
蓝光对盐胁迫下番茄幼苗生长的影响
2023-07-23 13:47刘增喜许可欣祝海燕孙小桉张敬敏于丽艳桑晓明
中国瓜菜 2023年7期
刘增喜,冯 棣,许可欣,祝海燕,孙小桉,张敬敏,于丽艳,桑晓明
(1.山东省高校设施园艺重点实验室·潍坊科技学院 山东寿光 262700;2.寿光市农业技术推广中心 山东寿光 262700)
番茄(Solanum lycopersicumL.)是每个国家不可或缺的蔬菜作物,具有较高的营养价值和经济价值[1],同时还是科研人员用于遗传学及抗逆生理生化研究的重要模式植物[2]。近年来,盐胁迫对番茄的生长发育带来越来越严重的损害。因此,从调控番茄盐分胁迫下的光质入手,对提高番茄的抗盐能力以及番茄抗盐高效栽培具有很重要的意义。众所周知,光是调节植物生长发育的重要环境因子。前人研究结果表明,光质会影响植物的生长发育[3],而且它还对植物的形态建成、物质代谢及基因表达等有明显的调节效果[4]。实际上,植物在不同光质下的生长发育情况也不完全一样,并且与大田环境相比较,温室内光质会发生很大变化,所以要改变温室内的光环境,就需要通过补光来实现,最后达到调节植物生长的效果[5]。不同光源补光能极大改善番茄幼苗形态和提高番茄幼苗全株的干鲜质量。红光能够加快兰花叶片生长,同时还可以使叶绿素含量降低,但这一过程可以被蓝光逆转[6]。蓝光对植物的光形态建成、向光性、光合作用、胚轴伸长以及酶调节和合成等有明显的影响[7]。蓝光能促进大白菜抗坏血酸基因生物合成以及转录水平提高,进一步提高了抗坏血酸含量[8]。蓝光还能提高西蓝花芽苗菜营养品质[9],而且还可以促进茶树叶片花青素和儿茶素的积累[10]。但关于蓝光对盐胁迫下番茄幼苗的生长发育影响方面的研究鲜见报道。笔者的研究旨在探明蓝光对番茄幼苗形态和生理指标、根系生长特征的影响,以期为解析蓝光调控番茄耐盐机制提供科学依据。
1 材料与方法
1.1 材料
试验于2021 年9—10 月在潍坊科技学院生物技术研发中心进行。供试番茄幼苗材料为玉玲珑,由寿光市潍科种业有限公司提供。营养液配方选用山崎番茄配方。
1.2 试验设计
在LED 蓝光下设置3 个NaCl 盐分浓度梯度(0、50、100 mmol·L-1),并以常用补光模式LED 红、蓝光光照度之比为4∶1 下3 个NaCl 盐分处理为对照组,共6 个处理。按照盐浓度由低到高各处理分别标记为RB1、RB2、RB3、B1、B2、B3,8 次重复。将番茄幼苗穴盘育苗至3 叶1 心,之后移入水培杯中,每杯1 棵,随机摆放,每个处理8 株。每3 d 更换1 次营养液。昼/夜两个阶段培养时间各12 h。红、蓝光光照度比为4∶1 和蓝光光谱分布如图1 所示,光合有效辐射分别为405.62、316.08、298.59 μmol·m-2·s-1,光照度分别为2810、110、2100 lx。
1.3 测定项目与方法
1.3.1 生长指标测定 在移栽后30 d 测量每个处理下的所有番茄幼苗的株高,然后计算株高生长速率(即株高变化量与间隔时间之比)。从每个处理中取3 株生长一致的番茄,采用精度为0.001 g 的电子天平称量茎、叶鲜质量;之后将各部分在105 ℃下杀青0.5 h,在75 ℃下烘干至恒质量,使用精度为0.001 g 的电子天平称得干质量。取根系鲜样,使用根系扫描仪EPSON EXPRESSION 10000 XL 扫描样品,再使用根系分析系统WinRHIZO Pro分析各处理样品的总根长(cm)、总根表面积(cm2)、总根体积(cm3)、根平均直径(mm)。
1.3.2 生理指标测定 移栽后30 d 从每个处理中取3 株生长一致的番茄幼苗,摘取第3、4 片叶,采用95%乙醇-分光光度计法测定幼苗叶片的叶绿素a、叶绿素b 和类胡萝卜素含量[11],叶片的超氧化物歧化酶(SOD)、过氧化物酶(POD)、抗坏血酸过氧化物酶(APX)活性均采用试剂盒进行测定,试剂购买自苏州科铭生物技术有限公司。
1.4 数据分析
采用EXCEL 软件进行处理试验数据分析并绘制图表。采用SPSS 26.0 数据处理软件、LSD 法进行方差分析及显著性检验。
2 结果与分析
2.1 蓝光对盐胁迫下番茄株高生长速率的影响
由图2 可知,在相同光质下,番茄株高均随着盐胁迫程度的增加而降低。由图3 可知,在相同光质下,番茄幼苗株高生长速率均随着盐胁迫程度增加而降低,且各处理间差异显著;相同盐胁迫下,与RB1 处理相比,B1 的株高生长速率显著降低22.2%,但B2 与RB2 处理、B3 与RB3 处理株高生长速率差异均不显著。

图2 不同光质和盐胁迫下番茄植株生长情况
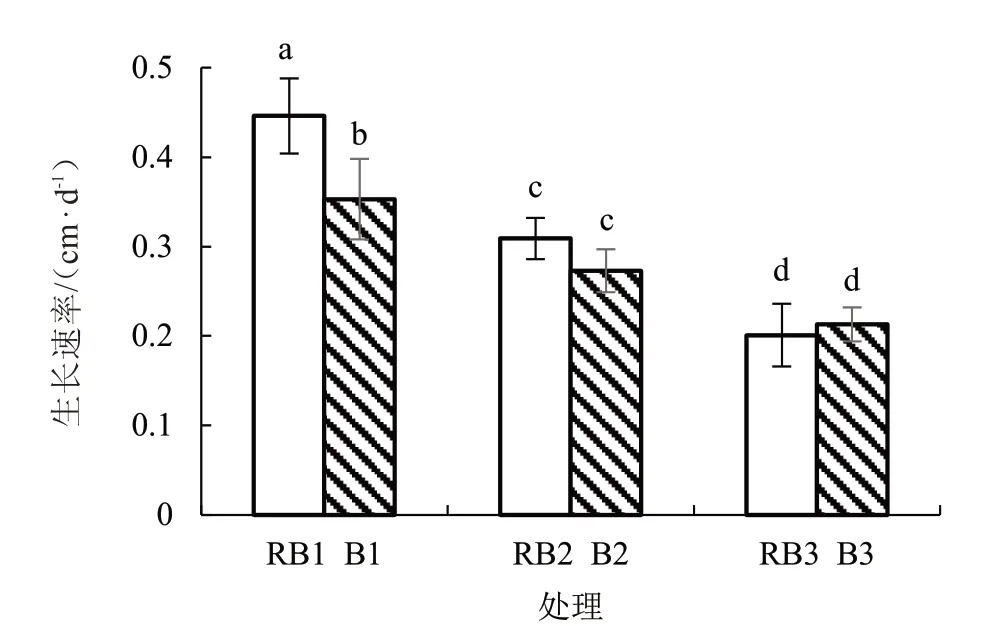
图3 不同光质和盐胁迫下番茄株高生长速率
可见,盐分胁迫会抑制番茄株高增长;在不同光质下,相对于RB 处理,B 处理在没有盐胁迫时不利于番茄株高增长,但在高盐(100 mmol·L-1)和低盐(50 mmol·L-1)胁迫下与RB 处理差异不显著。
2.2 蓝光对盐胁迫下番茄干鲜质量的影响
由表1 可以看出,在相同光质下,随着盐胁迫程度的增加,番茄茎和叶的鲜、干质量、根干质量、根冠比均降低。B2 与B3 处理茎鲜质量无显著差异,RB2 与RB3、B1 与B2 茎干质量均无显著差异,B2 与B3 叶干质量无显著差异,RB1 与RB2、RB2与RB3、B1 与B2、B2 与B3 根干质量均无显著差异,各处理间根冠比无显著差异。在相同盐胁迫下,不同光质处理在鲜/干质量方面,仅B1 处理的茎鲜/干质量、B2 处理的叶鲜质量、B3 处理的茎干质量均显著低于相应的RB 处理,降幅依次为14.81%、26.63%、13.19%、43.94%,其余指标在处理间差异不显著。可见,盐胁迫显著抑制了RB 茎/叶鲜质量、叶干质量、B 叶鲜质量;在2 种光质下,相对于RB 处理,B 处理在无盐胁迫下显著抑制了茎鲜/干质量,B 处理在低盐(50 mmol·L-1)胁迫下显著抑制了叶鲜质量,B 处理在高盐(100 mmol·L-1)胁迫下显著抑制了茎干质量。
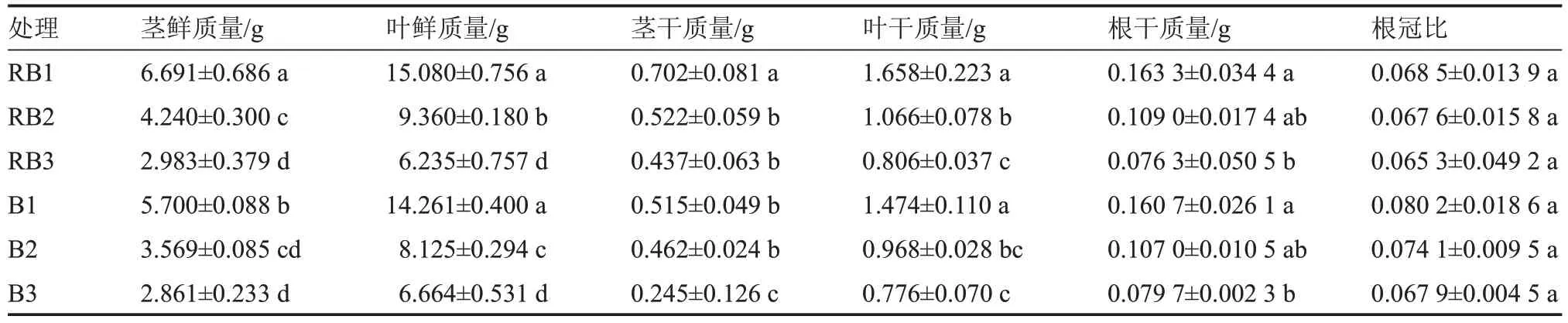
表1 不同光质和盐胁迫下番茄干鲜质量
2.3 蓝光对盐胁迫下番茄根系生长特征的影响
由表2 可以看出,在相同光质下,随着盐胁迫程度的增加,番茄的总根长、总根表面积、总根体积、根平均直径均逐渐降低,但RB2 与RB3 总根长、总根表面积、总根体积、根平均直径均无显著差异,B2 与B3 根平均直径无显著差异。在相同盐胁迫下,仅B3 处理的总根长显著小于RB3 处理,降幅为23.69%,其余指标在处理间无显著差异。可见,盐胁迫对番茄幼苗根系生长产生了显著抑制作用;相对于RB 处理,B 处理表现出在无盐胁迫下促进番茄根系生长,在低盐(50 mmol·L-1)胁迫下抑制番茄总根长和根平均直径;在高盐(100 mmol·L-1)胁迫下抑制根系长度的生长。

表2 不同光质和盐胁迫下番茄根系生长特征
2.4 蓝光对盐胁迫下番茄生理指标的影响
2.4.1 不同光质和盐胁迫下番茄幼苗叶绿素及类胡萝卜素含量的变化 由图4 可以看出,在RB 处理下,随着盐胁迫程度的增加,叶绿素a、b、a+b 和类胡萝卜素含量均逐渐下降,但叶绿素a/b 变化幅度小。在B 处理下,叶绿素a、b、a+b 含量与RB 处理变化趋势相同,但类胡萝卜素含量和叶绿素a/b随着盐胁迫程度的增加均呈先下降后上升的趋势。在相同盐胁迫下,与对应的RB 相比,B1、B2、B3 的叶绿素a、b、a+b、类胡萝卜素含量均降低,但叶绿素a 含量降幅小,而叶绿素b 含量降幅大,B1、B2、B3 的叶绿素a/b 均显著高于对应的RB 处理。可见,相对于RB 处理,B 处理对盐胁迫下番茄幼苗叶绿素及类胡萝卜素含量起到抑制作用,但有利于叶绿素a/b 值的提高,其中叶绿素a 含量和类胡萝卜素含量对光质不敏感,而叶绿素b 含量对光质敏感。
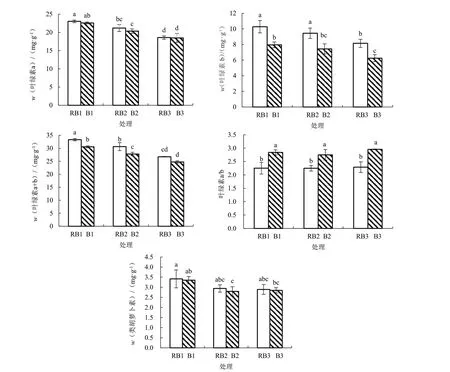
图4 不同光质和盐胁迫下番茄叶绿素和类胡萝卜素含量变化情况
2.4.2 不同光质和盐胁迫下番茄幼苗抗氧化酶活性变化 由图5 可知,在RB 处理下,随着盐胁迫程度的增加,POD 和APX 酶活性均逐渐下降,SOD酶活性呈先下降后上升的趋势;在B 处理下,SOD酶活性变化趋势与RB 处理相同,而POD 酶活性随着盐胁迫程度的增加呈先上升后下降的趋势,APX酶活性表现与RB 处理相反。在相同盐胁迫下,与对应的RB 处理相比,B1、B2、B3 处理的SOD 酶活性均降低,仅B3 处理显著低于RB3 处理,降幅为12.5%;B 处理POD 酶活性均显著降低,降幅分别为44.2%、20.4%、23.0%;APX 酶活性在B1 处理显著降低79.1%,但在B2 和B3 处理分别显著增加73.6%和691.6%。在3 种酶活性中,POD、APX 酶活性对光质尤为敏感;相对于RB 处理,B 处理在盐胁迫下抑制了番茄幼苗SOD 和POD 酶活性,但显著提高了APX 酶活性。
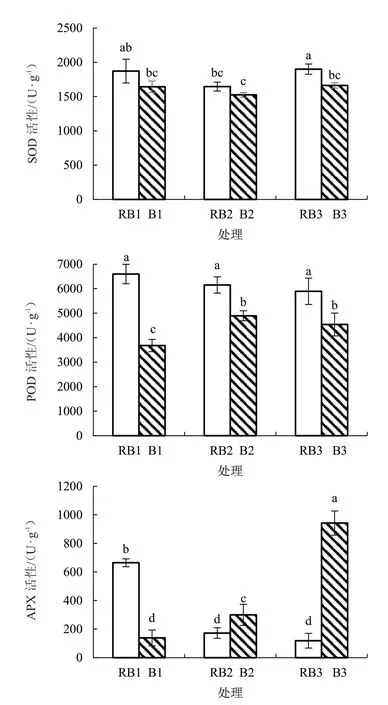
图5 不同光质和盐胁迫下番茄SOD、POD、APX酶活性变化情况
3 讨论与结论
光是影响植物生长代谢最重要的环境因子之一,其对植物的光合作用、形态建成、根系发育、物质代谢及基因表达等均有显著影响。而盐胁迫制约着生态环境与农业生产的发展,对植物生长发育造成了负面影响[12]。笔者的试验结果表明,B 处理在无盐胁迫下番茄株高生长速率显著低于RB 处理,所以在蓝光中添加红光有利于株高增长[13]。在两种光质下,B 处理的番茄鲜/干质量在无盐胁迫下较RB 处理低,说明在无盐胁迫下抑制了番茄幼苗干物质积累,此时添加红光有利于干物质积累。在相同盐胁迫下,番茄幼苗的株高生长速率在低盐(50 mmol·L-1)、高盐(100 mmol·L-1)胁迫下无显著差别,而且其干物质积累在低盐(50 mmol·L-1)胁迫下差别不显著,说明盐胁迫降低了番茄对光质的敏感度。与RB 处理相比,B 处理高盐(100 mmol·L-1)胁迫下根系受到的抑制作用更强,说明在高盐(100 mmol·L-1)胁迫下缺少红光不利于根系生长[14]。在这2 种光质下,相同盐胁迫下的处理间叶绿素a 和类胡萝卜素含量差异不大,表现出对光质不敏感的特性,而相同盐胁迫下的处理间叶绿素b含量差异显著,RB 处理叶绿素b 含量显著高于B处理[15],对光质尤为敏感。与B 处理相比,在各个盐浓度下,RB 处理都有较低的叶绿素a/b 比值,而生长在B 处理下的叶片叶绿素含量低于RB 处理,但叶绿素a/b 比值较高,这与吕喆等[16]对轮叶黑藻组织的研究一致。在不同光质处理下,酶活性表现出一定的差异性,而且POD、APX 酶活性对光质尤为敏感。在相同盐胁迫下,RB 处理的SOD、POD酶活性均高于B 处理,这与前人研究一致[17-18],说明缺少红光不利于SOD 和POD 酶活性的提高;在无盐胁迫下,RB 处理的APX 酶活性高于B 处理[19],但随着盐胁迫程度的增加,B 处理的番茄幼苗叶片中APX 抗氧化酶活性显著升高,与RB 处理表现出相反趋势,说明蓝光有利于提高盐(100 mmol·L-1)胁迫下番茄幼苗APX 酶活性,可增强盐胁迫下番茄幼苗叶片的抗氧化能力,这与邵久之等[20]对黄瓜幼苗酶活性的研究一致。
(1)在无盐胁迫下,蓝光较对照处理不利于番茄地上部生长,但对根系生长影响不显著,此外显著降低了叶片叶绿素b 含量以及POD 和APX 酶活性。
(2)在盐胁迫下,番茄幼苗的叶绿素b 含量、POD 和APX 酶活性对光质敏感,而株高生长速率、干质量、根冠比、叶绿素a 含量、类胡萝卜素含量等对光质不敏感。纯蓝光较对照处理显著降低了番茄叶片的叶绿素b 含量、抑制了SOD 和POD 酶活性,但显著提高了叶片的APX 酶活性,从而增强番茄幼苗自身清除H2O2的能力和减轻H2O2在盐胁迫下光合电子传递链和酶促代谢中的累积和毒害。
综上所述,盐胁迫总的来说降低了番茄对红蓝光质的敏感程度,但盐分胁迫下蓝光可能通过大幅提高叶片APX 酶活性增强番茄幼苗清除H2O2的能力,同时也需要增补部分红光来促进幼苗根、茎、叶的生长发育。
猜你喜欢
四川林业科技(2022年1期)2022-03-22
上海农业学报(2022年1期)2022-03-08
农村百事通(2019年17期)2019-10-08
家庭影院技术(2019年8期)2019-08-27
家庭影院技术(2019年1期)2019-01-21
家庭影院技术(2018年10期)2018-11-02
浙江农业学报(2017年1期)2017-05-17
现代农业科技(2017年1期)2017-03-06
江苏农业科学(2016年8期)2017-02-15
江苏农业科学(2016年8期)2017-02-15
