kg−1,全钾18.17 g·kg−1。
试验开展于2017 与2018 年的水稻生长季。试验样地设有原位自动小型气象站(AWS800,Campbell Scientific,Inc.,USA)用以监测并记录试验期间的气象数据。
1.2 试验设计
CO2控制试验平台由应用较为广泛的开顶式气室(Open-Top Chambers,OTC)和CO2浓度控制系统组成。OTC 外形呈正八边形棱柱状,对边直径为3.75 m,高3 m,底面积约为12 m2,主体采用铝合金框架,室壁材料为高透光性玻璃,为了使气体散失速度放缓,顶部开口向内倾斜45o。关于CO2浓度控制系统的细节,详见刘晓萌等(2018)和Yang et al.(2021)。
中等典型浓度路径情景(RCP4.5 与RCP6.0)预测全球CO2浓度在本世纪末将达到538-670 μmol·mol−1(IPCC,2022)。因此,利用CO2控制试验平台进行CO2浓度模拟,共设置两个CO2浓度处理:一个是背景大气CO2浓度(a[CO2],大约400µmol·mol−1),另一个CO2浓度是在a[CO2]的基础上增加200 µmol·mol−1(e[CO2])。a[CO2]与e[CO2]各有4 个OTC 作为重复。
1.3 试验材料
供试水稻品种为常规粳稻“南粳9108”,全生育期为149-153 d。在2017 年,水稻种植在OTC内的土壤;在2018 年,水稻种植在OTC 内的盆栽土壤(OTC 内土壤与盆栽土壤相同,盆钵上口外径18 cm×18 cm,下口外径14.5 cm×14.5 cm,高18 cm)。两年均在5 月20 日播种,6 月20 日移栽,移栽密度为每穴3 株主茎苗。全生育期施氮量约为20 g·m−2,分3 个时期施用,各时期氮肥施用比例为基肥꞉分蘖肥꞉穗肥=4꞉3꞉3。基肥采用复合肥(15% N:15% P2O5:15% K2O),分蘖肥和穗肥均为尿素(含氮量为46.7%)。水分管理和虫害治理等田间管理措施统一按当地常规栽培要求执行。两年水稻主要生育时期见表1。

表1 水稻主要生育时期Table 1 Rice calendars
1.4 测定项目与方法
水稻株高的测定为量取植株地表至最高叶顶(抽穗前)或最高稻穗顶(抽穗后);测定时间为移栽缓苗至抽穗后1 周(抽穗后1 周的株高认定为最终株高),每隔5 天测定一次。茎蘖数的测定为手动观测且小于3 张叶片的茎蘖不予计入;测定时间为移栽缓苗后至抽穗期,每隔5 天测定一次。使用SPAD-502 plus 型叶绿素仪(SPAD-502,Konica Minolta Optics,Inc.,Japan)测定旗叶、倒二叶与倒三叶的SPAD,测定位置为叶片中间的1/2 部位,且以8 个点测定的平均值代表该叶片的SPAD 值;测定时间为抽穗至成熟期,每隔5-7 天测定一次。2017 年与2018 年均测定了株高动态与SPAD 动态,而茎蘖动态的测定仅在2018 年进行。此外,本研究还测定了2016 年(OTC 土壤)、2019 年(OTC 盆栽土壤)与2020 年(OTC 土壤)抽穗1 周后(9 月1 日左右)的株高。
1.5 数据处理
应用Microsoft Excel 2016 进行数据处理;SPSS 21.0(IBM Crop.Armonk,NY,USA)进行单因素(CO2)与双因素(CO2与叶位)方差分析;使用R统计软件(4.1.3,http://www.r-project.org/)进行模型拟合;Origin 2017 软件进行绘图。
2 结果与分析
2.1 CO2浓度升高下株高动态及其模拟
无论背景CO2还是CO2浓度升高下,水稻株高总体上随GDD 呈现先迅速增加,后缓慢增长,再逐渐趋于平缓的特征,其动态变化曲线为“S”型(图1)。CO2浓度升高对两年水稻抽穗前的株高均无影响,但2017 年与2018 年的最终株高对CO2的响应不同:2017 年,CO2浓度升高显著增加了最终株高,其增幅为7.1%;但是,2018 年并未观测到CO2促进效应。因此,在2017 年和2018 年分别建立株高动态模型(孙成明等,2006b)。结合其他年份(2016、2019 和2020 年)最终株高对CO2升高的响应,通过整合分析发现:CO2升高对最终株高的促进效应与抽穗1 周后(9 月1 日左右)的有效积温有关,即,当抽穗1 周后的有效积温大于720 ℃·d 时,CO2浓度升高对最终株高具有促进作用,反之,则无促进效应(图2)。

图1 2017 年(a)与2018(b)年CO2 浓度升高下株高随有效积温(GDD)的动态变化Figure 1 Dynamics of plant height with growing degree day (GDD) under e[CO2] in 2017 (a) and 2018 (b)

图2 CO2浓度升高下株高相对变化量与抽穗1 周后有效积温(GDD)的关系Figure 2 The relationship between relative change of plant height and growing degree day (GDD) of one week after heading under e[CO2]
因此,本研究将GDD 作为自变量,采用Logistic方程对株高动态进行拟合。
式中:
TGDD——移栽后的有效积温(℃·d);
Ti——移栽后第i天的日平均温度;
10——水稻生长的生物学下限温度(℃)(高亮之等,1992)。
a[CO2]下的株高动态模型:
式中:
H——a[CO2]下理论最大株高;
a1、a2——模型系数。
根据Ziska et al.(2007)提出的CO2响应方程:
式中:
β——CO2响应比;
Ya与Ye——a[CO2]与e[CO2]的测定值;
Ca与Ce——a[CO2]与e[CO2]的CO2浓度,分别取400、600 μmol·mol−1。
则,e[CO2]下的株高动态模型:
式中:
β——株高促进的CO2响应比,根据2017 年实测数据取0.139。
株高动态模型模拟的结果见表2。该模型可准确模拟出株高对CO2升高响应的年际性差异。两年的均方根误差(RMSE)分别为2.96 cm 与3.52 cm,表明模拟值与观测值之间的误差较小。此外,两年模型的r2分别为0.970 与0.953,表明模型的模拟结果较好。图3a 显示了两年株高模拟值与观测值的比较结果,其散点基本均落在1꞉1 线附近,说明模拟值与观测值间具有很好的一致性。
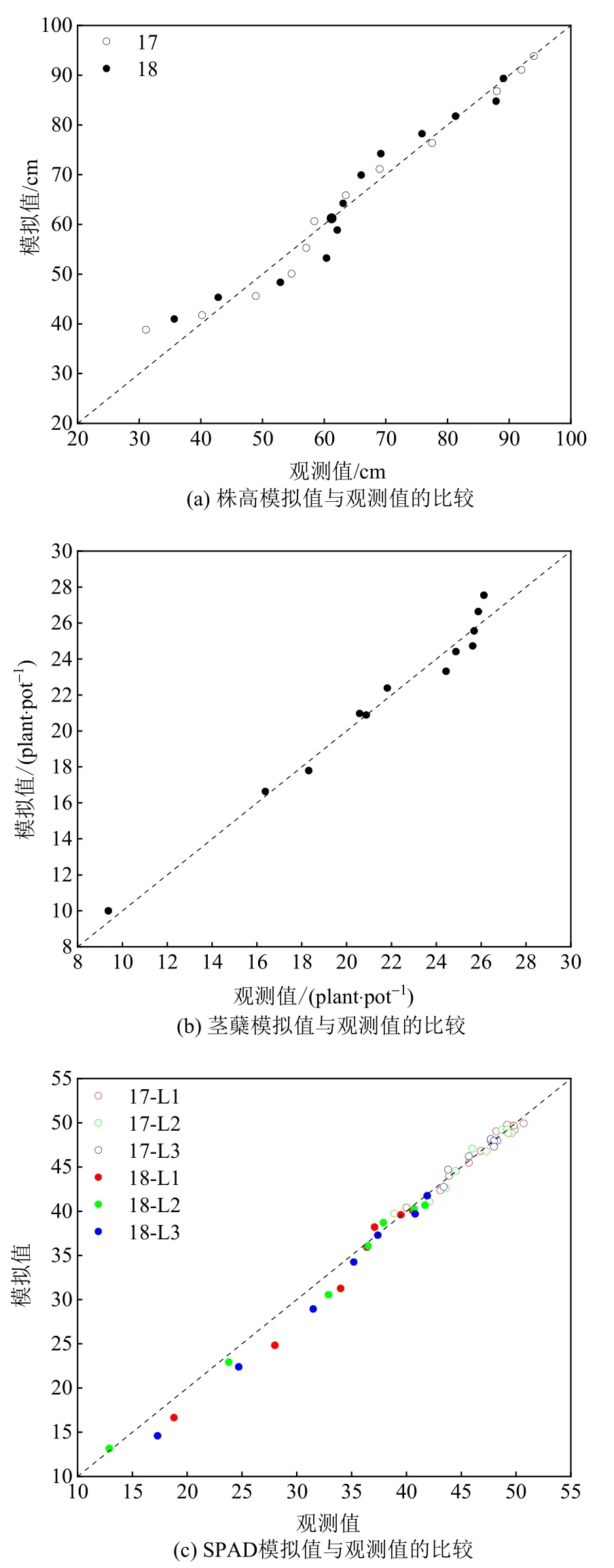
图3 株高(a)、分蘖(b)与SPAD(c)动态观测值与模拟值的比较Figure 3 Comparison of observations and simulations in plant height (a), tiller number (b) and SPAD(c) dynamics

表2 株高动态模型参数值Table 2 Parameters of dynamic model of plant height
2.2 CO2浓度升高下茎蘖动态及其模拟
无论背景CO2还是CO2浓度升高下,水稻群体茎蘖呈现先增长后消亡的动态变化特征(图4)。茎蘖增长阶段:自移栽缓苗后分蘖开始增加,至拔节期前后(大约移栽后35 d 左右)达到最高苗,总体上为正“S”型曲线;茎蘖消亡阶段:在最高苗出现以后,一部分茎蘖生长减缓,成为无效分蘖,在之后的生长过程中逐渐死亡,总体上为反“S”型曲线。水稻的分蘖数在CO2浓度升高下总体上呈现增加的趋势,但是CO2促进效应不明显,尤其是在移栽后10 d 以内(图4)。

图4 CO2浓度升高下茎蘖数随移栽时间的动态变化Figure 4 Dynamics of tiller number with transplanting time under e[CO2]
根据水稻茎蘖的动态变化特征,本研究采用Logistic 修正方程与倒Logistic 修正方程分别对茎蘖增长与消亡的动态过程进行模拟:
a[CO2]下的茎蘖增长模型:
式中:
T1——a[CO2]下最大分蘖数;
a1、a2、a3——模型系数;
t——移栽后天数。
a[CO2]下的茎蘖消亡模型:
式中:
T2——a[CO2]下最大死亡分蘖数;
b1、b2、b3——模型系数;
t——移栽后天数。
根据方程(3),则e[CO2]下茎蘖消长动态模型表示为:
式中:
β1与β2——茎蘖增长与消亡的CO2响应比,根据2018 年实测数据分别取0.174 与0.167。
茎蘖增长与消亡的动态拟合结果见表3。从模拟结果可知,常规条件下水稻每盆理论茎蘖最大存活数与最大死亡数分别为27 株与10 株。无论是茎蘖增长还是茎蘖消亡模型,均具有较高的拟合度,其r2分别为0.971 与0.996。茎蘖增长与消亡模型的RMSE 分别为0.77 株/盆与0.65 株/盆,说明拟合误差值较小。此外,从模拟值与观测值的1꞉1 关系图可知(图3b),所有点均落在该线附近,茎蘖增长与消亡模型具有较高的模拟精度。

表3 茎蘖消长动态模型参数值Table 3 Parameters of dynamic model of tiller growth and extinction
2.3 CO2浓度升高下不同叶位SPAD 动态及其模拟
水稻抽穗后旗叶、倒二叶与倒三叶的SPAD 均随抽穗后天数呈逐渐降低的动态变化(图5)。总体上,SPAD 的大小为:旗叶>倒二叶>倒三叶,这种趋势差异在2017 年更为明显(图5,表4)。CO2浓度升高对2017 年(OTC 内土壤)与2018 年(OTC 内盆栽土壤)旗叶与倒二叶SPAD 的影响不同:在2017年,CO2浓度升高增加了抽穗后35 d 旗叶与倒二叶的SPAD,并且随生育期的进程,其CO2促进效应逐渐增加;但是,2018 年旗叶与倒二叶的SPAD 对CO2升高几乎无响应(表4)。此外,CO2浓度升高对两年内倒三叶的SPAD 几乎均无影响(表4)。

图5 2017 年(a)与2018 年(b)CO2浓度升高下抽穗后SPAD 的动态变化Figure 5 Dynamics of SPAD with days after heading under e[CO2] in 2017 (a) and 2018 (b)

表4 不同测定日期下CO2浓度升高与叶位对SPAD 影响Table 4 Effects of e[CO2] and leaf position on SPAD at different measured time
由于SPAD 动态在2017 年和2018 年差异显著,本研究采用多项式回归方程,分年份对水稻不同叶位SPAD 动态特征进行模拟:
a[CO2]下不同叶位SPAD 动态模型:
式中:
t——抽穗后天数;
a1、a2、a3——模型系数。
根据方程(3),2017 年(OTC 内土壤)e[CO2]下不同叶位SPAD 动态模型:
旗叶与倒二叶:
式中:
β1与β2——抽穗后0-35 d 与抽穗后35 d 之后的 CO2响应比,旗叶的取值分别为 0.070 与0.000 4×(t−35)2+0.003 9×(t−35)+0.085 5,倒二叶的取值分别为 0 与 0.001 2×(t−35)2−0.010 8×(t−35)+0.104 8。
倒三叶:
2018 年(OTC 内盆栽土壤),e[CO2]下不同叶位SPAD 动态模型与方程(8)相同。
水稻抽穗后不同叶位SPAD 的动态拟合结果见表5。从两年的拟合结果可知,所有叶位SPAD 的拟合模型在年际间(不同生长状态)具有较大差异。总体上,旗叶与倒二叶的拟合模型相似,但两者与倒三叶的拟合模型差异较大。两年不同叶位SPAD模型的r2在0.960-0.985 之间,表明其模拟效果较好。图3c 显示了两年所有叶位SPAD 模拟值与观测值的比较结果:尽管两年所有的点均落在1꞉1 线附近,但2018 年在抽穗后35 d 内的散点大多落于1꞉1 线之下,并且从RMSE 的结果也可看出,2017年的拟合精度略优于2018 年。
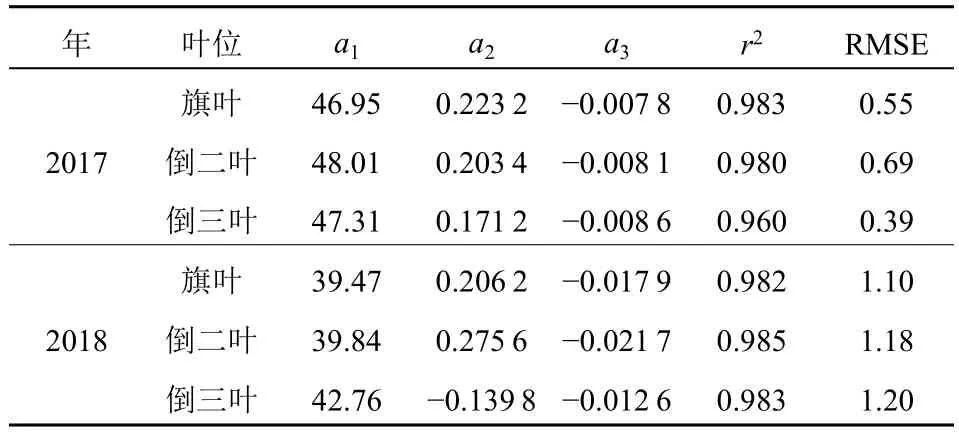
表5 SPAD 动态模型参数值Table 5 Parameters of SPAD dynamic model
3 讨论
3.1 CO2浓度升高下株高动态响应及其模拟
常规与CO2浓度升高条件下,株高随有效积温的动态变化均为“S”型曲线(图1)。无论是温室、开顶箱还是FACE 试验,CO2浓度升高一般对农作物的株高具有促进作用,增幅为4%-10.6%(黄建晔等,2002;杨连新等,2007;Liu et al.,2008;周宁,2020),且随生育进程逐渐增加,在抽穗期扬花期最显著(谢辉,2005;赖上坤,2011)。本文结果与前人研究基本一致:CO2浓度升高对抽穗之前株高无显著影响,但增加了抽穗后株高(最终株高增幅为7.1%)(图1)。进一步的整合分析发现,CO2浓度升高对株高的促进效应与有效积温有关(图2)。这表明生长温度调控水稻株高对CO2浓度升高的响应。水稻是喜高温作物,其茎鞘伸长的最适温度为28.5 ℃(Sanchez et al.,2014),且在CO2浓度升高条件下,水稻生理生长的最适温度将提升2.2-4.8 ℃(Lü et al.,2020)。因此,较高的有效积温有利于CO2浓度升高对株高的促进。此外,水稻的生长空间也可能影响CO2对株高的促进效应,但在2016 年与2020 年(OTC 内土壤),CO2浓度升高并未影响株高,因而生长空间并不是制约株高对CO2浓度升高响应的因素。
以大于10 ℃的有效积温与CO2响应比为驱动变量,采用Logistic 方程可有效模拟CO2浓度升高下株高的动态曲线(表2,图3a)。与大多数模型以生育期天数为驱动变量不同,本研究采用有效积温建立模型,并将有效积温进行阈值划分(720 ℃·d),这能够反映温度在CO2浓度升高下对水稻生长的影响。因此,当年际间气象环境条件存在差异时,也可有效建立CO2浓度升高下水稻的株高动态模型,具有较好的适用性。
3.2 CO2浓度升高下茎蘖动态响应及其模拟
CO2浓度升高条件下,水稻产量增加的主要原因是有效穗数的增加(Lü et al.,2020;Hu et al.,2021)。有效穗数的多少不但取决于水稻品种自身的分蘖能力、移栽密度和水肥管理,还与最大茎蘖密度等有着密切的联系(黄耀等,1994)。水稻茎蘖消长由产生的新生分蘖与死亡的无效分蘖共同体现,其增长阶段呈正“S”型,而消亡阶段呈反“S”型(图4)。以往的研究结果表明,CO2浓度升高提高了水稻的分蘖能力(Ziska et al.,1997;Kim et al.,2003;Hu et al.,2022;孙成明等,2006b)。本研究也发现CO2升高增加了分蘖数,但总体上这种促进效应并不明显,尤其是在移栽后10 d 内(图4)。
以移栽后天数与CO2响应比为驱动变量,采用Logistic 修正方程与倒Logistic 修正方程可有效模拟CO2升高下水稻茎蘖增长与消亡的动态(图3b,表3)。Yang et al.(2006)发现CO2浓度升高下茎蘖增长速率与消亡速率有差异,从而导致成熟期最终有效穗数与拔节期最大茎蘖数的比例不一致。因此,本研究在孙成明等(2006a)茎蘖动态模型的基础上,进一步区分了茎蘖增长与消亡阶段的CO2响应比。
3.3 CO2浓度升高下不同叶位SPAD 动态响应及其模拟
前人的研究表明,CO2升高对叶绿素含量的影响无一致性规律,或增加(蒋跃林等,2005;Zhu et al.,2009;谢立勇等,2015)、或不变(Koti et al.,2007)、或降低(Bindi et al.,2002;周宁等,2017)。本研究发现,SPAD 对CO2浓度升高的响应因叶位选择(旗叶、倒二叶vs倒三叶)与测定时间而异(表4)。总体上,CO2浓度升高增加了2017 年抽穗后35 d 旗叶与倒二叶的SPAD,但对其他时期或叶位的SPAD 无显著影响。此外,CO2浓度升高对SPAD 的影响在年际间差异较大。造成这种差异的原因可能是:相较于2017 年种植在大田的水稻(根系呈伞状发散形),2018 年种植在盆栽的水稻(根系围绕着土壤呈圆柱形),其根系生长空间较小,从而导致养分吸收受限。
以移栽后天数与CO2响应比为驱动变量,采用多项式回归模型可对CO2升高下抽穗后不同叶位SPAD 的动态曲线进行有效模拟(图3c,表5)。尽管两年不同叶位SPAD 模型的r2均大于0.96,但CO2浓度升高对SPAD 的影响因多种外界因素而异,其拟合模型可能具有局限性。
本研究主要基于两年的田间观测,观测时间较短、观测数据有限,模型并未进行不同年份的验证。但基于两年的结果,我们发现CO2效应因年际而异。田间试验和实际农业生产中,年际间气象因素的差异可能是造成年际间CO2效应差异的原因,这需要进一步开展长时间以及多因子的试验研究,建立CO2响应比与气象等环境因素的关系,并进行有效验证,从而使模型更具有普适性。
4 结论
水稻株高对CO2升高的响应由生长温度调控,可采用以大于10 ℃的有效积温和CO2响应比为驱动变量的Logistic 方程进行有效模拟。CO2升高总体上提高了水稻的分蘖能力,以移栽后天数和CO2响应比为驱动变量,可采用Logistic 修正方程与倒Logistic 修正方程有效模拟茎蘖增长与消亡动态曲线。SPAD 对CO2升高的响应因叶位选择与测定时间与而异,多项式回归模型能有效模拟抽穗后不同叶位SPAD 动态,其中抽穗后天数和CO2响应比是驱动变量。
致谢:南京信息工程大学刘超与商东耀对CO2控制平台维护与管理;北京天航华创科技股份有限公司对CO2浓度控制系统提供技术支持;江苏省农业科学院王才林研究员提供水稻种子,一并致谢。
猜你喜欢
农业与技术(2023年11期)2023-06-19
中国农业科学(2022年15期)2022-08-09
干旱地区农业研究(2022年4期)2022-07-28
作物学报(2022年9期)2022-07-18
中国马铃薯(2022年2期)2022-07-05
四川蚕业(2022年1期)2022-06-06
干旱地区农业研究(2021年6期)2021-12-08
今日农业(2021年17期)2021-11-26
四川农业科技(2019年10期)2020-01-06
黑龙江粮食(2019年10期)2019-11-23
