
新疆塔里木河叶尔羌高原鳅群体遗传学研究
2023-07-22 01:33赵文浩易少奎沈建忠李大鹏周小云
水产科学 2023年4期
赵文浩,易少奎,周 琼,沈建忠,李大鹏,周小云
( 1.华中农业大学 水产学院,农业农村部淡水生物繁育重点实验室,湖北 武汉 430070; 2.湖州师范学院 生命科学学院,浙江 湖州 313000 )
叶尔羌高原鳅(Triplophysayarkandensis)属鲤形目鳅科条鳅亚科高原鳅属,是新疆南部塔里木河流域的特有鱼类。叶尔羌高原鳅个体大、生长快、广泛分布于各个水系,曾是塔里木河的主要经济鱼类之一[1]。然而,近几十年来,塔里木河干流水流量日益减少,盐碱化程度加剧,加上人类的高强度捕捞,鲤(Cyprinuscarpio)、鲫(Carassiusauratus)等外来鱼类的引入造成的生存空间、饵料生物竞争等问题[2],导致其种群数量急剧减少,即将成为继扁吻鱼(Aspiorhynchuslaticeps)和塔里木裂腹鱼(Schizothoraxbiddulphi)之后的又一濒危物种[1]。因此,开展叶尔羌高原鳅种群资源调查,从分子水平解析其遗传多样性现状、群体遗传结构特征和种群历史动态等信息,对其种质资源保护和科学开发利用具有重要的指导意义。
在众多分子标记中,线粒体基因和微卫星标记在水产动物的遗传多样性和种属进化关系研究中的应用最为广泛。线粒体COⅠ基因进化速率适中,其5′端500~700 bp序列具有保守性强、碱基变异丰富等特点,常应用于脊椎动物的物种鉴定、群体遗传学等研究中[3]。微卫星标记具有稳定性好、多态信息含量丰富、呈共显性遗传、结果重复性高等优势,常用于动植物的群体遗传多样性评价、遗传连锁图谱构建和亲缘关系鉴定等方面[4]。Tsoi等[5]用线粒体DNA(COⅠ和16S rRNA)和微卫星标记研究西太平洋日本囊对虾(Penaeusjaponicus),发现其可以分为2个变种,并且均有很高的遗传多样性;袁希平等[6-7]用COⅠ和微卫星标记分别对铜鱼(Coreiusheterodon)和文蛤(Meretrixmeretrix)进行了遗传多样性和群体遗传变异研究。这些结果均为水产动物种质资源的保护、利用以及制定相应的保护策略提供了重要的基础数据。
目前,关于叶尔羌高原鳅遗传多样性和遗传结构的研究较少,仅见王锦秀等[8]用微卫星标记对塔里木河5个群体的研究报道。为了更多地了解叶尔羌高原鳅的种质资源现状,笔者从叶尔羌高原鳅的转录组测序数据中筛选到10个多态性高、稳定性好的四碱基重复微卫星标记,利用这些微卫星标记结合线粒体COⅠ基因对塔里木河两条支流(渭干河和车尔臣河)的8个群体进行群体遗传学分析,以期为叶尔羌高原鳅的资源保护及科学开发利用提供数据基础。
1 材料与方法
1.1 样品采集及DNA提取
试验所用叶尔羌高原鳅为2019年5—10月从塔里木河支流渭干河和车尔臣河采集,8个群体共采集到174尾样本(图1)。根据形态学特征[9]确认物种后,取鳍条于100%乙醇中暂存,运回实验室后置于冰箱-80 ℃保存备用。
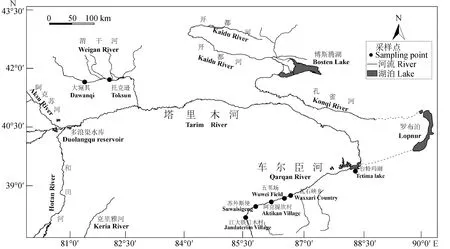
图1 塔里木河流域及叶尔羌高原鳅采样位点Fig.1 The Tarim River Basin and sampling sites of T. yarkandensis
用醋酸铵/异戊醇法提取基因组DNA。DNA质量和质量浓度分别用1%的琼脂糖凝胶电泳和NanoDrop 2000紫外分光光度计(Thermo Scientific, 美国)检测。将合格的DNA稀释至50 ng/μL用于后续试验。
1.2 线粒体COⅠ序列扩增及群体遗传学分析
从叶尔羌高原鳅的线粒体基因组(GenBank: KT224367.1)中查找COⅠ序列,用Premier 5.0设计引物(F:5′-GGCGGATTTGGAAACTGGC-3′,R:5′-ATCGGAATACCGTCGTGGC-3′),产物长度约1100 bp。PCR体系10 μL:DNA模板1 μL,上、下游引物各0.25 μL(10 μmol/L),2×PCR Master Mix 5 μL,双蒸水3.5 μL。PCR程序为:94 ℃预变性5 min,35个循环(94 ℃变性30 s,56 ℃退火30 s,72 ℃延伸50 s),72 ℃延伸10 min。PCR产物经1%琼脂糖凝胶电泳检测为单一、明亮且符合预期大小的条带后,送武汉擎科生物科技有限公司测序。
测序结果用DNASTAR[10]拼接并校对,用DnaSP 5.1[11]统计核苷酸多样性指数、单倍型数目和单倍型多样性指数;用Arlequin 3.5[12]进行分子方差分析,研究群体遗传变异来源及群体间遗传分化指数;用MEGA 7.0[13]基于K2P模型统计群体间遗传距离;用Barrier[14]基于群体间地理距离与遗传分化指数研究群体间潜在遗传障碍;用PopART[15]构建单倍型网络结构图。
1.3 微卫星位点筛选及群体遗传学分析
1.3.1 微卫星引物开发
用MISA软件在叶尔羌高原鳅转录组(DOI:10.6084/m9.figshare.16989073)中查找所有微卫星位点,筛选重复单元为四至六碱基、且四碱基和五碱基重复5次以上、六碱基重复4次以上、产物在300 bp以内的位点作为候选微卫星标记,用Premier 5.0设计引物。
1.3.2 多态性微卫星位点筛选
随机选取8个叶尔羌高原鳅鳍条DNA,用上述引物扩增,PCR体系及程序同1.2,产物用1%琼脂糖凝胶电泳检测,初步选出能稳定扩增出条带的引物,再用8%非变性聚丙烯酰胺凝胶电泳进一步筛选出产物在样本间有多态性的引物。
对筛选得到的多态性引物,在其上游引物的5′端用FAM、Hex等荧光基团进行修饰得到荧光引物,用该上游引物和相应的下游引物进行扩增,产物送武汉天一辉远生物公司进行毛细管电泳验证微卫星位点多态性。
1.3.3 基因分型及数据分析
用筛选得到的多态性微卫星位点进行群体遗传学分析。PCR体系及程序同1.2,产物在ABI 3730XL基因分析仪上分型,原始数据用GeneMarker[16]分析得出峰图等数据文件,人工校读并统计数据。用PopGene 3.2[17]统计等位基因数、有效等位基因数、观测杂合度、期望杂合度、基因流、香农-维纳多样性指数、F-统计量、群体间根井正利遗传距离等。用Mega 7.0[13]分析群体间的根井正利遗传距离,用非加权组平均法构建系统进化树;用Cervus 3.0.7[18]计算多态信息含量;用Migrate-n 3.6.11[19]统计各群体的历史迁移率;用Structure 2.3.4[20]对群体遗传结构进行分析,将运算结果提交至在线软件Structure harvester[21]推算最佳K值,然后用CLUMPP[22]与Distruct[23]对结果进行可视化。
2 结 果
2.1 基于线粒体COⅠ序列的分析结果
测序结果经拼接及校对后,选取968 bp的COⅠ序列用于后续分析。变异位点统计结果显示,多态性位点有30个,其中简约信息位点24个,单一突变位点6个。在8个群体的174个样本中共检测到22个单倍型,平均单倍型多样性与核苷酸多样性分别为0.8829和0.0032。此外,与车尔臣河群体相比,渭干河各群体遗传多样性水平普遍较低(表1)。
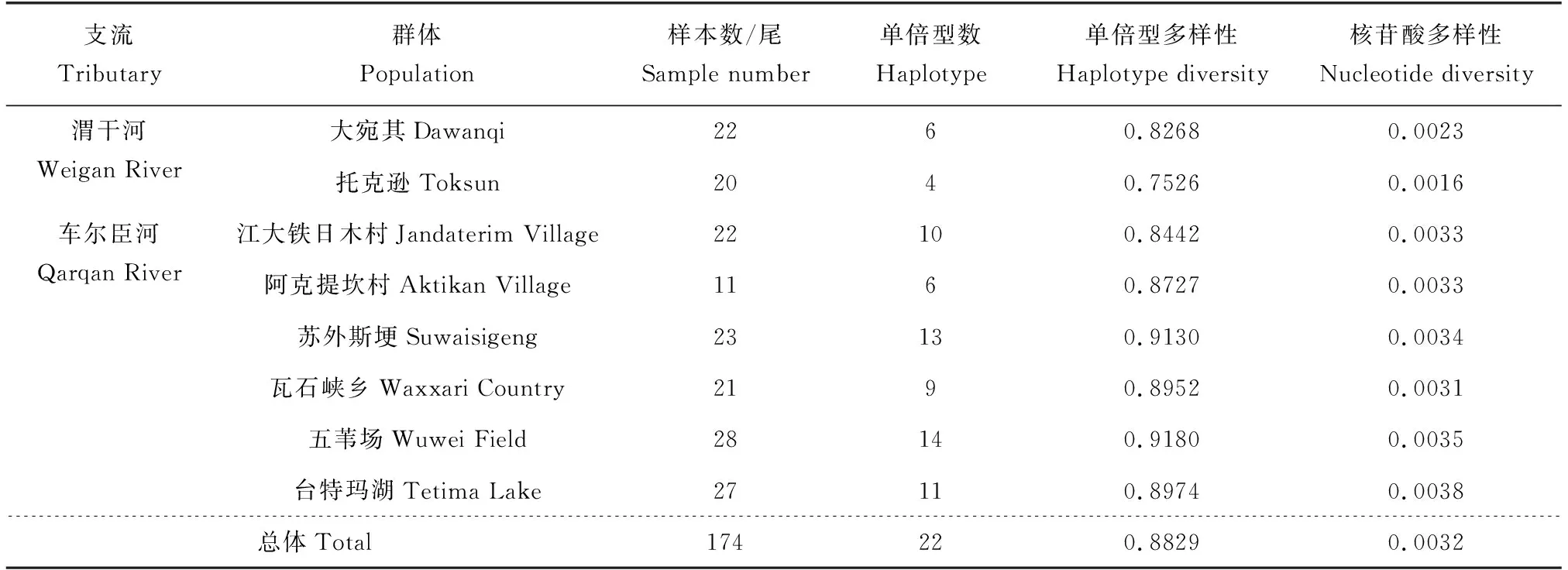
表1 基于线粒体COⅠ基因的叶尔羌高原鳅各群体遗传多样性参数Tab.1 Genetic diversity parameters of T. yarkandensis in each population based on COⅠ gene
各群体间遗传距离为0.0019~0.0041,远低于不同种或亚种的区分阈值(0.06);大部分群体间遗传分化指数小于0.05。此外,同一支流群体间遗传分化指数均小于不同支流群体间的遗传分化指数,表明两支流群体间存在遗传变异。为进一步探讨变异来源,将8个群体按支流分为两个亚群进行分子方差分析,结果显示,群体内的遗传变异占总变异的95.12%,支流间与支流内群体间分别仅占总变异的4.00%和0.89%,表明遗传变异主要来源于群体内部,并且,渭干河群体和车尔臣河群体间存在一定的遗传分化(表2)。
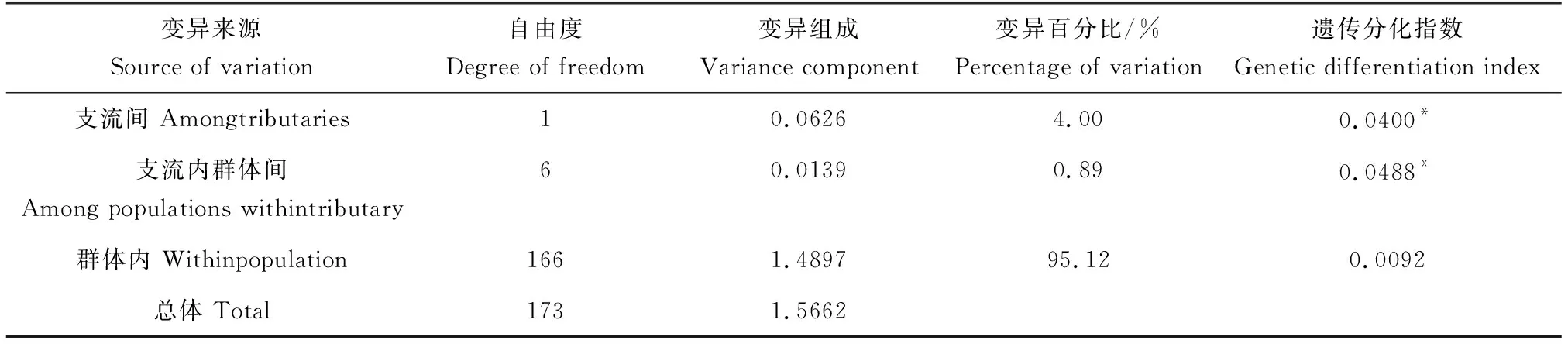
表2 基于分子方差分析的叶尔羌高原鳅群体间遗传变异Tab.2 Genetic variation between populations of T. yarkandensis based on AMOVA
基于样品构建的系统发育树(图2a)和基于单倍型构建的网络结构图(图2b)均显示,渭干河的群体(托克逊与大宛其)和车尔臣河群体(阿克提坎村、江大铁日木村、苏外斯埂、台特玛湖、瓦石峡乡和五苇场)的样本交错分布于各个遗传分支,未能形成明显的地理聚群和谱系结构。此外,单倍型网络图显示,Hap-6与Hap-2为8个群体共享单倍型,但Hap-6与其他单倍型之间联系更为紧密,因此推测其为古老单倍型;特有单倍型仅出现在车尔臣河3个群体中(五苇场、苏外斯埂和江大铁日木村);渭干河群体(托克逊和大宛其)与其他群体间均存在共享单倍型。对各群体进行Barrier分析发现,渭干河群体与车尔臣河群体之间存在潜在遗传障碍(图3)。
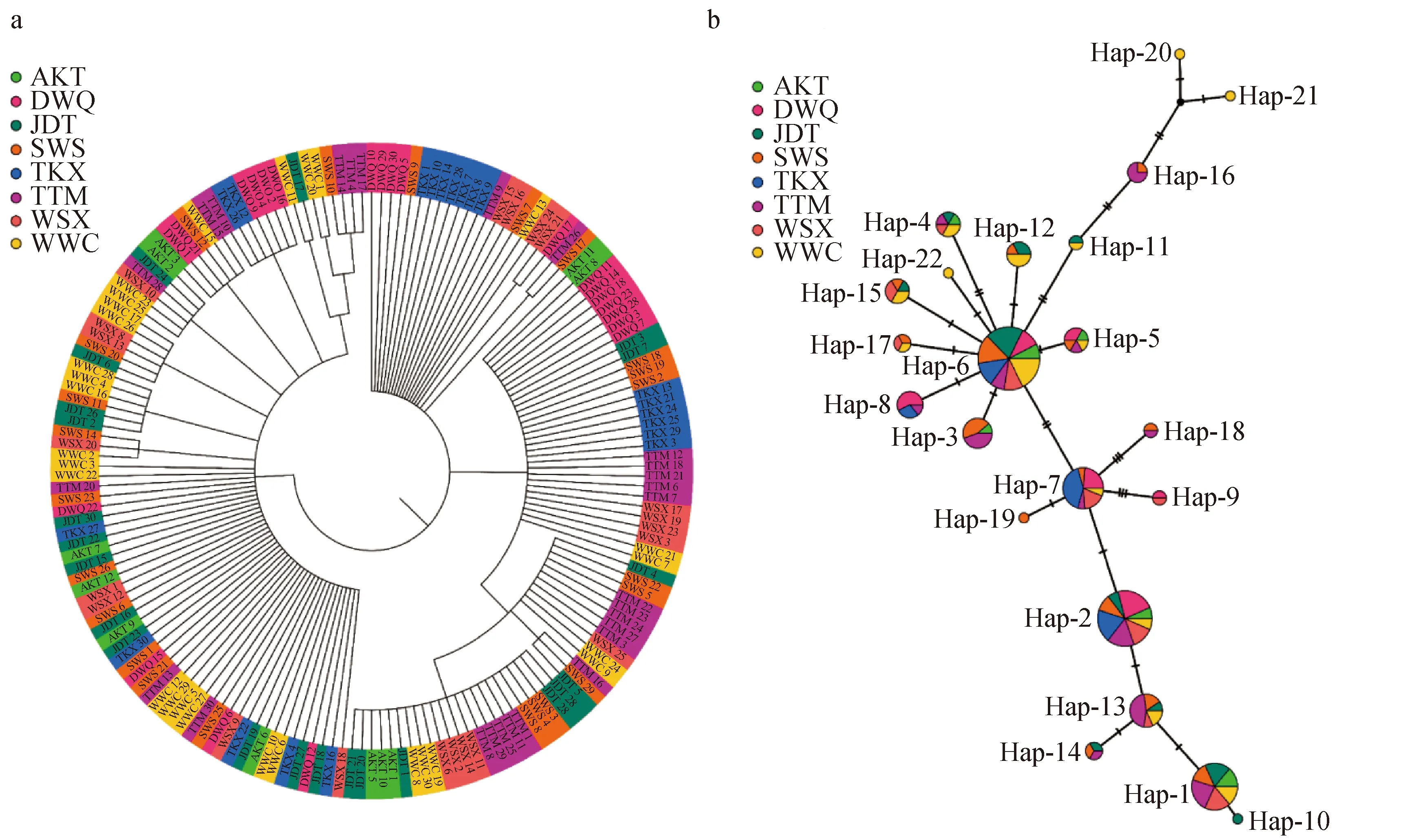
图2 基于样品构建的系统发育树(a)和基于单倍型构建的网络结构(b)Fig.2 NJ phylogenetic trees (a) and median-joining network (b) constructed based on samples and haplotype, respectivelyDWQ、TKX、JDT、AKT、SWS、WSX、WWC、TTM分别代表该群体采自大宛其、托克逊、江大铁日木村、阿克提坎村、苏外斯埂、瓦石峡乡、五苇场和台特玛湖;下同.DWQ, TKX, JDT, AKT, SWS, WSX, WWC and TTMmeans the samples were from Dawanqi, Toksun, Jandaterim Village, Aktikan Village, Suwaisigeng, Waxxari Country, Wuwei Field and Tetima Lake, respectively; et sequentia.
2.2 基于微卫星标记的分析结果
经1%琼脂糖凝胶电泳初筛、8%非变性聚丙烯酰胺凝胶电泳复选、毛细管电泳进一步验证,最终筛选得到10个多态性微卫星位点(表3),各位点的多态信息含量值均大于0.5,表明各位点均具有较高的多态性[24],可用于后续群体遗传学分析。
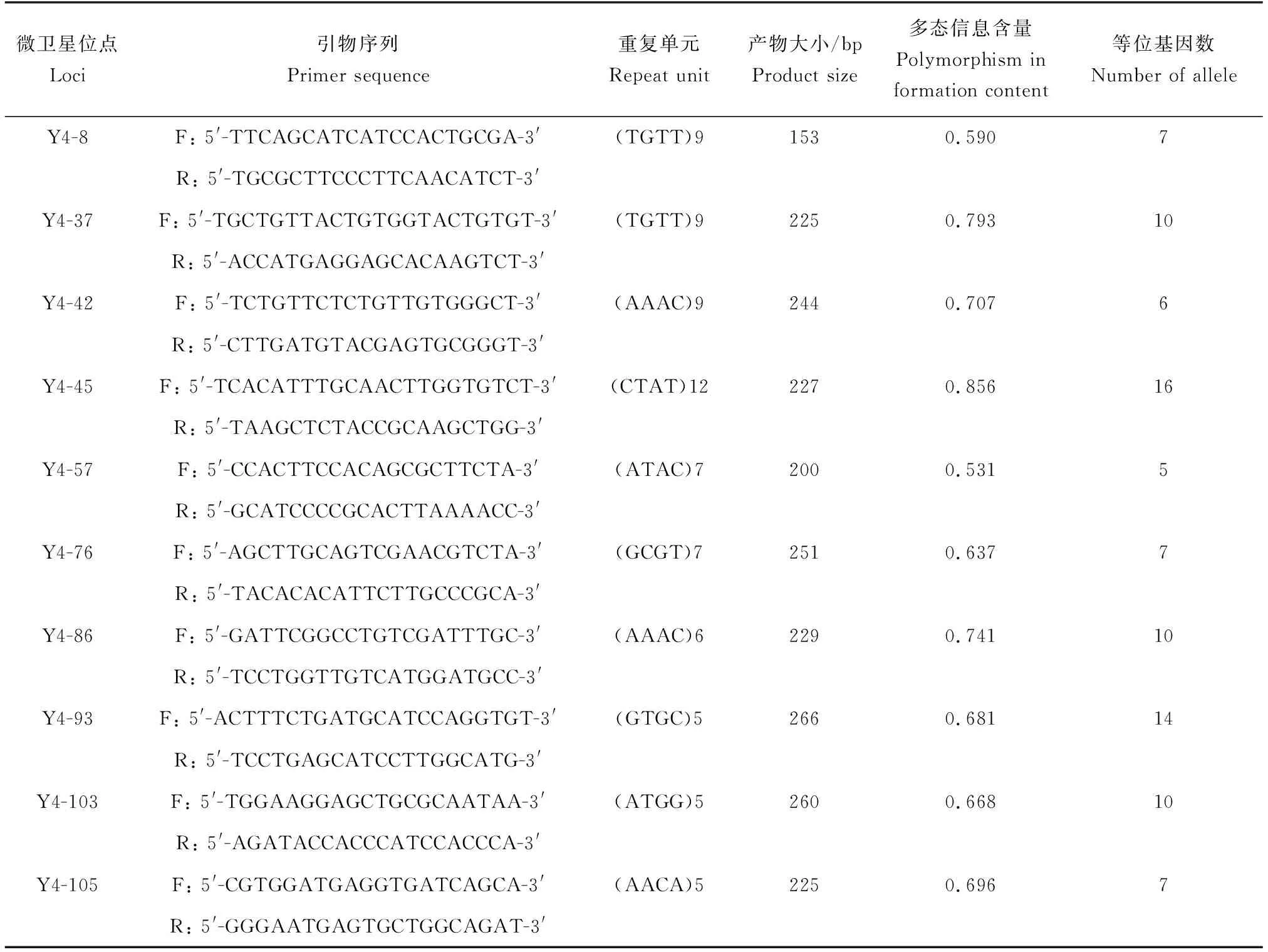
表3 多态性微卫星位点及其引物信息Tab.3 Polymorphic microsatellite loci and their primer information
用10个微卫星标记对各群体的样本进行分析,结果显示,台特玛湖群体的等位基因数最多为6.500(表4)。各群体观测杂合度和期望杂合度分别为0.607~0.736和0.655~0.755,平均多态信息含量为0.629~0.696,香农-维纳多样性指数为1.300~2.249。托克逊群体平均香农-维纳多样性指数最高(1.540)。

表4 8个叶尔羌高原鳅群体遗传多样性参数Tab.4 Genetic diversity parameters of eight populations of T. yarkandensis
基于10个微卫星标记的分子方差分析结果显示,遗传变异主要来源于群体内部(97.23%);两支流群体间遗传分化与支流内群体间遗传分化贡献程度很低,仅为2.20%与0.57%;同时,两支流群体间也存在一定的遗传分化(表5)。
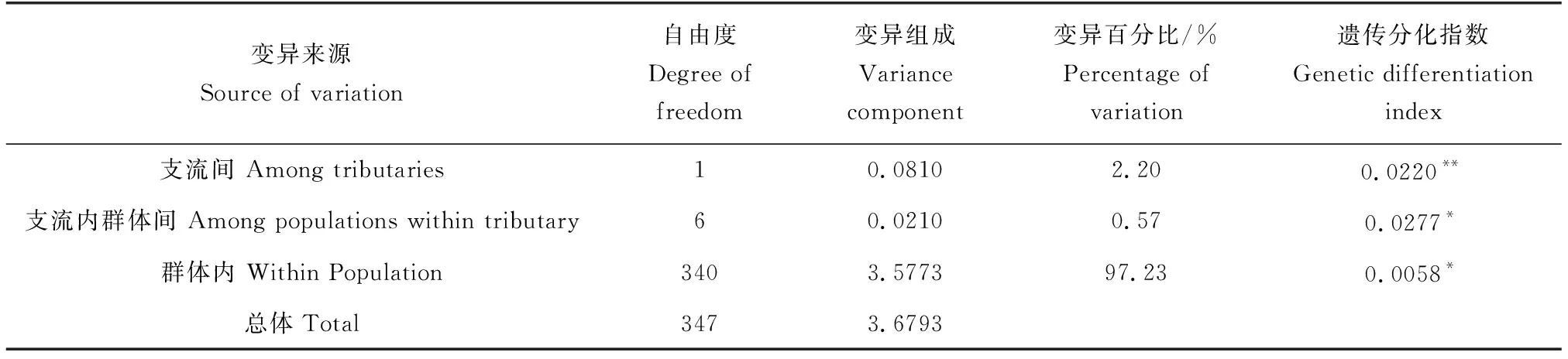
表5 基于分子方差分析的群体间遗传变异Tab.5 Genetic diversity parameters between populations based on AMOVA
各位点的F-统计量和基因流结果显示,群体内近交系数与整体近交系数分别为0.0331和0.0669。群体间的基因流为4.3027~9.5282,均大于1,加之各群体遗传分化指数为0.0248~0.0549,表明各群体间存在一定的基因交流,遗传分化程度较弱。比较各群体历史迁移率发现,阿克提坎村、江大铁日木村与托克逊群体主要为迁入,其他群体主要为迁出,其中托克逊群体迁入率(14.33)远大于迁出率(8.52),而车尔臣河台特玛湖群体的迁出率(14.80)远大于迁入率(10.11)(表6)。
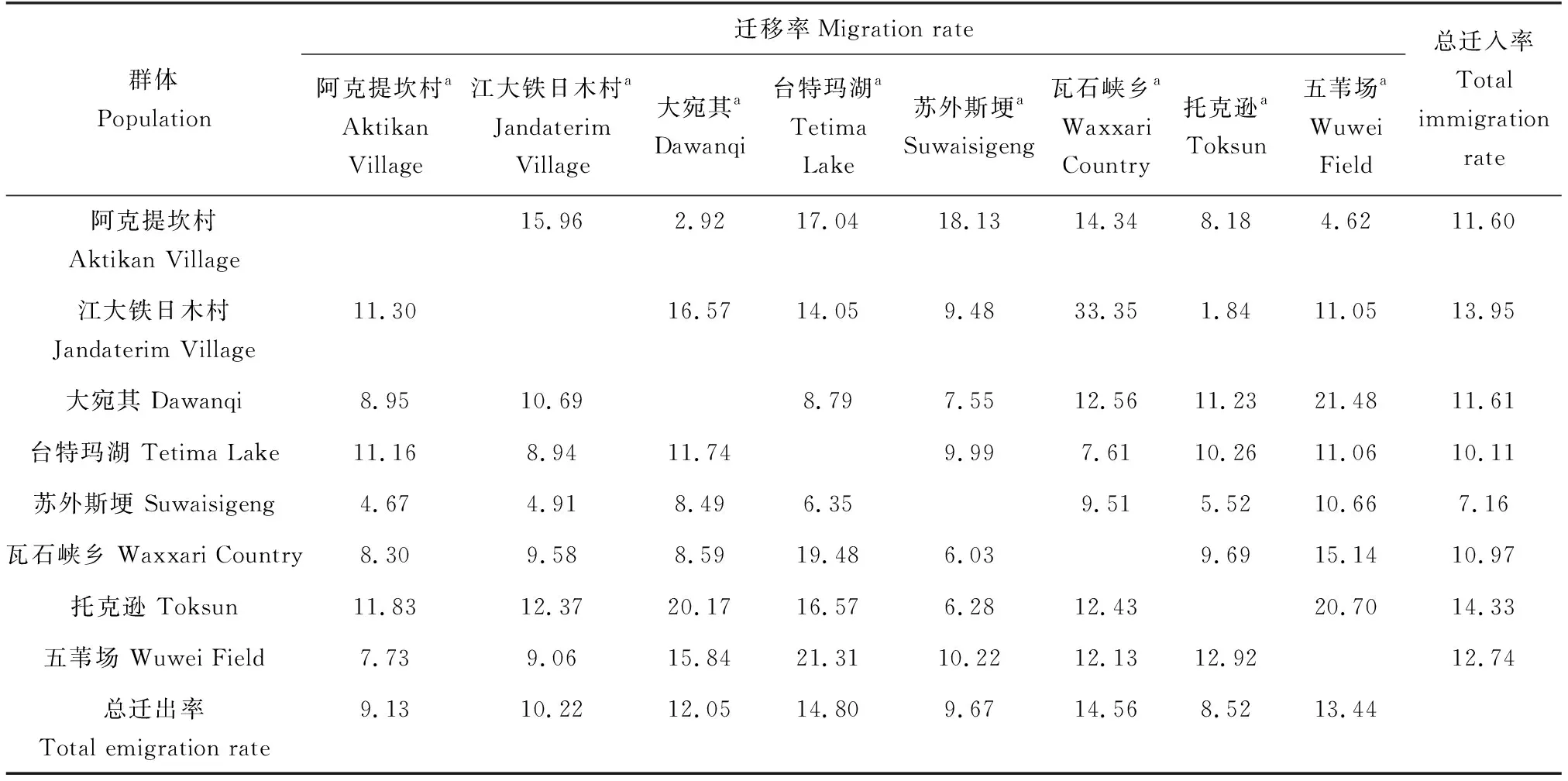
表6 叶尔羌高原鳅各群体迁移率Tab.6 The migration rate of each T. yarkandensis populations
各群体间根井正利遗传距离为0.0389~0.2090,其中瓦石峡乡与大宛其两群体间遗传距离最远(0.2090),但并未达到种的分类标准(<0.3)。群体间遗传一致度为0.8114~0.9618,表明各群体有较高的遗传同质性。以根井正利遗传距离为基础,非加权组平均法构建的系统进化树显示,8个群体分为2支,渭干河2个群体(托克逊和大宛其)和车尔臣河群体(阿克提坎村、瓦石峡乡、苏外斯埂、江大铁日木村、台特玛湖和五苇场)分别聚为一支(图4a)。
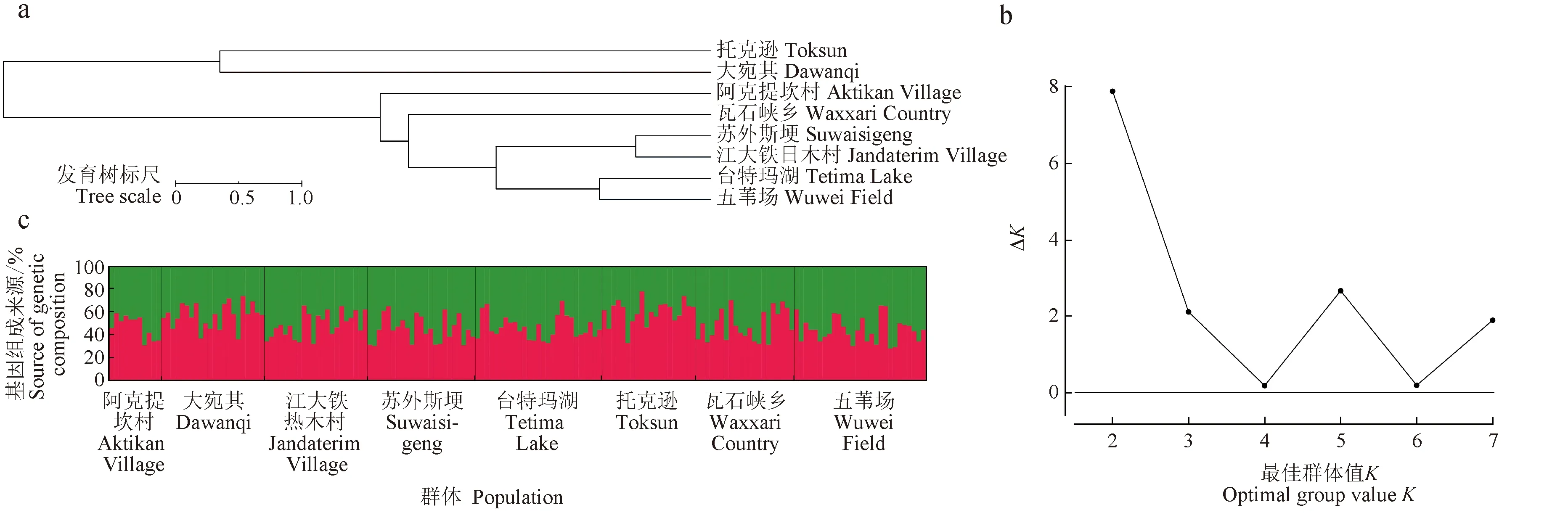
图4 叶尔羌高原鳅各群体间的非加权组平均系统发育树(a)、基于ln P (D)模型为准则的K值结构分析(b)和Structure结构分析(c)Fig.4 UPGMA phylogenetic tree (a), model choice criterion ln P (D) of the Structure analysis for each K value (b) and structure analysis (c) of the T. yarkandensis populations
ΔK计算显示,在最适状态下存在2个组群(K=2)(图4b)。当K=2时,各群体并无明显的聚类情况(图4c)。对各群体基因组成所占比例分析发现,渭干河群体基因组成主要来源于组群1(红色),而车尔臣河群体基因组成主要来源于组群2(绿色),表明两支流的群体间存在一定的遗传分化。
3 讨 论
3.1 叶尔羌高原鳅微卫星标记开发
本研究中,基于转录组测序数据筛选到10个四碱基重复微卫星标记。与二碱基重复微卫星标记相比,四碱基重复微卫星标记不容易出现因PCR扩增链滑脱而产生的影子带,因此数据读取更方便、准确,此外,由于转录组序列是功能基因的表达片段,在其中发现的微卫星标记可能会与功能基因相关,并有可能与一些生产性状相关联[25]。因此,笔者开发的微卫星标记不仅可用于群体遗传学分析,而且在叶尔羌高原鳅的分子标记辅助选择育种等研究中也有较高的潜在价值。
3.2 群体遗传多样性
物种的遗传多样性水平与其环境适应能力和进化潜能密切相关,一个物种的遗传多样性水平越高,遗传变异越丰富,其对环境的适应能力就越强,就越容易扩展其分布范围和开拓新的环境[26]。线粒体DNA的单倍型多样性和核苷酸多样性是衡量物种遗传多样性的重要参数,Grant等[27]将单倍型多样性指数0.5、核苷酸多样性指数0.05作为分界点。本研究中,各群体的平均单倍型多样性指数为0.8829,平均核苷酸多样性指数为0.0032,为高单倍型多样性和低核苷酸多样性。这种“高单倍型多样性、低核苷酸多样性”现象在鱼类中比较常见,已有研究显示,其是由于物种受到瓶颈效应后种群数量迅速扩张导致的,即种群单倍型多样性积累很快,但核苷酸变异还未能积累[28-29]。叶尔羌高原鳅曾是塔里木河的优势种群,但由于引水灌溉等因素,20世纪70年代,罗布泊、台特玛湖曾一度干涸,塔里木河下游长时间断流,导致叶尔羌高原鳅、塔里木裂腹鱼等土著鱼类资源量大幅度下降。自2000年起,塔里木河下游启动了生态输水工程,极大地缓解了塔里木河下游土著鱼类生态环境严重退化的趋势,这可能是塔里木河叶尔羌高原鳅曾遭受瓶颈效应出现“高单倍型多样性、低核苷酸多样性”特征的原因。
基于微卫星标记的多态信息含量、观测杂合度和期望杂合度也是评价群体遗传多样性的重要指标[24]。笔者利用新开发的10个微卫星标记对各群体进行的遗传多样性分析发现:8个群体的多态信息含量为0.629~0.696,均大于0.05,表明多态性水平较高;各群体观测杂合度和期望杂合度分别为0.607~0.736和0.655~0.755,均大于基于13种淡水鱼类统计得出的平均杂合度0.46[30],因此,各叶尔羌高原鳅群体具有较高的遗传多样性水平。这与王锦秀等[8]用微卫星标记对5个叶尔羌高原鳅群体的研究结果一致。
此外,笔者发现基于线粒体COⅠ序列分析的渭干河群体核苷酸多样性和单倍型多样性均低于车尔臣河群体,表明渭干河群体的遗传多样性水平低于车尔臣河。王锦秀等[8]的研究也发现,塔里木河下游群体(如车尔臣河群体、台特玛湖群体)的遗传多样性水平高于中上游群体(如阿克苏河群体、台南河群体),并认为造成这种现象的原因可能是上游叶尔羌高原鳅容易随水流到达下游,促进了基因交流,使得下游群体具有较多的等位基因数和较高的遗传多样性水平。
3.3 群体结构和遗传分化
遗传分化指数是衡量群体间遗传分化的重要指标。当遗传分化指数<0.05时为极小遗传分化,在>0.05~0.15时为中度遗传分化,在>0.15~0.25时为较大遗传分化,在>0.25时则为极大遗传分化[31]。本研究基于线粒体COⅠ序列和微卫星标记分析的结果均显示,各群体间的遗传分化指数均小于0.05或位于0.05附近,因此属于低分化水平,并且,遗传变异的主要来源为群体内部,群体间的遗传结构差异不大,相似度较高。推测造成上述结果的原因是群体间存在一定的基因交流。基因流是基因在群体间的流动,Slatkin[32]认为:基因流大于1时能发挥均质化的作用,即能有效抑制种群间的分化;基因流小于1时会促进群体发生遗传分化。本研究基于微卫星标记的群体间基因流值均大于1,证实各群体间有一定的基因交流,这与基于遗传分化指数的分析结果一致。
分析各群体的迁入率和迁出率参数发现,台特玛湖群体的迁出率(14.80)最高,且远大于迁入率(10.11)。笔者推测,台特玛湖群体为塔里木河中下游的扩散中心,扩散方向为渭干河和车尔臣河上游,其生态学依据在于:(1)台特玛湖是塔里木河的尾闾湖泊,1972年以来曾一度干涸,为防止干流断流,自2000年起,塔里木河下游启动了生态输水工程[33],这可能促进了台特玛湖叶尔羌高原鳅向各支流的扩散;(2)叶尔羌高原鳅有溯河洄游的繁殖习性,每年4月中旬至7、8月,大批叶尔羌高原鳅溯河洄游至塔河上游,寻找适宜产卵场[34],这可能是台特玛湖群体向各支流中上游扩散的另一重要原因。此外,这种扩散也可能是群体间基因流处于较高水平(4.3027~9.5282)的原因之一,其有效阻止了群体内遗传变异的减少,降低了群体间的遗传分化。
基于线粒体COⅠ序列和微卫星标记的分析结果均表明,两支流群体间的遗传变异(线粒体COⅠ,4.00%;微卫星,2.20%)远高于支流内群体间的遗传变异(线粒体COⅠ,0.89%;微卫星,0.57%);基于COⅠ序列的群体间Barrier分析显示,两支流群体间存在潜在遗传障碍;基于微卫星标记使用非加权组平均法构建的系统发育树显示,渭干河群体和车尔臣河群体位于不同的遗传分支;并且群体Structure聚类结果显示,渭干河与车尔臣河群体的基因组成有明显差异。这些结果均表明,渭干河群体与车尔臣河群体间存在一定的遗传分化。由于鱼类的种群遗传关系和结构与地理隔离、水系分布等密切相关[35],渭干河位于塔里木河的中游,而车尔臣河位于塔里木河的下游,两支流的地理位置相对较远,并且,随着塔里木河流域水资源稀缺加剧,其流域源下游和干流中下游经常出现季节性断流[36],导致叶尔羌高原鳅生境片段化问题。这些可能影响了两支流群体间的基因交流,使得两支流群体间遗传分化程度变大,进而产生差异。王锦秀等[8]也发现,塔河各支流的叶尔羌高原鳅之间存在一定的遗传分化,且遗传距离与群体地理位置的分布相吻合。
此外,采用COⅠ序列分析时,各群体未能形成明显的地理聚类和谱系结构,这与采用微卫星标记分析的结果明显不同,对于这种差异,笔者推测可能是由线粒体COⅠ序列变异度相对较小,与微卫星标记相比分辨率较差造成的[37]。并且,笔者基于微卫星标记得到的群体间遗传距离、群体聚类以及支流群体间的遗传分化结果与群体的实际地理分布更吻合,因此笔者认为用微卫星标记分析的结果更准确。
4 结 论
本研究中,笔者结合微卫星标记和线粒体COⅠ序列的分析结果发现,叶尔羌高原鳅各群体的遗传多样性水平较高,但遗传变异主要来源于群体内部,群体间遗传分化程度较小,其进化史上曾遭受明显的瓶颈效应。两支流群体间存在潜在的遗传障碍,并发生了一定的遗传分化,这可能是由于塔里木河中下游经常发生季节性断流,造成叶尔羌高原鳅生境片段化,进而降低了支流间的基因交流水平。因此,有必要采取有效的措施对叶尔羌高原鳅的栖息环境进行保护,如控制上游荒地开发,尤其是控制高耗水作物种植面积的扩大,防止上游供水量急剧减少,严格执行每年1次的下游生态补水工程等,避免因生境片段化问题而导致的种群遗传多样性降低。
猜你喜欢
特产研究(2022年6期)2023-01-17
家教世界·创新阅读(2020年4期)2020-06-03
当代工人·精品C(2020年1期)2020-05-20
家教世界·创新阅读(2020年1期)2020-05-11
家教世界·创新阅读(2020年2期)2020-04-07
意林绘阅读(2019年2期)2019-03-05
下一代英才(2018年12期)2018-02-20
四川动物(2017年4期)2017-07-31
集美大学学报(自然科学版)(2015年4期)2015-02-28
河北遥感(2014年3期)2014-07-10
