
冷季补饲对泌乳期藏羊母羊乳汁成分与代谢产物的影响
2023-07-14 01:41侯生珍桂林生罗增海
安徽农业大学学报 2023年3期
周 力,侯生珍,桂林生*,罗增海
冷季补饲对泌乳期藏羊母羊乳汁成分与代谢产物的影响
周 力1,3,侯生珍1,桂林生1*,罗增海2*
(1. 青海大学农牧学院,西宁 810016;2. 青海省畜牧总站,西宁 810001;3 宁夏大学动物科技学院,银川 750021)
为了使用乳品快速分析仪和液相色谱-质谱联用(LC-MS)技术对放牧与补饲条件下藏羊母羊乳成分和代谢产物进行差异分析,选取胎次一致、体重相近(43.85 kg±7.06 kg)且发育良好的3岁龄藏羊母羊440只,随机分成两组,即自然放牧(control group,CG),放牧+补饲0.25 kg·d-1精料补充料(test group,TG)。试验期为60 d(配种后150~210 d)。试验结束时,从两组中各收集5只母羊的乳汁分析营养成分,同时检测其代谢产物。结果显示:(1)与CG组相比,TG组藏羊母羊的产奶量、脂肪、蛋白质、灰分及非脂固形物含量均有所提高,其中产奶量与脂肪含量差异显著(<0.05)。(2)两组共获得已注释显著性差异代谢物39种,其中24种上调,15种下调;结合KEGG通路分析,发现脂肪细胞脂解调节(regulation of lipolysis in adipocyte),含血清素神经突触(serotonergic synapse),醛固酮合成与分泌(aldosterone synthesis and secretion),FcεRI信号通路(Fc epsilon RI signaling pathway),不饱和脂肪酸生物合成(biosynthesis of unsaturated fatty acid),苯丙氨酸、酪氨酸和色氨酸生物合成(phenylalanine, tyrosine and tryptophan biosynthesis)是最为显著性关键代谢通路,并筛选出花生四烯酸(arachidonic acid),二十碳五烯酸(eicosapentaenoic acid)和D-果糖-1-磷酸(D-fructose-1-phosphate)作为差异显著相关代谢物。综上,通过冷季补饲提高了泌乳期藏羊母羊乳汁品质,并能改善其代谢产物,这为今后绵羊母乳的研究与开发提供一定理论依据与技术支持。
藏羊;精料补饲;代谢组;乳品质;泌乳期;脂肪酸
近年来,乳汁的营养价值逐渐走进人们的视野,已成为食品领域研究的热点[1]。羊乳含有丰富的蛋白质、脂肪、维生素以及矿物质等,已被公认其营养组成是最为接近母乳的乳产品[2]。据有关报道,羊乳中酪蛋白含量较少,易被机体消化吸收,所以更适合被婴幼儿、老年及消化不良的人群饮用[3]。同时乳中不仅能够提供幼畜生长发育和人们日常所需的养分,同时还能含有大量的生物活性成分。除此之外,在羊乳中还发现其他抗氧化成分,例如维生素C、维生素E以及超氧化物歧化酶等物质,能够清除机体内的自由基,延缓其衰老[4-5]。随着人们对羊乳营养价值的不断重视,羊乳及其相关产品的种类也越来越多,由此可见,未来羊乳将具有十分广阔的市场前景。
目前我国绵羊的饲养量、出栏量、肉产量、皮产量以及绒产量均居于世界前列[6]。而藏羊()作为青藏高原特有的绵羊品种资源,对低温、少氧以及强紫外辐射等恶劣环境具有较强的适应性,在羊产业中占有十分重要的地位[7]。但由于其所处环境无明显的四季分界,只有冷季(10月—翌年5月)和暖季(5月—10月)之分[8],加之冷季牧草的产量低且营养素含量也极度匮乏。因此,在寒冷季节对高原土著动物及时补饲不仅能够满足机体的营养需要,同时还有利于幼畜的快速生长。例如在冷季对牦牛进行“少量补饲”能够在一定程度上缓解其体重掉膘[9]。其他试验也进一步证实,冷季给予怀孕牦牛补饲精料可以提高犊牦牛的初生重[10]。
代谢组学是继基因组学、转录组学、蛋白质组学等之后,又一兴起的组学技术,能够从整体和多角度阐明机体的内源性代谢变化[11]。因其具有高分辨率、高灵敏度、高通量等优点而得到广泛应用,研究对象主要为相对分子质量<1 000的小分子代谢物[12]。代谢物作为机体生化反应的终点,直接体现生物功能的最终效应,能够表明机体异常的生理状态[13]。其技术主要包括核磁共振技术(NMR)、气相色谱-质谱联用技术(GC-MS)和液相色谱-质谱联用技术(LC-MS)等[14]。其中LC-MS因分离能力高、灵敏度强以及选择性好等优点已成为一种不可或缺的检测方式[15]。
因此,本研究以高海拔地区藏母羊为试验对象,探讨放牧与补饲条件下乳汁营养物质的差异,并筛选其差异代谢物,为绵羊母乳的加工和生产提供基本理论数据。
1 材料与方法
1.1 试验设计
试验选用胎次一致、体重相近(43.85 kg±7.06 kg)及发育健康的3岁龄藏母羊440只,每组220只,即对照组(control group, CG)和试验组(test group, TG)。对照组在青海天然牧场进行放牧,试验组在自然放牧基础上每天19 : 30归牧后每只母羊补饲精料补充料0.25 kg。试验于青海省海南州同德县河北乡夏知迈村进行,整个试验期为60 d(配种后150~210 d),基础饲粮设计参考按照《肉羊饲养标准(NY/T 816—2004)》[16]进行,其组成及营养水平见表1。
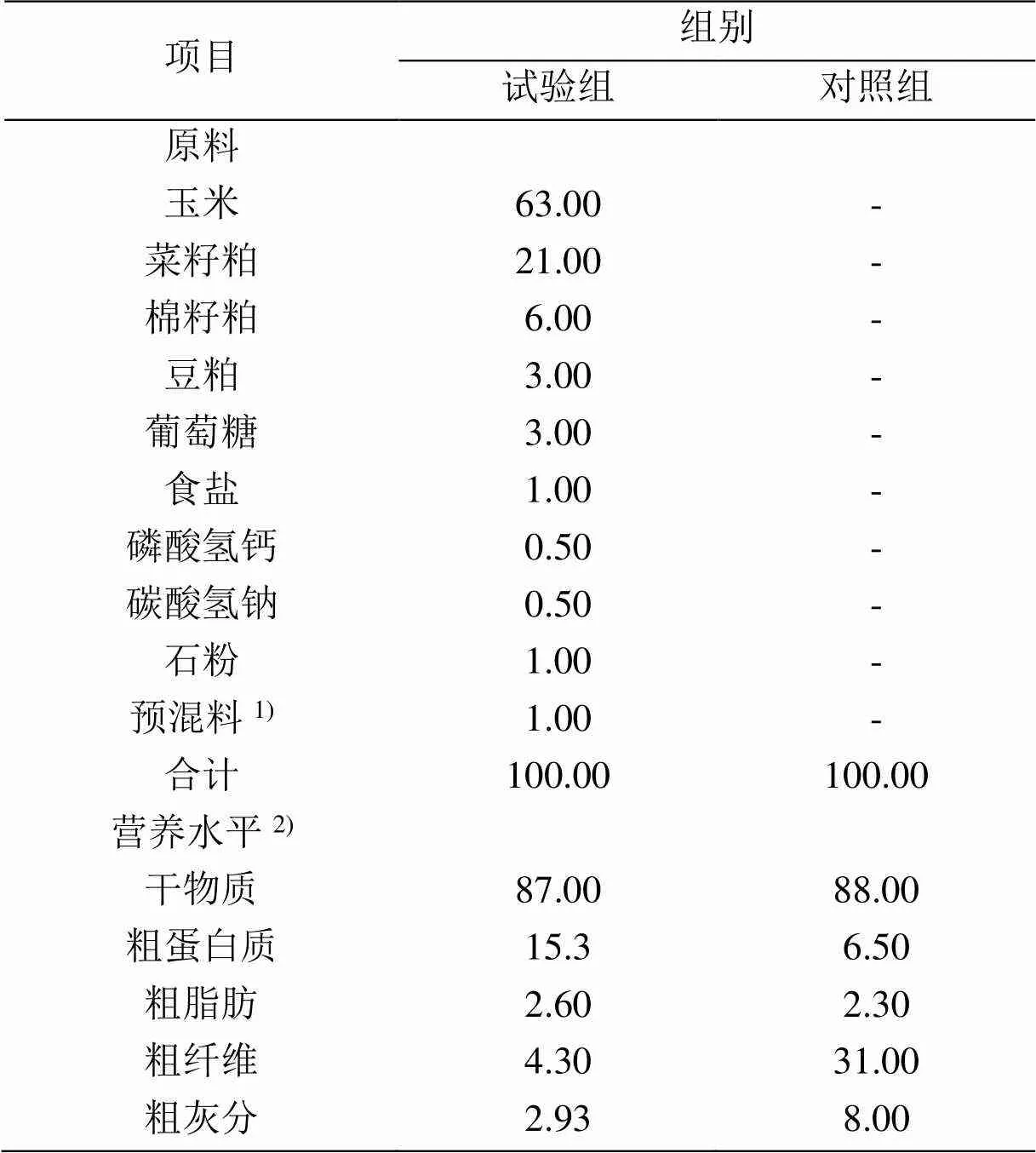
表1 基础饲粮组成和营养水平(干物质基础)
1)预混料为每千克日粮提供:铁 Fe 9.0 g,铜 Cu 1.8 g,锰Mn 6.0 g,锌Zn 10.0 g,碘I 100.0 mg,硒Se 25.0 mg,钴Co 25.0 mg。2)营养水平均为实测值。
1.2 乳样采集
母羊出生后第760 天,每日对羔羊进行哺乳3次(07:30、12:00和18:30),完成哺乳后对所有羔羊进行体重称量并与母羊隔离,当日泌乳量为3次测量之和,将每只母羊采集到的乳汁进行混合摇匀,用于乳品质分析以及进行非靶向代谢组学的测定。
1.3 试验方法
1.3.1 乳汁营养成分分析 采用乳品快速分析仪MASTER(YQ1-014,保加利亚)测定藏母羊乳中脂肪、蛋白质、灰分和非脂固形物含量。
1.3.2 代谢物提取 首先吸取100 μL乳汁,加入300 μL甲醇、20 μL内标,涡旋瞬时震动后,在冰水浴条件下超声提取5 min。在-20℃条件下静置2 h,于4℃条件下13 000 r·min-1离心15 min,再吸取200 μL上清液于2 mL进样瓶,进行液相色谱-质谱联用技术分析。
1.3.3 LC-MS检测 ① UPLC条件:样品盘温度为4 ℃,进样体积:1 μL,UPLC HSS T3(1.7 μm,2.1 mm×100 mm)液相色谱柱对样品化合物进行色谱分离,液相色谱A相为水相(负模式:5 mmol·L-1醋酸铵水溶液),B相均为乙腈。
②质谱条件:Q Exactive Orbitrap高分辨质谱仪,采用电喷雾电离源(ESI),负离子(-3 100 V)模式下进行一级、二级质谱数据的采集。毛细管温度:320℃;鞘气流速:45 Arb;辅助气流速:15 Arb;扫描范围:70~1 000 m·z-1;一级分辨率:70 000;二级分辨率:17 500;分步碰撞能量的强度取值:3;分步碰撞能量:20、40和60 eV;扫描速率:7 Hz。
1.4 数据处理分析
试验数据先用Excel 2010初步整理后,检验法进行两组间显著性分析,结果以“平均数±标准差”表示,<0.05作为显著性判断标准。
2 结果与分析
2.1 冷季补饲对泌乳期藏羊母羊乳成分的影响
藏羊母羊乳成分的结果见表2。TG组藏羊母羊的产奶量显著高于CG组(<0.05);与CG组相比,TG组藏羊母羊的脂肪、蛋白质、灰分和非脂固形物分别提高了15.29%、2.65%、5.56%和2.29%,其中脂肪含量差异显著(<0.05)。

表2 放牧与补饲下藏羊母羊乳成分差异比较
注:同列不同字母表示差异显著(>0.05),相同字母或无字母表示差异不显著(>0.05)。
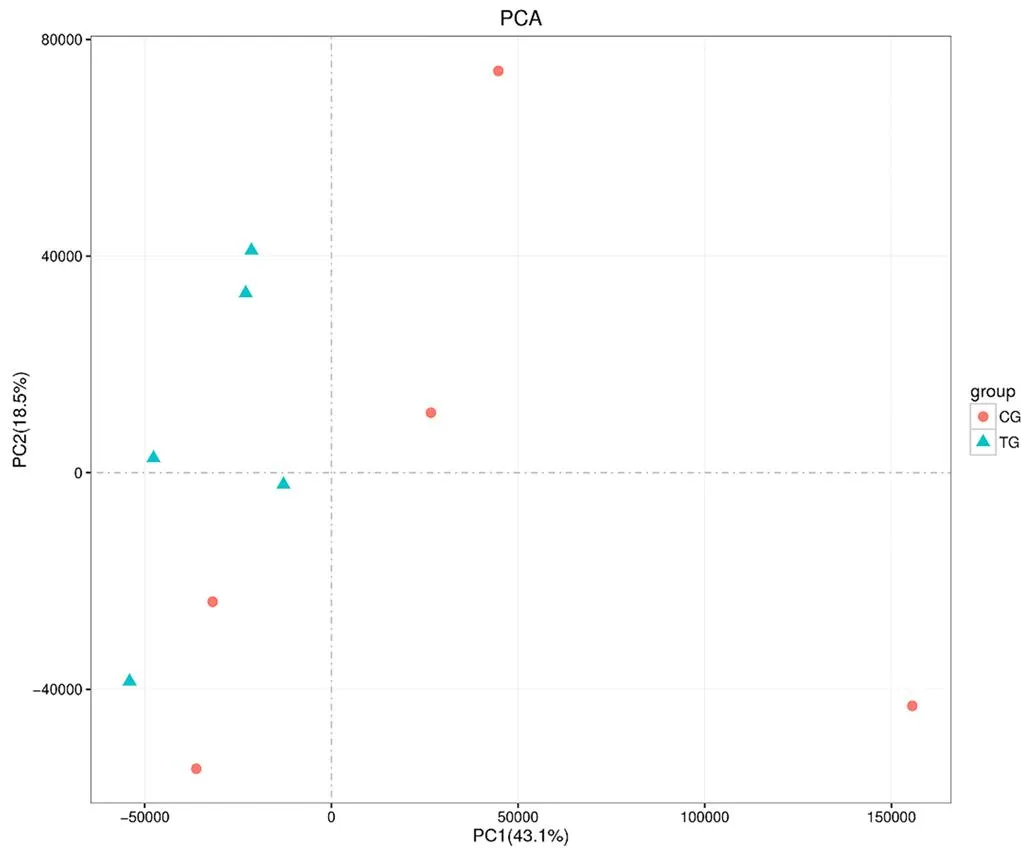
图1 放牧与补饲下藏羊母羊乳汁PCA图
Figure 1 PCA diagram of milk of grazing and supplemented Tibetan sheep ewes
2.2 差异代谢物主成分分析(PCA)
利用主成分分析(principal components analysis, PCA)方法能够直观清楚地反映各处理间的差异性及相似性。由图1所示,两组羊藏羊母羊乳汁中第一主成分(PC1)和第二主成分(PC2)解释率分别为43.10 %和18.50 %,两者累计贡献率可达61.60 %,由此可见,两组间存在较为明显的分离趋势。
2.3 差异代谢物偏最小二乘判别分析(PLS-DA)
采用偏最小二乘法判别分析(partial least squares discriminant analysis, PLS-DA)对构建的模型进行验证,两组藏羊母羊的乳汁样本PLS-DA得分图如图2(a)所示,在负离子模式下所建模型对乳汁的累积判别解释能力R2X = 0.767,R2Y = 0.999,Q2= 0.854,这3个指标越接近于1时表示PLS-DA模型越可靠。PLS-DA置换检验结果如图2(b)所示,负离子模式下R2Y=0.09,Q2=0.215。由此可以看出原模型具有意义,不存在过拟合现象。
2.4 差异代谢物筛选与分析
利用OPLS-DA的VIP值和单变量统计分析t检验中值来筛选两组间藏羊母羊乳汁显著性差异代谢物,差异显著的阈值为:VIP≥1且<0.05。在负离子模式下共筛出N-苯乙酰甘氨酸(N-Phenylacetylglycine)、尿苷-5-二磷酸-D-半乳糖(Uridine-5-diphospho--D-galactose)、柠檬酸盐(Citraconate)、肌醇(Myoinositol)和6-磷酸海藻糖(6-Trehalose phosphate)等已注释39种显著性差异代谢物,其中24种代谢物相对含量显著上调,15种代谢物的相对含量显著下调(见表3)。
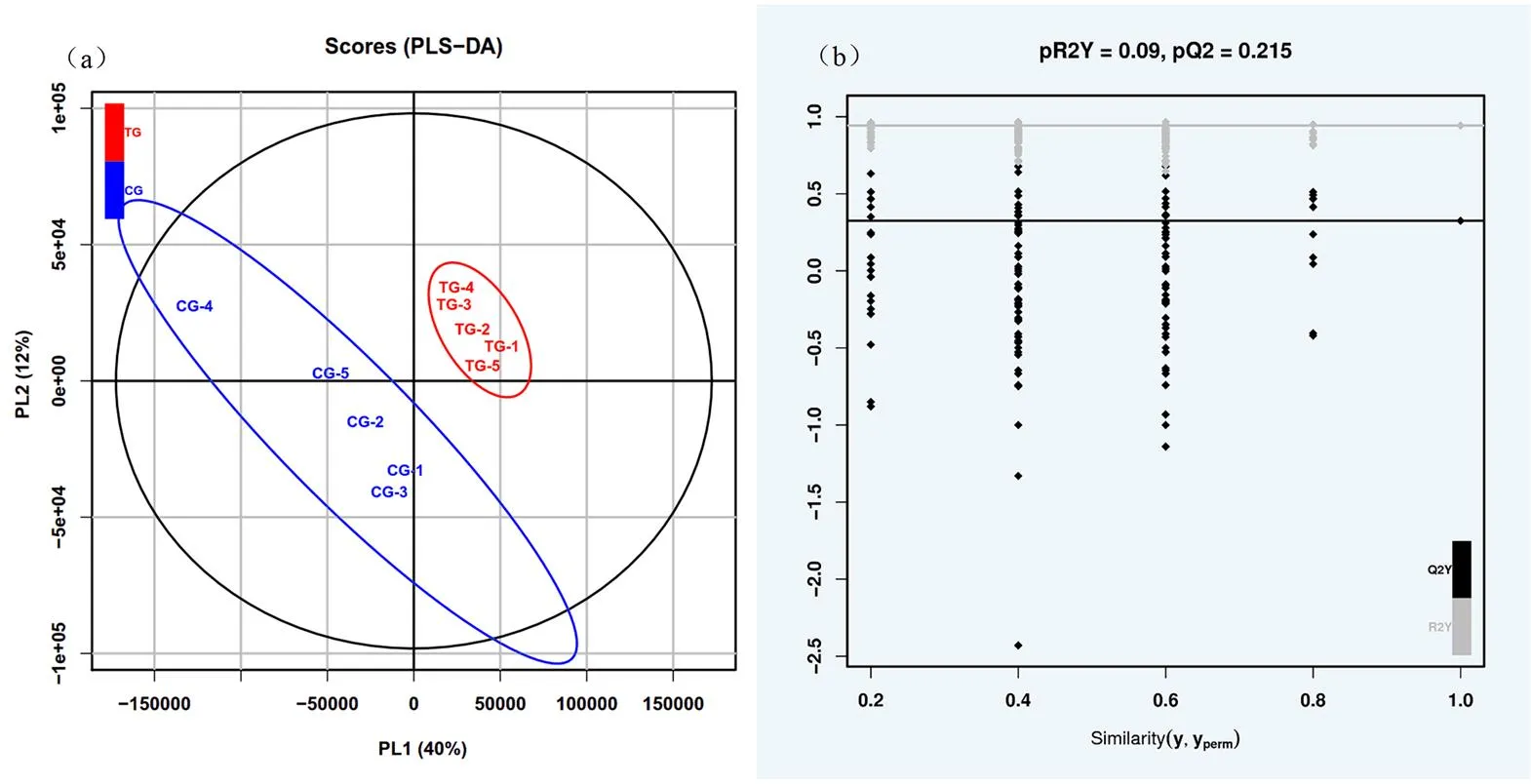
图2 放牧与补饲下藏羊母羊乳汁PLS-DA得分图(a)和PLS-DA置换图(b)
Figure 2 PLS-DA score (a) and PLS-DA replacement (b) of Tibetan sheep ewes ' milk under grazing and supplementary feeding
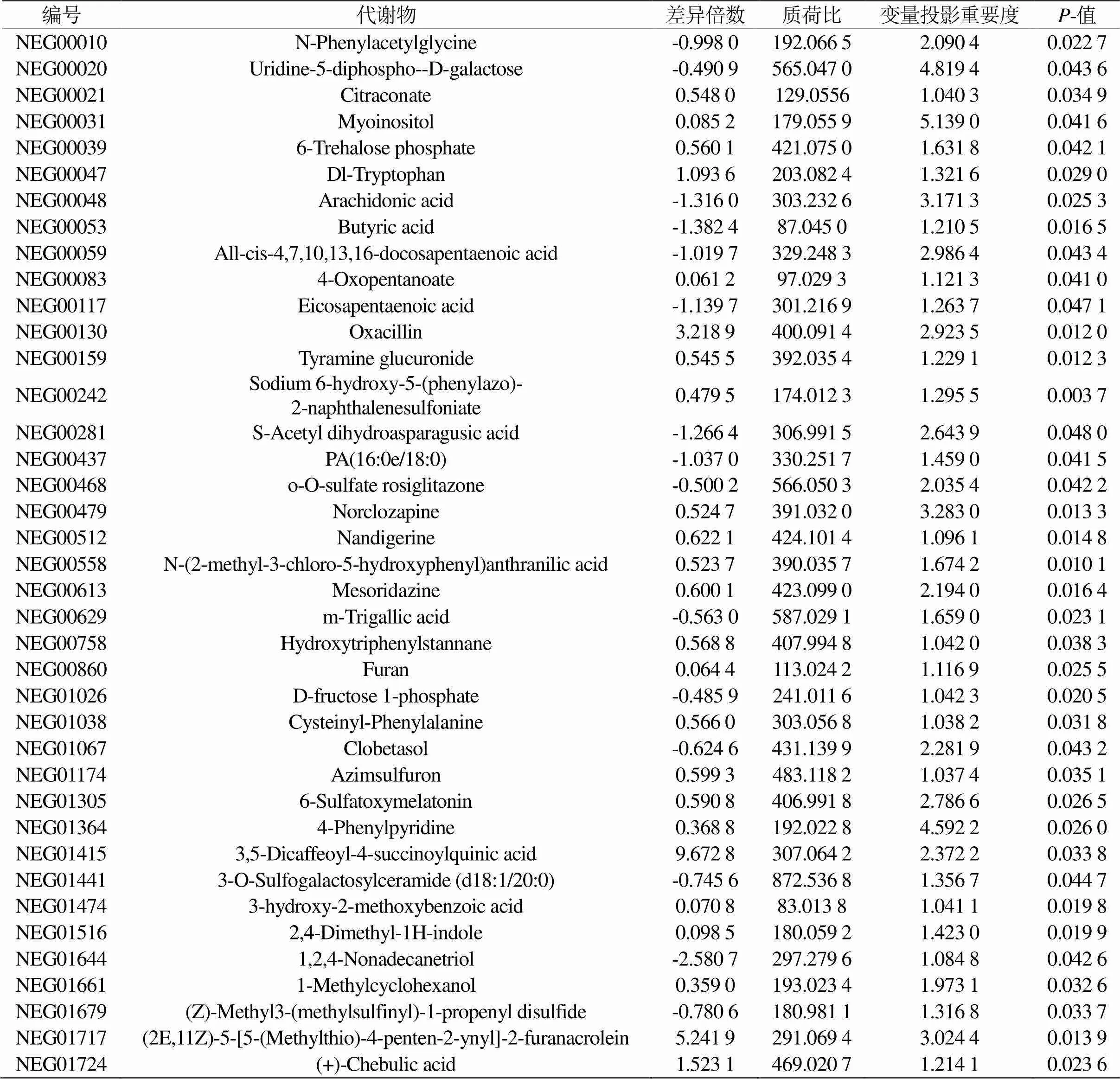
表3 放牧与补饲下藏羊母羊乳汁差异代谢物
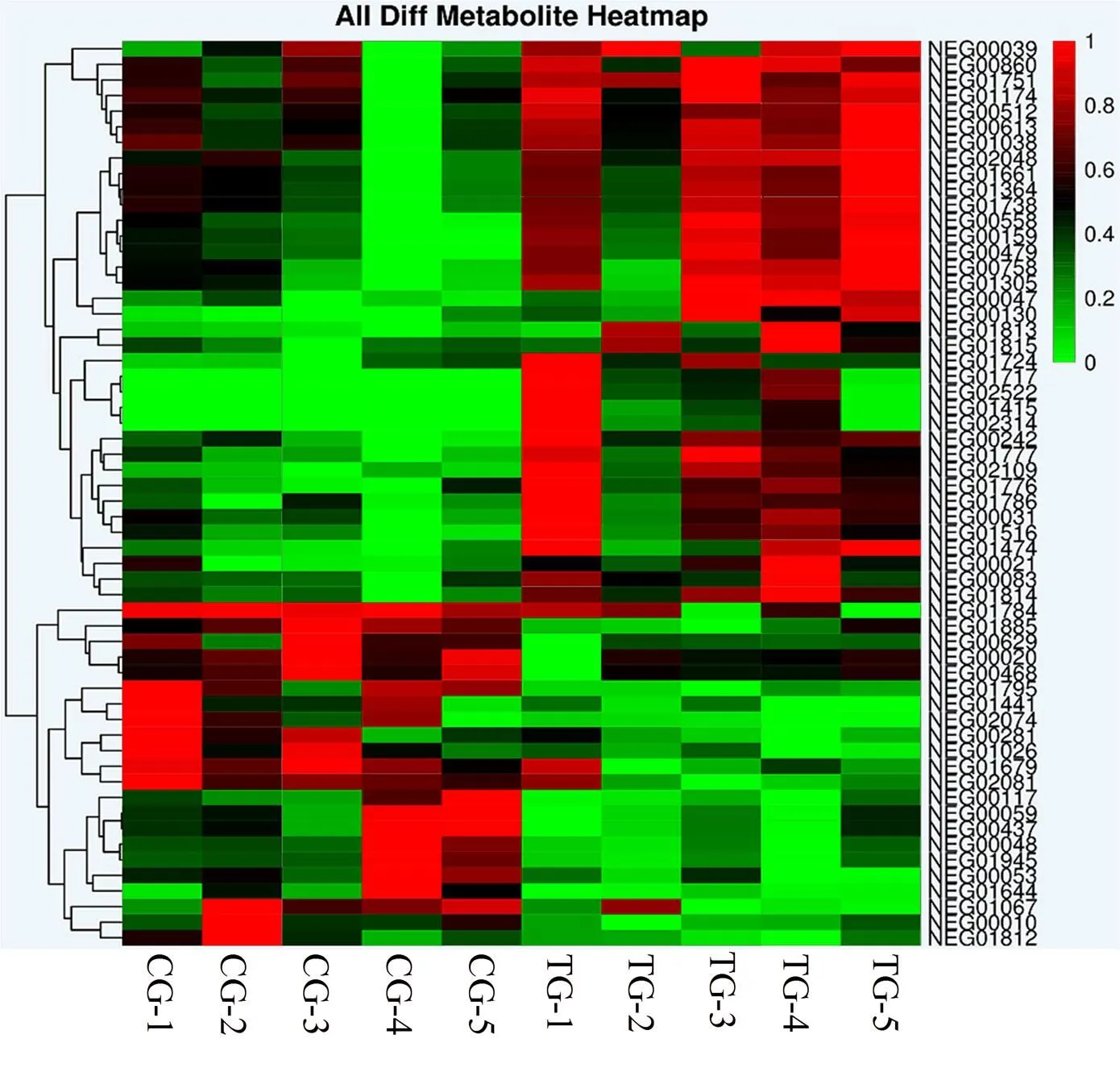
图3 放牧与补饲下藏羊母羊乳汁聚类热图
Figure 3 Heat map of lactation clustering of Tibetan sheep ewes under grazing and supplemental feeding
2.5 差异代谢物聚类分析
负离子模式下两组藏羊母羊乳汁差异代谢物的聚类分析结果如图3所示。图中的颜色代表了乳汁代谢物在该组样本中相对表达量的大小,颜色由蓝到红代表了代谢物丰度逐渐升高。通过层次聚类分析可以看出,与传统放牧相比,放牧+补饲精料对藏羊母羊乳中差异代谢物具有较为明显的影响。
2.6 差异代谢物KEGG富集分析
图4是负离子模式下两组差异代谢物富集于前20条代谢通路如表示,再根据<0.05值,筛选出最为显著的关键代谢通路6条:脂肪细胞中脂肪分解的调节(regulation of lipolysis in adipocyte),含血清素的神经突触(serotonergic synapse),醛固酮的合成和分泌(aldosterone synthesis and secretion),FcεRI信号通路(Fc epsilon RI signaling pathway),不饱和脂肪酸的生物合成(biosynthesis of unsaturated fatty acids),苯丙氨酸、酪氨酸和色氨酸的生物合成(phenylalanine, tyrosine and tryptophan biosynthesis),参与其中代谢物分别为花生四烯酸,二十碳五烯酸和D-果糖-1-磷酸。
3 讨论与结论
羊乳因其优异的营养价值而受到广大消费者的喜爱,其营养成分主要包括糖类、脂肪、蛋白质以及矿物质等[17]。羊乳能够提供各种营养物质,泌乳期母羊产奶量和乳成分对其胎儿生长十分重要[18]。反刍动物的泌乳性能主要受动物品种、饲粮组成、泌乳阶段以及环境条件等多种因素的影响,其中饲养方式也是一个重要的方面。由于藏羊母羊泌乳期正处于寒冷季节,此时牧草供应不足、营养价值低、适口性差,无法为放牧家畜提供充足的养分,因此适宜补饲是维持泌乳时期藏羊母羊生长发育必不可少的环节。产奶量是母畜生产性能的重要指标之一,产奶量越高则说明生产性能越好。以泌乳牦牛为试验对象[19],结果表明其产奶量得到显著提高。在西农萨能奶山羊亦发现[20],精粗比4∶6组的日产奶量、乳糖含量均低于精粗比6∶4组,其中乳糖含量差异显著。上述研究结果显示,随着精料比例的增加可以明显提高反刍动物的产奶量。在本试验中,与CG组相比,TG组的泌乳量提高显著。这可能是由于在精料补饲下非结构性碳水化合物含量较高,小肠氨基酸模式得到优化,有效调节体内泌乳反应,继而提高了产奶量。乳脂率是衡量奶品质的主要指标之一,乳脂率越高则表明奶品质越好[21]。王贤东等[22]研究证实,饲喂不同营养水平饲粮对妊娠后期伊犁马初乳中脂肪无显著影响,这与本试验结果不一致,在本试验中,TG组的乳脂肪含量显著高于CG组,其原因可能是动物品种、饲料组成以及年龄差异等不同造成的。
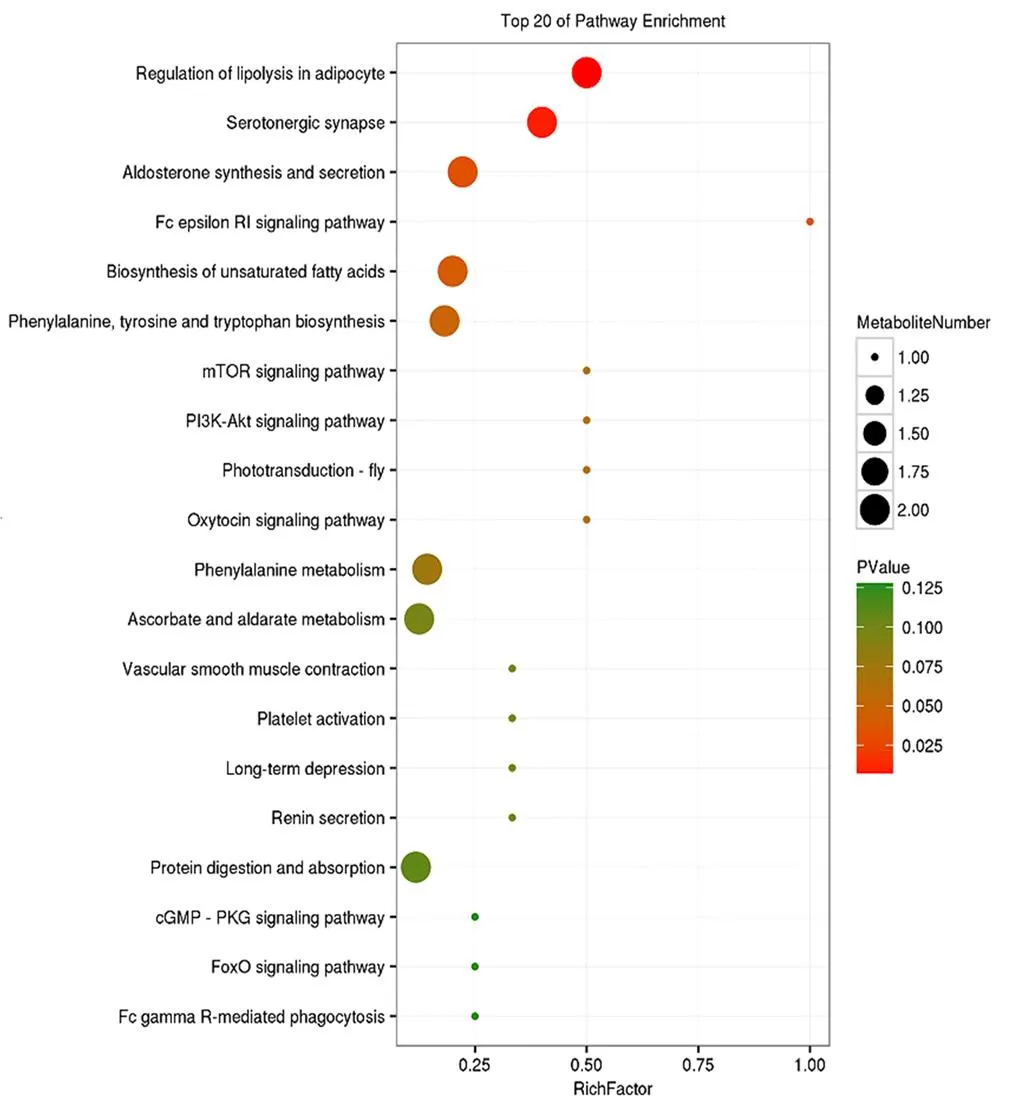
Regulation of lipolysis in adipocyte(脂肪细胞中脂肪分解的调节),Serotonergic synapse(含血清素的神经突触),Aldosterone synthesis and secretion(醛固酮的合成和分泌),Fc epsilon RI signaling pathway(FcεRI信号通路),Biosynthesis of unsaturated fatty acids(不饱和脂肪酸的生物合成),Phenylalanine, tyrosine and tryptophan biosynthesis(苯丙氨酸,酪氨酸和色氨酸的生物合成),mTOR signaling pathway(mTOR信号通路),PI3K-Akt signaling pathway(PI3K-Akt信号通路),Phototransduction-fly(Phototransduction-fly),Oxytocin signaling pathway(催产素信号通路),Phenylalanine metabolism(苯丙氨酸代谢),Ascorbate and aldarate metabolism(抗坏血酸和醛酸代谢),Vascular smooth muscle contraction(血管平滑肌收缩),Platelet activation(血小板活化),Long-term depression(长期抑郁症),Renin secretion(肾素分泌),Protein digestion and absorption(蛋白质消化吸收),cGMP - PKG signaling pathway(cGMP-PKG信号通路),FoxO signaling pathway(FoxO信号通路),Fc gamma R-mediated phagocytosis(FcγR介导的吞噬作用)。
Figure 4 Bubble diagram of differential metabolic pathways of lactation in Tibetan sheep ewes under grazing and supplemental feeding
代谢组学主要通过机体内代谢物的变化来揭示相关生命活动的机制[23]。已经被证实是一种有效的分析乳成分差异的方法[24]。乳汁代谢物组成不仅是机体代谢的重要表现,同时也是评估反刍动物健康、生理状态以及乳质的重要标志物[25]。本试验通过液相色谱-串联质谱技术分别对放牧和补饲下藏羊母羊乳汁进行了测序,鉴定分析出已注释39种显著性差异代谢物,经过KEGG通路分析,最终筛选出最为显著6条关键代谢通路,并映射出花生四烯酸、二十碳五烯酸和D-果糖-1-磷酸等3个为关键代谢产物。花生四烯酸[26]是一种十分常见的多不饱和脂肪酸,在动物界广泛存在。花生四烯酸及其代谢产物在调节动物新陈代谢、维持内环境平衡以及调控生理功能等方面都具有重要功能[27]。研究表明,二十碳五烯酸也是十分重要的多不饱和脂肪酸之一,可减少或预防血脂异常,具有调节血压与葡萄糖代谢、减肥、抵抗炎症以及降低抑郁症等方面功能[28]。此外,多不饱和脂肪酸不仅能够预防、治疗人类许多疾病,同时还可对其脂类代谢具有重要的调节功能[29]。Vanleeuwen等[30]研究表明,多不饱和脂肪酸可抑制动物肌肉中脂肪酸的合成。从本试验来看,与CG相比,TG组的乳中脂肪含量显著提高,推测可能正是由于对照组藏母羊乳中花生四烯酸与二十碳五烯酸含量过高抑制脂肪合成所致。因此,D-果糖-1-磷酸可能参与苯丙氨酸,酪氨酸和色氨酸的生物合成途径来调节氨基酸的合成。
综上,在本试验条件下,冷季补饲提高了泌乳期藏羊母羊乳汁营养水平,并对其代谢产物具有积极的调控作用,这为今后绵羊乳品质的营养调控提供了重要的理论依据。
[1] 毛学英, 韩立华, 袁绮晨, 等. 乳源活性成分功能特性的研究进展[J]. 食品科学技术学报, 2022, 40(2): 15-24.
[2] 房艳,于思雨,高俊海,等. 基于UPLC-QTOF-MS识别牛乳与羊乳差异[J]. 食品工业,2020,41(7):157-162.
[3] MAATHUIS A, HAVENAAR R, HE T, et al. Protein digestion and quality of goat and cow milk infant formula and human milk under simulated infant conditions[J]. J Pediatr Gastroenterol Nutr, 2017, 65(6): 661-666.
[4] SUNARIC S, ZIVKOVIC J, PAVLOVIC R, et al. Estimation of nutritive value of cow and goat milk based on Vitamin E content[J]. Eur J Pharmacol, 2011, 668 (Suppl 1): e23-e24.
[5] MORENO-FERNANDEZ J, DIAZ-CASTRO J, ALFÉREZ M J, et al. Fermented goat milk improves antioxidant status and protects from oxidative damage to biomolecules during anemia recovery[J]. J Sci Food Agric, 2017, 97(5): 1433-1442.
[6] 孙玲伟, 何孟纤, 戴建军, 等. 宫内生长受限湖羊新生羔羊的血浆代谢组学研究[J]. 中国农业科技导报, 2022, 24(7): 123-131.
[7] 周力, 任昊, 马博妍, 等. 日粮精粗比对青海牧区藏羊母羔肺脏抗氧化功能及形态发育的影响[J]. 饲料研究, 2021, 44(18): 7-10.
[8] 王彩莲, 吴建平, 刘立山, 等. 精料补饲水平对藏羊屠宰性能和器官发育的影响[J]. 草业科学, 2021, 38(2): 348-357.
[9] YANG C, HOU F, SUN Y, et al. Oats hay supplementation to yak grazing alpine meadow improves carbon return to the soil of grassland ecosystem on the Qinghai-Tibet Plateau, China[J]. Glob Ecol Conserv, 2020, 23: e01158.
[10] 王威, 张建勋, 康坤, 等. 冷季补饲精料对牦牛繁殖性能和生长性能的影响[J]. 中国畜牧杂志, 2013, 49(7): 78-80.
[11] LI S B, TIAN Y F, JIANG P, et al. Recent advances in the application of metabolomics for food safety control and food quality analyses[J]. Crit Rev Food Sci Nutr, 2021, 61(9): 1448-1469.
[12] 陈雪, 罗欣, 梁荣蓉, 等. 代谢组学在肉及肉制品品质监测中的应用[J]. 农业工程学报,2020,36(17):291-300.
[13] HUANG Q Y, ZHANG J, MARTIN F L, et al. Perfluorooctanoic acid induces apoptosis through the p53-dependent mitochondrial pathway in human hepatic cells: a proteomic study[J]. Toxicol Lett, 2013, 223(2): 211-220.
[14] 吴艳, 李刚, 皮劲松, 等. 鹌鹑蛋蛋清与蛋黄类靶向代谢组学分析[J]. 中国家禽, 2022, 44(9): 1-7.
[15] 钟晓琴,陈佳. 液相色谱质谱联用技术在果蔬农药残留检测中的应用[J]. 食品安全导刊,2020,(30):115-118.
[16] 中华人民共和国农业部. 肉羊饲养标准:NY/T 816—2004[S]. 北京:中国农业出版社,2004.
[17] 张荣, 吴欣雨, 贾玮. 不同泌乳期羊乳和牛乳的高通量定量乳清蛋白质组学[J]. 食品科学, 2022, 43(10): 107-113.
[18] 王思宇, 赵小琪, 李银江, 等. 日粮钙水平对云南半细毛羊乳成分的影响[J]. 中国饲料,2021(13):61-65.
[19] 周义秀, 郝力壮, 刘书杰. 三江源区高寒草场泌乳牦牛冷季补饲精料对其产奶量及乳中矿物质元素含量的影响[J]. 动物营养学报, 2020, 32(9):4194-4204.
[20] 张桂芳,石蕊,姜发彬,等. 不同精粗比日粮对泌乳奶山羊肝脏氨基酸代谢和产奶性能的影响[J]. 南京农业大学学报,2013,36(6): 73-79.
[21] 曹宏斌, 韩旭彪, 王锡波,等. 钙锌混合剂对泌乳期奶牛生产性能及乳品质的影响[J]. 饲料研究, 2022, 45(6): 6-9.
[22] 王贤东, 于全平, 方美烟, 等. 饲粮不同营养水平对伊犁马妊娠后期体重、消化代谢及分娩后初乳成分的影响[J]. 畜牧兽医学报,2017, 48(2):272-279.
[23] 白凤媛, 赵冬梅, 蔡仁军, 等. 基于UHPLC-QE-MS的CCI模型大鼠的血清代谢组学研究[J]. 光谱学与光谱分析,2022,42(8):2528-2531.
[24] 李萌, 卢玺丞, 黄云艳, 等. 基于代谢组学技术分析不同纬度山羊乳的差异性[J]. 分析化学, 2021, 49(11): 1864-1875.
[25] 张林鑫, 黄波, 陈浩林, 等. 贵州黑山羊不同泌乳阶段乳成分及其相关性分析[J]. 黑龙江畜牧兽医, 2021(8): 42-44.
[26] ARAÚJO B C, RODRIGUEZ M, HONJI R M, et al. Arachidonic acid modulated lipid metabolism and improved productive performance of striped bass () juvenile under sub- to optimal temperatures[J]. Aquaculture, 2021, 530: 735939.
[27] 钟宇, 陈滨, 李健, 等. LC-MS/MS分析血浆中脂肪酸及代谢产物[J]. 质谱学报, 2018, 39(3): 310-315.
[28] 王磊, 吴小莉, 崔伟民, 等. EPA微藻油及EPA的功能研究进展[J]. 中国油脂, 2021, 46(8): 101-104.
[29] NAKAMURA M T, YUDELL B E, LOOR J J. Regulation of energy metabolism by long-chain fatty acids[J]. Prog Lipid Res, 2014, 53: 124-144.
[30] VAN LEEUWEN K A, CAMIN F, JERÓNIMO E, et al. Dietary effects on stable carbon isotope composition of fatty acids in polar and neutral fractions of intramuscular fat of lambs[J]. J Agric Food Chem, 2017, 65(43): 9404-9411.
Effects of supplementary feeding in cold season on milk composition and metabolites of Tibetan sheep ewes during lactation
ZHOU Li1,3, HOU Shengzhen1, GUI Linsheng1, LUO Zenghai2
(1. College of Agriculture and Animal Husbandry, Qinghai University, Xining 810016; 2. Qinghai General Station of Animal Husbandry, Xining 810001; 3. School of Animal Science and Technology, Ningxia University, Yinchuan 750021)
Rapid milk analyzer and Liquid chromatography-mass spectrometry (LC-MS) was used to analyze the difference of milk components and metabolites between grazing and supplementary feeding conditions in Tibetan sheep ewes. 440 healthy Tibetan sheep ewes of 3 years old, similar parity and weight (43.85 kg±7.06 kg) were randomly divided into two groups, including natural grazing group (control group, CG) and grazing+supplementary feeding 0.25 kg·d-1concentrate group (test group, TG). The experimental period was 60 days (150-210 days after breeding). At the end of the experiment, the milk of 5 ewes in each group was collected to analyze their nutritional components and detect their metabolites. The results showed that:(1) Compared with control group, the milk yield, fat, protein, ash and non fat solids of Tibetan sheep ewes in test group were increased, and the milk yield and fat content was significantly different (<0.05). (2) There were 39 significantly different metabolites annotated in the milk of Tibetan sheep ewes in the two groups, of which the relative contents of 24 metabolites were significantly increased and the relative contents of 15 metabolites were significantly decreased. Combined with KEGG functional annotation analysis, it was found that regulation of lipolysis in adipocyte, serotonergic synapse, aldosterone synthesis and secretion, FCεFc epsilon RI signaling pathway, biosynthesis of unsaturated fatty acids, biosynthesis of phenylalanine, tyrosine and tryptophan were the most significant key metabolic pathways. Finally, arachidonic acid was screened Eicosapentaenoic acid and D-fructose-1-phosphate was significantly related to the differential metabolites. In conclusion, the quality and metabolites of milk of lactation Tibetan sheep ewes were improved by supplementary feeding during cold season, which provides a theoretical basis and technical support for the research and development of milk of sheep in the future.
Tibetan sheep; concentrate supplementary feeding; metabolome; milk quality; lactation period; fatty acid
10.13610/j.cnki.1672-352x.20230625.021
2023-06-26 16:19:58
S826.83
A
1672-352X (2023)03-0450-07
2022-06-02
青海省科技厅项目(2022NK169)和青海省昆仑英才高端创新创业人才领军人才项目(2020年)共同资助。
周 力,博士研究生。E-mail:472737885@qq.com
通信作者:桂林生,博士,副教授。E-mail:2369830773@qq.com 罗增海,教授。E-mail:597257700@qq.com
[URL] https://kns.cnki.net/kcms2/detail/34.1162.s.20230625.1521.042.html
猜你喜欢
水产科学(2018年2期)2018-03-27
广东饲料(2016年6期)2016-12-01
饲料博览(2016年10期)2016-04-05
中国粮油学报(2016年5期)2016-01-23
中国粮油学报(2016年5期)2016-01-23
畜牧兽医学报(2015年3期)2015-07-05
食品工业科技(2014年15期)2014-03-11
现代检验医学杂志(2014年1期)2014-02-06
食品科学(2013年17期)2013-03-11
食品科学(2013年15期)2013-03-11
