
新疆达坂城盐湖嗜盐细菌分离鉴定及活性分析
2023-07-28 13:32:24李怡歆刘晓禄艾尼江尔斯满徐李娟刘倩倩包晓玮宋素琴
新疆农业科学 2023年7期
李怡歆,陈 勇,刘晓禄,艾尼江·尔斯满,徐李娟,4,刘倩倩,包晓玮,宋素琴
(1.新疆农业科学院微生物应用研究所/新疆特殊环境微生物重点实验室,乌鲁木齐 830091;2.新疆农业大学食品科学与药学学院,乌鲁木齐 830052;3.新疆农业科学院土壤肥料与农业节水研究所,乌鲁木齐 830091;4.新疆农业大学农学院,乌鲁木齐 830052;5.新疆生产建设兵团第十三师农业科学研究所,新疆额敏 839000)
0 引 言
【研究意义】极端环境包括高压、高碱、高酸、高温、高盐和高辐射等[1]。极端环境的微生物因其生存的极端环境决定其生长繁殖以及代谢路径等[2]。且代谢产物能抵抗外界对自身的伤害[3]。盐湖因蕴含丰富的氯化钠,是一种咸水化水体。新疆是我国内陆盐类主要产地之一,例如罗布泊盐湖、艾比盐湖、巴里坤盐湖、达坂城盐湖和乌尔禾盐湖等。【前人研究进展】从达坂城盐湖分离到的微生物大多属于Firmicutes和γ-proteobacteria两大类群,包括盐单胞菌属(Halomonas)、枝芽孢杆菌属(Virgibacillus)、芽孢杆菌属(Bacillus)、喜盐芽孢杆菌属 (Halobacillus)、盐水球菌属 (Salinicoccus)、Ornithinibacillus、Oceanobacillus、Thalassobacillus和Salimicrobium[4];以及中度嗜盐菌Moderatelyhalophilicbacteria[5]、嗜盐古菌Natrialba、Haloarcula、Haloterrigena、Halorubrum等[6]。【本研究切入点】从普通环境例如土壤及植物体内分离出的活性菌株及其活性代谢产物的种类具有极高的重复率。目前,对新疆达坂城盐湖嗜盐细菌的研究较少,需要进一步挖掘达坂城盐湖的嗜盐菌资源。【拟解决的关键问题】采集达坂城盐湖湖边淤泥,选用不同的分离培养基对其进行嗜盐细菌的分离培养,根据形态进行分离,结合16S rRNA 基因序列,对部分放线菌开展活性研究,为研究新疆达坂盐湖地区嗜盐细菌物种基因资源的多样性提供理论依据。
1 材料与方法
1.1 材 料
1.1.1 样 品
于新疆达坂城盐湖(88°03′53″~88°12′15″E, 43°21′00″~43°25′25″N),湖水边缘,用采集器采集15~20 cm深处的淤泥样品,分装于无菌密封袋,置于4℃冰箱保存。
1.1.2 培养基
M-2(海藻糖-脯氨酸)培养基:Trehalose 6g,Proline 1 g,KNO30.5 g,Na2HPO40.3 g,MgSO4·7H2O 0.2 g,CaCl20.5 g,agar15 g,H2O 1 L,pH 7.2
MM琼脂培养基[7]:glucose 0.5 g, yeast extract 0.5 g,K2HPO40.3 g,NaCl 0.5 g,MgSO4·7H2O 1 g,FeSO4·7H2O 0.01 g,CaCO30.1 g,MnCl·4H2O 0.02 g,ZnSO40.07 g,agar 15 g,H2O 1 L,pH 7.2。
M-7(甘油-天门冬酰胺)培养基:Glycerol 10.0 g,Asparagine 1 g,K2HPO41 g,微量盐1 ml,agar 15 g,H2O 1 L,pH 7.2。
M-1(TWYE):Yeast 0.25 g, K2HPO40.5 g,agar 15 g,H2O 1 L,pH 7.2。
每种培养基灭菌前均添加终浓度为25 mg/L的萘啶酮酸和50 mg/L的制霉菌素。
拮抗植物病原菌采用PDA培养基。
1.1.3 主要试剂
NaOH、NaCl、MgSO4、MnCl2等均为化学纯。
1.1.4 植物病原菌
西瓜枯萎(尖孢镰刀菌Fusariumoxysporum)、水稻恶苗(串珠镰孢菌Fusariummoliniforme)、棉花枯萎(尖孢镰刀菌Fusariumoxysporum)、核桃腐烂病(壳囊孢Cycosporasp.)、苹果树腐烂病(壳囊孢Valsamalivar.mali)。以上病原菌由中国农业大学种子健康中心提供。
肿瘤细胞:肿瘤细胞采用人肝癌细胞HepG2,由浙江大学海洋学院提供。
1.2 方 法
1.2.1 菌株的分离和纯化
称取6 g土样,在干燥箱中进行70℃烘干处理2 h[8],之后从中称取2 g加人盛有18 mL无菌水的三角烧瓶中,振荡1 h后吸取1 mL土壤悬浮液到含9 mL无菌水的试管中,制成10-2的土壤悬液,按10倍法依次稀释至10-4。吸取10-2和10-4浓度的稀释液各0.2 mL,分别接于上述平板培养基上,每个稀释度2个重复,用涂布棒均匀涂布后置于30和37℃恒温下倒置培养,并采用划线分离法对菌株进一步的纯化分离,4℃斜面保存,根据形态特征和培养特征对放线菌进行鉴定[9]。
1.2.2 相关模式菌株
构建系统进化树外源菌:Bacillussubtilis(AS1.1849)。
1.2.3 16S rRNA序列的测定
1.2.3.1 细菌总DNA的提取
在灭菌的2 mL/1.5mL的离心管中加入20 μL 无菌水,用枪头挑取适量菌体,混匀并标记。100℃沸水浴沸腾5 min,放入-20℃冷冻5 min,此步骤重复2次,28℃,5 000 r/min,离心3 min。
1.2.3.2 16S rRNA序列的PCR扩增
采用细菌通用引物27F(AGAGTTTGATCCTGGCTCAG-3`)和1492R(5’-GGTTACCTTGTTACGACTT-3`)扩增菌株16S rRNA 序列。PCR扩增程序:95℃预变性5 min,主循环(94℃,30 s;54℃,30 s;72℃,30 s),循环30次,终延伸72℃,7 min,4℃保存。扩增中利用水代替 DNA 模板作为空白对照。PCR扩增产物采用1% 琼脂糖凝胶电泳检测,阳性产物送至北京鼎国昌盛生物技术责任股份有限公司进行纯化测序,测序结果与 GenBank 的序列进行BLAST比对。
1.2.4 基于16S rRNA 的系统发育学
用MEGA 7.0的 Neighbor-Joining 法构建16S rRNA 相似序列系统发育进化树的构建。
1.2.5 菌株发酵液以及粗提物的制备
在超净工作台内,用10 μL的枪头挑取放线菌菌落,转接于甘油-天门冬酰胺液体培养基,30℃,180 r/min,恒温震荡培养5~7 d。待发酵完成后,用乙酸乙酯等体积混合震荡萃取发酵液3次,回收浓缩得发酵液粗提物。
1.2.6 拮抗菌株筛选
1.2.6.1 抗菌活性
采用平板对峙法测定所分离到的放线菌对植物病原菌的抑菌效果。在超净台内,用5 mm打孔器将PDA平板打孔,每个平板打四个孔,打孔位置距离培养皿中心距离相等,用灭菌竹签挑取植物病原菌的新鲜孢子接于平板中心,再用枪头吸取100 μL离心后的菌株发酵液上清液,注入孔内,以不接发酵液的平板为对照,28℃培养, 5~7 d后观察记录有无抑菌圈产生。
1.2.6.2 抗肿瘤活性筛选
选用人源细胞株HepG2(人肝癌细胞株)为检测对象,采用磺酰罗丹明B(SRB)方法[10]检测菌株发酵液粗提物对HepG2细胞增殖的抑制作用,菌株粗提物用DMSO配成10 mg/ml得浓度。
收集对数生长期HepG2细胞并以8×103细胞/孔的密度种于96孔板内,过夜培养后加药,以阿霉素为阳性药,设置3个重复,继续培养。72 h后取出96孔板,弃去培养基后每孔加入100 μL 10%的三氯乙酸水溶液,4℃冰箱中固定至少2 h。然后取出96孔板,弃去三氯乙酸溶液,并用缓慢流动的水冲洗至少4遍并烘干,再加入80 μL 1%醋酸配制的0.4% SRB溶液染色20 min,期间配置1%醋酸水溶液,染色结束后用1%醋酸水溶液冲洗4遍以去除未结合的染料,再次烘干至完全干燥后每孔加入100 μL 10 mM Tris溶液,并震荡使与蛋白结合的染料完全溶解,再置于酶标仪中于515 nm处检测吸光度。
抑制率=(对照组OD值-给药组OD值)/对照组OD值×100%。
2 结果与分析
2.1 达坂城盐湖嗜盐细菌的分离鉴定
研究表明,共获得14个属18株嗜盐细菌,其中 M-7培养基(甘油-天门冬酰胺) 共分离到 11个属 13个不同种的嗜盐细菌。表1,图1
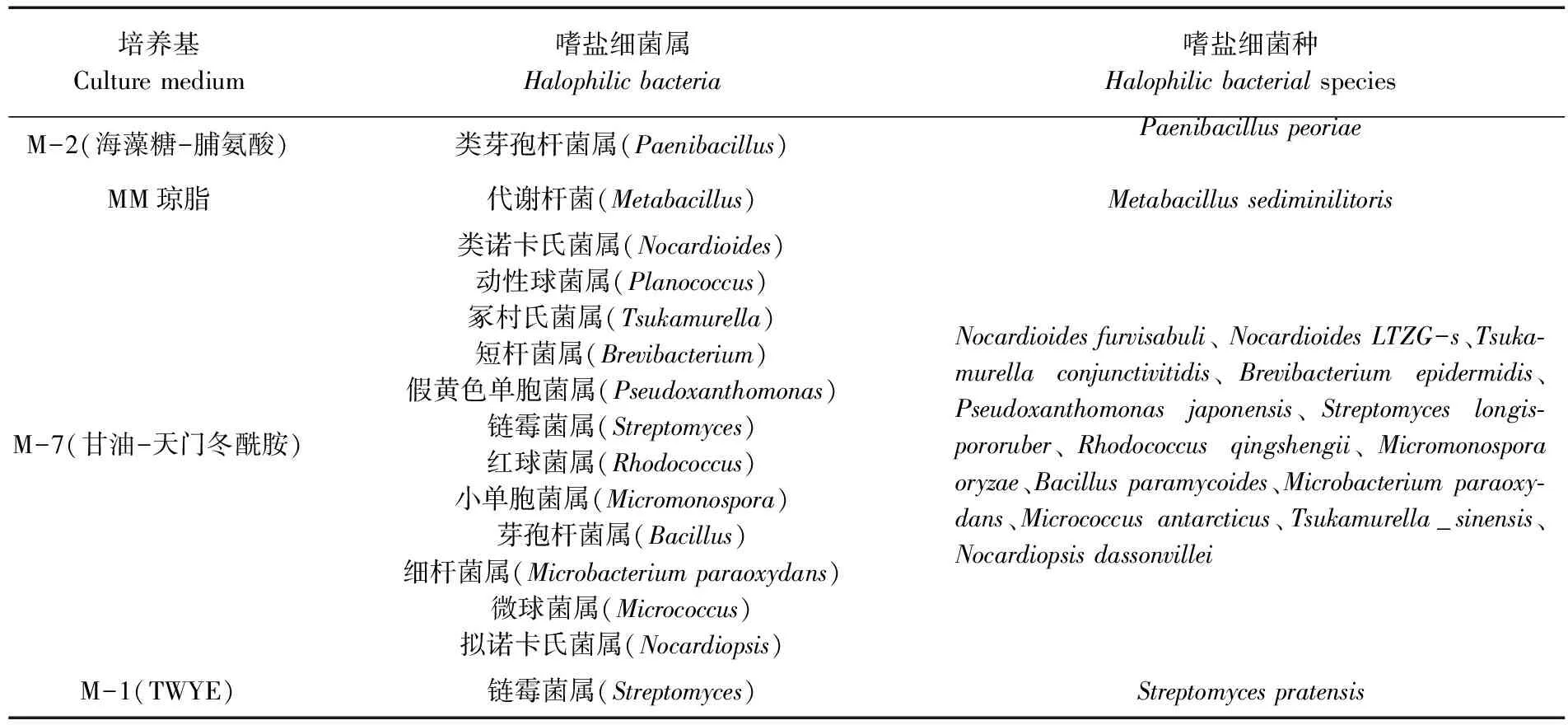
表1 不同培养基分离到的嗜盐细菌种属
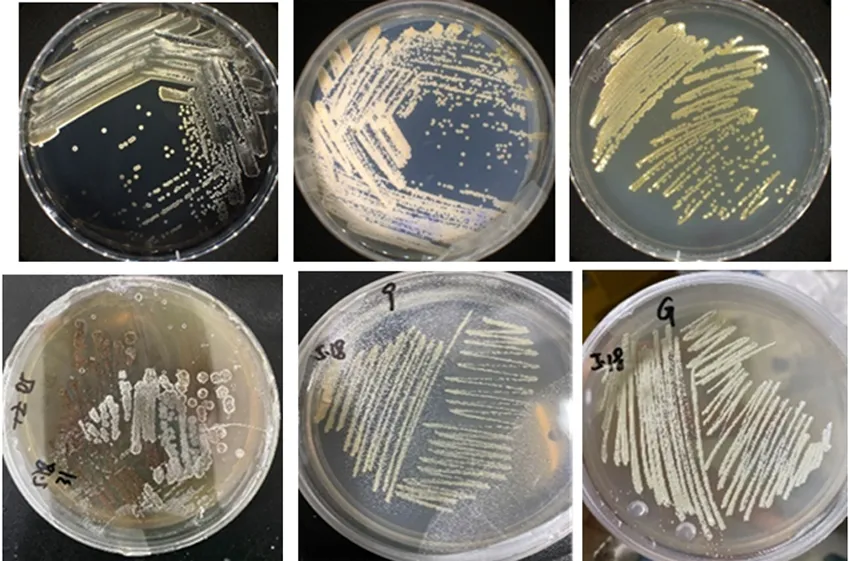
注:a:菌株DBC23;b:菌株DBC30;c:菌株DBC31;d:菌株DBC5;e:菌株DBC9;f:菌株DBC10
2.2 菌株的系统发育学
研究表明,菌株18位于Bacillus的属中,和众多Bacillus在同一个分支上,菌株18与其亲缘关系较近,同源性为95.93%。菌株23属于Nocardioides属,且单独位于一个较长分支上,同源性为97.43%。图2,图3

图2 菌株18基于 16S rRNA 序列系统发育树
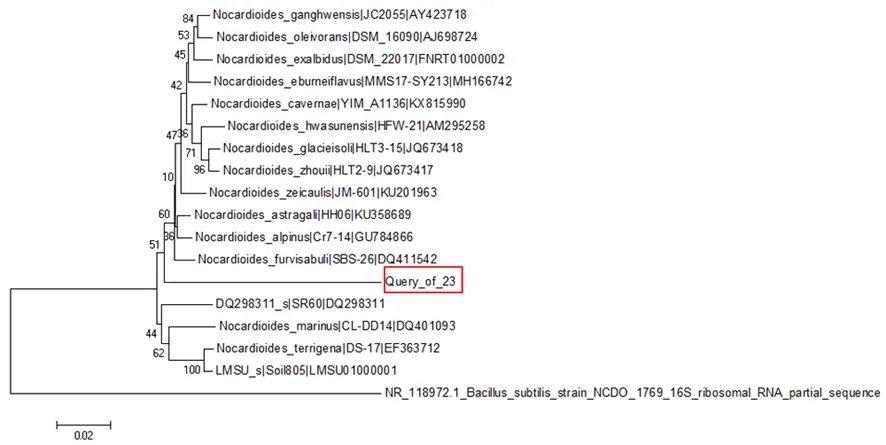
图3 菌株23基于 16S rRNA 序列系统发育树
2.3 放线菌发酵液的抗菌活性
研究表明,DBC26、DBC31和DBC5对病原菌具有潜在的抗菌活性,其中DBC26和DBC31有拮抗棉花枯萎病菌活性,为Tsukamurellasinensis和Nocardiopsiscrassaminis,DBC5有抗水稻恶苗病菌活性,是Streptomyceslongispororuber。表3

表3 达坂城盐湖嗜盐放线菌抗菌活性
2.4 放线菌粗提物抗肿瘤活性
研究表明,菌株DBC5对HepG2细胞增殖的抑制率最高,达到了96.4%;其次是菌株DBC9,抑制率为91.8%;菌株DBC10和菌株DBC26的抑制率分别为87.8%和83.2%;菌株DBC24的抑制活性较弱,抑制率仅为16.7%,而菌株DBC3对HepG2细胞增殖没有抑制率。表4
3 讨 论
3.1研究共获得14个属18株嗜盐细菌,其中芽孢杆菌属(Bacillus)、诺卡氏菌属(Nocardiopsis)和链霉菌属(Streptomyces)为优势菌属,且放线菌大多为稀有放线菌。其中 M-7培养基(甘油-天门冬酰胺) 共分离到 11个属 13个不同种的嗜盐细菌,分离效果好于其他3种培养基, 认为M-7(甘油-天门冬酰胺)培养基分离盐湖可以得到较多种类的嗜盐细菌。可能是因为甘油碳源,能适当的调节培养基渗透压;而L-天门冬酰胺可能是盐湖微生物赖以生存的一种有机酸,满足了高盐环境嗜盐细菌的生理和特殊的营养需求。用M-7(甘油-天门冬酰胺)培养基分离死海嗜盐放线菌[11]。

表4 盐湖放线菌发酵液粗提物对HepG2细胞的抑制作用
3.2极端环境生长的放线菌所需的营养条件不同于普通环境的放线菌,盐湖环境除了高盐的特性,还存在着丰富的化学离子,如Na+、K+、Mg2+、SO42-等,李二阳[12]和唐蜀昆[13]等研究发现当湖水化学成分出现差异,会导致菌群结构特异演化。离子成分对盐湖微生物的生长起着驱动作用,为培养基成分的选择提供了参考。因此,在分离盐湖环境的菌株时,应充分考虑分离条件和培养基的设计。分离出较多放线菌的M-7培养基中,成分包括了除Na+外的K+、Mg2+。此外,试验设置了30和37℃2个温度梯度,发现37℃条件下分离放线菌的效果优于30℃,冢村氏菌(Tsukamurella)是从达坂城盐湖分离出的稀有放线菌,放线菌的分离存在着温度效应[14]。
3.3菌株DBC23和菌株DBC18的16S rRNA序列与已知的菌株 16S rRNA序列相似度分别在97.43%和95.53%,为潜在新种,需要进一步研究。抑菌实验中筛选到3株具有拮抗活性的放线菌,进行了初步的拮抗活性筛选,其中具有拮抗活性的放线菌3株,属于链霉菌属(Streptomyces)、拟诺卡氏菌属(Nocardiopsis)冢村氏菌属(Tsukamurella)。而关于冢村氏菌的活性报道较少,放线菌(DBC26)值得进一步研究。
4 结 论
从新疆达坂城盐湖湖边15-20cm深的淤泥样品中分离出了18株菌,鉴定为14个属,是Bacillus(2株)、Pseudoxanthomonas(1株)、Rhodococcus(1株)、Micromonospora(1株)、Streptomyces(2株)、Brevibacterium(1株)、Tsukamurella(2株)、Planococcus(1株)、Nocardiodes(1株)、Metabacillus(1株)、Paenibacillus(1株)、Microbacterium(1株)、Micrococcus(1株)、Nocardiopsis(2株)。其中冢村氏菌(Tsukamurella)是从达坂城盐湖中分离出。对放线菌进行了抗植物病原菌测试,其中DBC5、DBC26和DBC31分别对水稻恶苗和棉花枯萎具有初步的抑制效果,DBC5、DBC9的细胞增殖抑制率均在90%以上。
猜你喜欢
江苏农业学报(2022年4期)2022-09-07 06:21:22
云南化工(2021年5期)2021-12-21 07:41:20
中国果业信息(2021年5期)2021-12-05 22:10:28
国际呼吸杂志(2019年22期)2019-12-09 09:20:36
北京文学(2017年12期)2017-12-12 20:23:37
吐鲁番(2015年1期)2015-11-17 19:53:54
应用海洋学学报(2014年4期)2014-11-22 07:43:54
天然产物研究与开发(2014年6期)2014-04-27 14:15:56
新晨(2013年1期)2014-04-03 03:40:48
中国医学科学院学报(2013年2期)2013-03-11 20:25:49
