
西太平洋深海底栖多毛类动物地理分布特征
2023-07-27 06:38:44王跃云严润玄王春生
海洋学研究 2023年2期
王跃云,严润玄,3,王春生*
(1.自然资源部海洋生态系统动力学实验室,浙江 杭州 310012; 2.自然资源部第二海洋研究所,浙江 杭州 310012; 3.河海大学 海洋学院,江苏 南京 210024)
0 引言
若以深度大于200 m的海洋水体来定义,深海区域超过了海洋的90%[1];若以深度大于1 000 m来定义,深海区域覆盖了地球表面的60%[2]。尽管人们对深海深度范围的界定不同,但都不可否认深海是地球上最大的生物分布区。近几十年来,随着一些深海新生境的发现,如海山、峡谷、海脊、冷泉、热液口、多金属结核区等,很多新物种也被不断地发现和描述[3]。除了发现和描述新物种外,深海生态系统的研究主题还包括物种数量、生物量、地理分布范围等[4]。但总体上人类对深海物种多样性的认识还十分有限。据估计海洋中的物种数量有220万种,其中有91%的物种尚未被描述[5],而对于物种在深海中的分布同样存在诸多认知上的空白。
多毛类动物是环节动物门(Annelida)中最大的类群,已经记录了80 多个科11 000多个有效种[6]。虽然在深海海底环境中,无论是在特殊的化能合成环境还是在普通的沉积环境中,多毛类动物的密度和物种丰富度在大型底栖生物类群中占优势[7-10],但是多毛类动物的分类学和生态学研究相对于其他深海底栖动物(如珊瑚、海绵)来说,关注度较低[11]。受采样技术手段等的限制,目前有关深海多毛类动物多样性的研究仅有限地在深海平原、盆地、海山等地貌区开展,并且,即便在研究相对较多的洋中脊海山系统,也还有90%的区域未被探索[12-13]。我国深海底栖多毛类动物分类学研究尚处于起步阶段[13-14],近年来在东海东侧冲绳海槽热液口附近[15]、台湾西南部冷泉区[16]、马里亚纳海沟[17]、马努斯盆地热液口[18-19]、卡罗琳海山[20]、采薇海山[21]、马里亚纳海盆中的KIOST海山[22]以及马绍尔海山群的一无名海山[23]发现并描述了多个多毛类新属、新种和新记录种。除了太平洋外,在印度洋热液区也发现了多个多毛类新物种[24-28]。这些深海新物种大部分来源于热液喷口,少部分来自深水海山,有关其地理分布特征的研究仅在少量特殊生境开展,如热液口多鳞虫属Branchinotogluma的生物地理模式[18]和印度洋中脊的热液口动物地理区划[29]。
2013年,WATLING et al[30]根据海底水文(水深、温度、盐度等)和颗粒有机物通量数据将全球深海海底生物地理区划分成14个下次深海区(lower bathyal provinces,801~3 500 m)和14个深海区(abyssal provinces,3 501~6 500 m),其中,西太平洋的下次深海区包括北太平洋北部区(Northern Pacific Boreal)、西太平洋区(West Pacific)和北太平洋区(North Pacific),深海区包括北太平洋区(North Pacific)、中北太平洋区(North Central Pacific)、西太平洋海盆区(Western Pacific Basins)、赤道太平洋区(Equatorial Pacific)、南太平洋区(South Pacific)和印度洋区(Indian)。但是,这一生物地理区划尚需物种分布数据进行验证。
全球开放数据库信息的完善极大促进了动物地理区划研究的发展[31]。海洋生物地理信息数据库(Ocean Biogeographic Information System,OBIS)源于“全球海洋生物普查计划”,是目前全球最大的海洋生物多样性信息开放数据库,也是全球生物多样性信息网络原始数据及网络生命大百科物种页面的主要提供者[32]。热液喷口是深海物种多样性的热点区,分布有很多特有性物种。全球海底活动热液口数据库提供热液口名称、地理位置、深度等信息[33],为研究深海生物地理分布提供了重要地质学背景信息。
本文将深海的深度范围界定为1 000 m以深,根据OBIS中多毛类动物的采集记录,结合海底热液喷口分布信息,分析西太平洋深海底栖多毛类动物的地理分布特征。
1 数据和方法
1.1 数据来源
本文研究区为西太平洋(55°S—60°N、110°E—180°E)海域。从OBIS(https://obis.org/)下载1 000 m以深底栖多毛类动物的原始记录,剔除其中错误数据、重复数据以及非底栖多毛类数据。从全球海底活动热液口数据库下载含热液喷口经纬度的原始数据,用于划分地理单元。水深数据通过R软件中的Marmap 包[34]从美国国家海洋和大气管理局网站(http://www.noaa.gov)获取。
1.2 数据处理
1.2.1 聚类分析与集合分析
将研究区按照10°×10°划分成84个栅格,根据栅格内物种分布信息构建“栅格×物种”矩阵。利用“栅格×物种”矩阵进行聚类分析,主要步骤如下:去除栅格中物种数<2的栅格,根据Jaccard相似性指数计算相关矩阵,利用Ward法进行聚类分析,对聚类结果分组,得到西太平洋深海底栖多毛类分布相似性聚类图。利用“栅格×物种”数据矩阵进行集合分析,研究栅格内物种数量及其共有物种数。聚类分析和集合分析分别通过R软件的Clustsig包[35]和UpSetR包[36]进行。
1.2.2 特有性简约分析
特有性简约分析(parsimony analysis of endemicity,PAE)是一种历史生物地理学方法[37],该分析方法类似于分支分析,通过区域共有分类群描述区域间的历史关系,广泛应用于生物地理学研究。特有性简约分析的主要步骤包括:1)在研究区域内选择一组生物地理单元;2)记录生物类群的地理分布;3)根据生物类群的地理分布信息构建“地理单元×类群”矩阵;4)利用简约算法分析数据矩阵,生成分支图,分析生物地理格局、特有性分布区等。
结合WATLING et al[30]提出的全球深海海底生物地理区,根据地理特征(海沟、海底平原、热液口、距大陆距离)人为将西太平洋划分为8个地理单元(图1a),依次为:日本海槽-千岛-堪察加海沟区(Jkk),日本海区(SoJ),相模湾区(Sb),冲绳海槽区(Okinawa),马努斯盆地区(Mb),斐济海盆区(NFb),新西兰区(NZ),澳大利亚东侧区(EA);构建 “地理单元×物种”矩阵,假定一个物种分布为0的地理单元为分支图定根,通过R软件的Phangorn包[38]进行分析,采用1 000次自展抽样估计分支支持度,分支支持度越高则分支的可信性越强。

图1 深海底栖多毛类动物记录分布点(a)及不同水深段物种丰富度分布(b)
2 结果
2.1 物种分布特征
经数据处理、分析,在西太平洋深海(>1 000 m)共得到底栖多毛类物种318个, 记录1 425条,记录多分布于海沟、海山等地貌单元,水深小于3 000 m(图1a)。物种丰富度总体呈随水深增加递减趋势,其中,在水深1 000~1 700 m,物种数急剧下降,在 2 500~3 000 m和4 000~4 500 m,各呈现出一小高峰(图1b)。
研究区共记录到多毛类51个科(图2)。不同科中物种记录数极不均匀(表1),其中记录数最多的前10个科为阿尔文虫科(Alvinellidae)、多鳞虫科(Polynoidae)、鳞沙蚕科(Aphroditidae)、米列虫科(Melinnidae)、叉螠科(Bonelliidae)、欧努菲虫科(Onuphidae)、双栉虫科(Ampharetidae)、竹节虫科(Maldanidae)、西伯加虫科(Siboglinidae)、仙虫科(Amphinomidae),其中最多的阿尔文虫科含247条物种分布记录。在种水平上,物种记录最多的为Paralvinellahessleri,含有237条记录,远多于第二记录的物种Melinnopsischadwicki,两者同属阿尔文虫科,特有性地分布在深海热液口环境中。物种丰富度最高的前10个科为多鳞虫科、欧努菲虫科、叉螠科、裂虫科(Syllidae)、西伯加虫科、竹节虫科、双栉虫科、沙蚕科(Nereididae)、金扇虫科(Chrysopetalidae)、索沙蚕科(Lumbrineridae),其中,多鳞虫科的分布水深范围最广(1 000 ~10 000 m),叉螠科、西伯加虫科和竹节虫科的分布水深范围也较广,三者的最大分布水深均超过7 000 m。

表1 西太平洋深海底栖多毛类记录数前10个科

图2 不同科中的物种丰富度
2.2 聚类及集合分析
根据物种分布记录,研究区含物种分布信息的栅格共28个(图3a),物种318个,构建了28×318的矩阵,聚类分析得到各栅格间的相似性,结果见图3b。西太平洋深海底栖多毛类动物的分布明显分为8组:冲绳海槽与日本东南部聚类组(栅格17, 23, 32),热液口聚类组(栅格47, 56, 63),澳大利亚东侧聚类组(栅格61, 67, 68, 74),新西兰区聚类组(栅格69, 70, 76, 77),白令海-鄂霍茨克海聚类组(栅格7, 11, 12, 25, 38),菲律宾吕宋岛聚类组(栅格29, 30)以及其他2个聚类组(栅格10, 18, 36, 45;栅格37, 43, 83)。总体上,地理距离较近的区域,其多毛类组成相似,南北半球的差异较为明显。
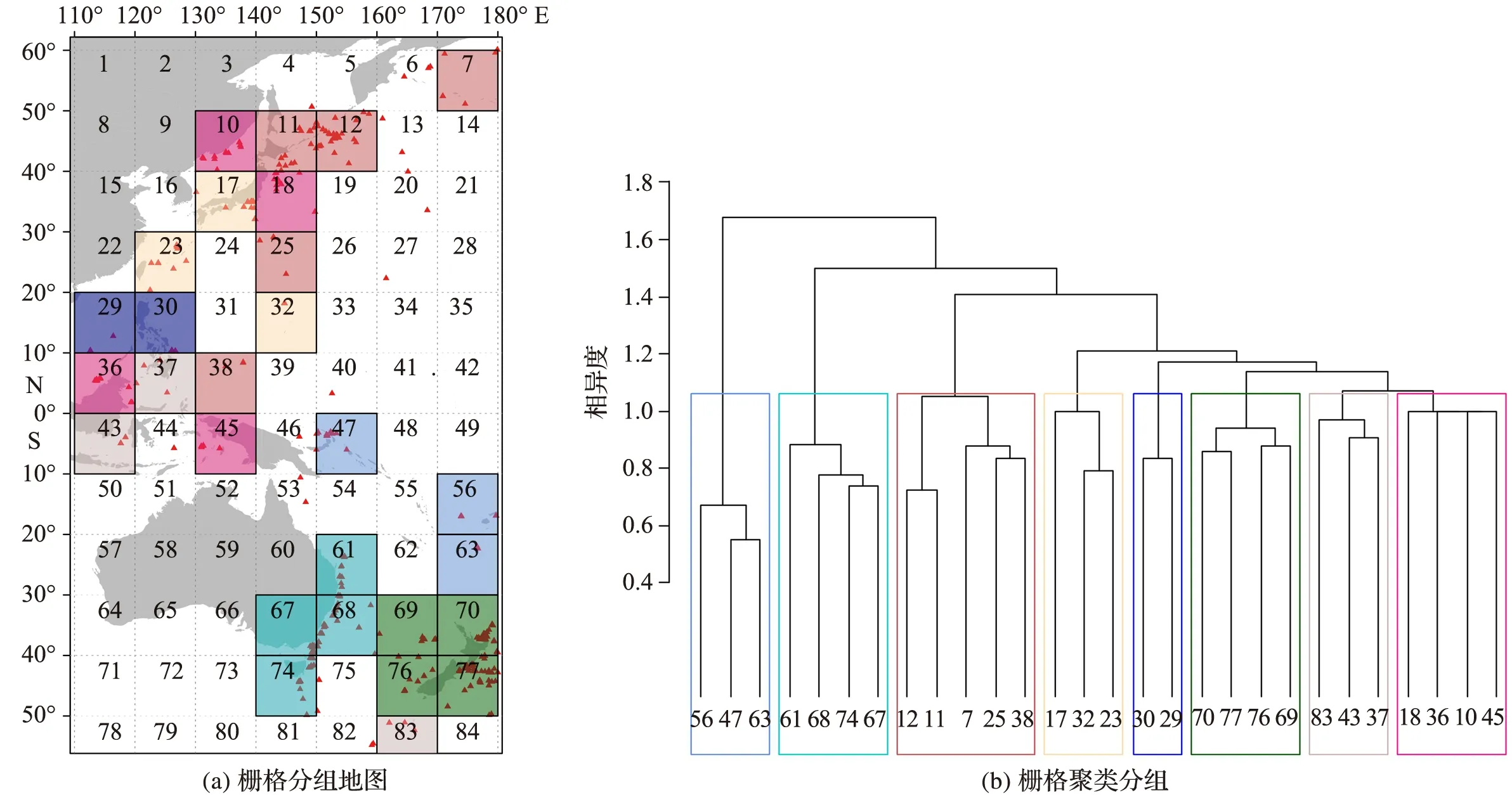
图3 基于Jaccard距离的Ward聚类
集合分析结果(图4)显示,总体上各栅格中特有种(仅出现于某个栅格的物种)比例较高,除23、25、38、63和83号栅格外,其余23个栅格中特有种的比例均高于25%;共有种(出现于2个及以上栅格的物种)比例较低,并且共有种的数量一般仅为1~2种。63号栅格共含14个物种,其中仅1个特有种,大部分物种与47、56号栅格共有,经进一步检视发现,这3个栅格记录的物种大多特有性地分布在深海热液口。77和68号栅格的物种数最多,其中特有种比例分别高达73%、35%。
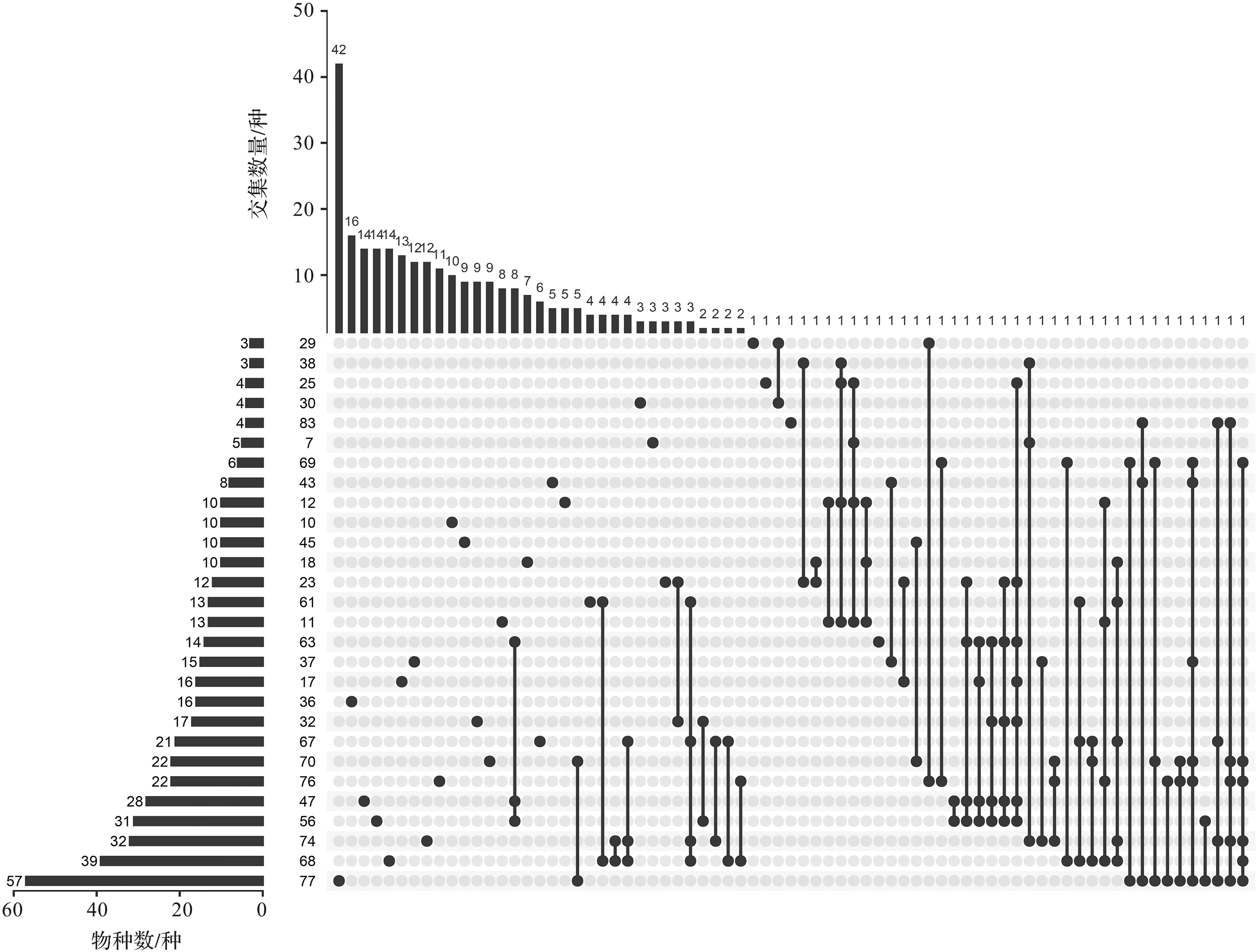
图4 栅格物种集合分析图
2.3 特有性简约分析
PAE分支图显示人为划分的8个地理单元可分成4个分支:日本海区(SoJ)和相模湾区(Sb)分别形成独立的一个分支,冲绳海槽区(Okinawa)、马努斯盆地区(Mb)及斐济海盆区(NFb)形成一个分支,日本海槽-千岛-堪察加海沟区(Jkk)、澳大利亚东侧区及(EA)及新西兰区(NZ)形成一个分支(图5a)。在地图上(图5b)可见,日本海区(SoJ)与其他地理单元存在一定的地理隔离;相模湾区(Sb)靠近大陆,深度平均仅为1 200 m,是所有地理单元中水深最浅的;冲绳海槽区(Okinawa)、马努斯盆地区(Mb)及斐济海盆区(NFb)均有密集的热液喷口分布;日本海槽-千岛-堪察加海沟区(Jkk)位于北半球,澳大利亚东侧区(EA)和新西兰区(NZ)位于南半球,虽然地理距离较远,但均以海沟和平原为特征。

Jkk—日本海槽-千岛-堪察加海沟区;SoJ—日本海区;Sb—相模湾区;Okinawa—冲绳海槽区;Mb—马努斯盆地区;NFb—斐济海盆区;NZ—新西兰区;EA—澳大利亚东侧区)
3 讨论
西太平洋深海的多毛类动物多样性研究多集中于靠近陆地的陆坡、海沟以及热液和冷泉等特殊地貌单元[15-23],相比于近海研究,尚有很大不足,尤其是在广袤的深海盆地和地形复杂的海山区域,还存在很多调查空白区。统计显示在中国南海和菲律宾海域已描述的多毛类为1 037种,隶属345属60科[39],约占全球已描述多毛类物种数的9%。在这些物种中,沙蚕科和多鳞虫科物种最多,其中大多数物种的模式产地位于近海,位于深海的很少。人们对于深海多毛类动物的物种多样性的认识还存在很多未知,在深海调查样品中常出现极高比例未被描述过的新物种,有时比例甚至超过1/2[40-42]。这种未知可能会对生物地理学分析结果产生影响,如曾被认为是在太平洋热液口特有性分布的阿尔文虫科,后发现在北印度洋热液口也有分布[43]。
一项深海(>2 000 m)多毛类动物普查发现,多鳞虫科是第一大类群,拥有最多的物种数量[44],与本文结果一致(图2)。在不同的地理环境中,多毛类的优势类群是不同的。如在千岛-勘察加海沟附近的深海平原区,优势类群依次为双栉虫科、海蛹科(Opheliidae)、竹节虫科[40],在南极洲的威德尔海盆和南桑威奇陆坡,数量最多的为异毛虫科(Paraonidae)和缨鳃虫科(Sabellidae)[10],而在东太平洋隆起和加拉帕戈斯附近的一些热液口附近则以巨型管栖蠕虫(西伯加虫科)为主要类群。
水深是决定深海生物群落结构最重要的因素之一,它与其他因素,如温度、压力、氧气、水团结构和食物供应等相关[45]。研究发现深海物种多样性在垂直分布上呈单峰曲线变化,一般在较浅的大陆坡(200~2 000 m)或大陆隆起(2 000~4 000 m)生境中物种多样性的值较高,在较深的生境中物种多样性的值较低[46-49]。而在本文中(图1),西太平洋底栖多毛类动物的物种多样性整体上随深度增加而降低,但在 2 500~3 000 m以及 4 000~4 500 m生境中,多样性略有增加。出现该现象的原因尚不十分明确,已有研究认为可能与特定生境的环境异质性相关,尤其是与沉积物类型的多样性相关[50]。
由于深海动物多样性数据的缺乏,目前深海动物地理区系划分主要依据海洋学指标(如地形、深度、水团结构、颗粒有机碳通量、底质特征等)进行[30,51],通过生物分布数据对这些生物地理分区进行校正,将增进对深海动物群落地区差异性的理解。在WATLING et al[30]的海洋生物地理分区中,Jkk与SoJ同属一个区(下次深海区的北太平洋北部区,深海区的北太平洋区),而在本研究中无论是聚类分析结果(图3)还是PAE分析结果(图5),均显示两者属于不同的区系。虽然Jkk和SoJ的地理距离较近,但因日本海基本与太平洋分隔,两地形成不同的深海动物区系是可能的。在WATLING et al[30]的研究中,Sb属下次深海区的西太平洋区,与本研究中Sb独立成支的结果(图5)一致。Jkk、EA和NZ属于同一个分支,其中EA和NZ之间支持度更大,与Jkk的支持度较低(图5),表明相邻深海的物种类群接近,支持WATLING et al[30]的EA和NZ同属新西兰-克马德克区的划分。
4 结论
多毛类动物是深海环境中最主要的大型底栖动物类群之一,其物种多样性以及地理分布特征尚未被完全揭示。西太平洋深海底栖多毛类动物中物种数量最高的类群为多鳞虫科,且它具有最为宽广的深度适应范围;物种多样性总体上随水深增加而降低,但在 2 500~3 000 m以及4 000~4 500 m水深处表现出较高的多样性。深海物种的特有性分布可能普遍存在,尤其是在热液口环境,形成了以热液口特有种为特征的多毛类动物区系。基于地理分区与前人基于海洋环境数据的动物地理区划基本一致,相近的地理环境中多毛类组成相似,但有部分区域存在异常,如日本海与日本海槽-千岛-堪察加海沟区的多毛类可能并不同属一个生物地理区。
目前深海动物多样性数据缺乏,特别是地形复杂的海山、广袤的深海平原等生境调查数据的缺乏,对认识深海动物区系构成了障碍,建议开展更多针对深海平原和海山环境的动物多样性调查,为理解深海多样性分布格局提供重要支撑。
猜你喜欢
Plasma Science and Technology(2020年9期)2020-09-14 01:13:08
37°女人(2019年4期)2019-04-20 02:17:42
西南石油大学学报(自然科学版)(2018年4期)2018-08-02 05:42:18
海洋与湖沼(2017年6期)2017-03-31 06:18:19
作品(2016年10期)2016-12-06 10:41:54
大江南北(2016年3期)2016-11-22 07:37:20
西藏科技(2016年5期)2016-09-26 12:16:42
石油知识(2016年2期)2016-02-28 16:20:19
钻探工程(2012年1期)2012-01-27 19:48:18
计算机应用文摘(2010年22期)2010-04-29 05:13:37
