
日粮精粗比对哺乳期犊牛生长性能、瘤胃发酵参数及细菌菌群结构的影响
2023-07-13 03:25马丽娜高海慧康晓冬梁小军
饲料研究 2023年8期
马丽娜 高海慧 康晓冬 梁小军
(宁夏农林科学院动物科学研究所,宁夏 银川 750002)
适宜的日粮精粗比有助于犊牛早期瘤胃微生物生长,促进瘤胃发育,进而提高犊牛的生长性能和养殖效益。吴瑜婷等[1]研究表明,不同精粗比饲粮对鲁西肉用奶牛的血糖、甘油三酯、蛋白含量无明显影响,但饲喂代乳粉牦牛犊牛血清生化指标均优于母乳组[2]。任春燕等[3]研究发现,犊牛开食料中15%中洗涤性纤维(NDF)水平可以增加犊牛断奶后采食量和体重,血清谷丙转氨酶(ALT)、谷草转氨酶(AST)、碱性磷酸酶(ALP)活性呈上升的趋势,提高了血清的抗氧化水平。周蓉等[4]发现,日粮精粗比为63∶35时,断奶犊牛血液白细胞含量显著高于其他组,犊牛的免疫力显著提高。
本试验旨在探讨不同精粗比日粮对哺乳期安格斯犊牛早期体重增长、瘤胃发酵及细菌菌群结构的影响,为早期构建犊牛瘤胃微生物区系提供参考。
1 材料与方法
1.1 试验设计
选择30 头体重(33.0±1.5)kg、出生1 周左右的健康安格斯犊牛(公、母各半),随机分为3 组,每组10 个重复,每个重复1 头牛。试验1 组犊牛饲喂高精粗比日粮(精粗比60∶40),试验2 组(对照组)犊牛饲喂中等精粗比日粮(精粗比50∶50),试验3 组犊牛饲喂低精粗比日粮(精粗比40∶60)。预试期10 d,正式试验期90 d。
1.2 饲养管理
犊牛随母哺乳,7 d开始补饲颗粒料,每日饲喂2次。试验犊牛每组每日苜蓿草饲喂量为2 kg,给予充足饮水。试验期间,犊牛补饲日粮组成及营养水平见表1。

表1 补饲日粮组成及营养水平(干物质基础)
1.3 测定指标及方法
1.3.1 生长性能
犊牛出生时,测定初生重,在试验开始(犊牛7 日龄)和结束(犊牛90 日龄)晨饲前称重。每周连续3 d称量犊牛饲草料剩余量,记录采食量,计算平均日增重和料重比。
1.3.2 瘤胃液样品采集与处理
试验结束时,利用犊牛瘤胃液采集器插入犊牛瘤胃抽取瘤胃液,分装至50 mL离心管中,置于-80 ℃冻存。
1.3.3 瘤胃液发酵参数
采用冯宗慈等[5]改进的比色法测定氨态氮含量,采用气相色谱-质谱联用(GC-MS)的方法测定挥发性脂肪酸含量。
1.3.4 瘤胃细菌菌群结构
将瘤胃液送至西安擎科生物科技有限公司,采用16S rDNA高通量测序技术对犊牛瘤胃细菌菌群种类进行检测。
1.4 数据统计与分析
试验数据通过Excel初步整理,利用SAS 8.02软件进行单因素方差分析,LSD 法进行多重比较。结果以“平均值±标准差”表示,P<0.05 表示差异显著,P<0.01 表示差异极显著。
2 结果与分析
2.1 日粮精粗比对犊牛生长性能的影响(见表2)

表2 日粮精粗比对犊牛生长性能的影响
由表2可知,试验1组犊牛90日龄体重、采食量、平均日采食量、平均日增重极显著高于试验3组(P<0.01),料重比显著高于试验2组、试验3组(P<0.05)。
2.2 精粗比日粮对犊牛瘤胃发酵参数的影响(见表3)
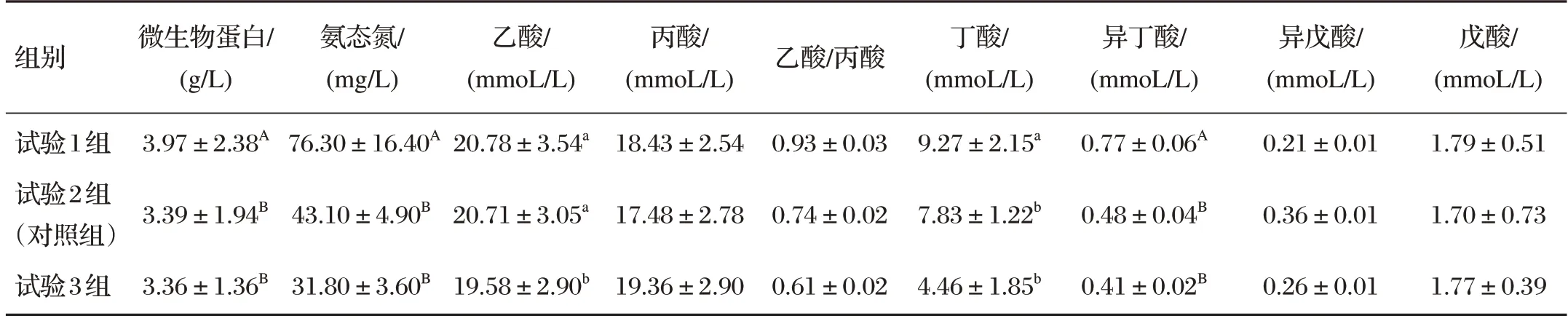
表3 日粮精粗比对犊牛瘤胃发酵参数的影响
由表3可知,试验1组微生物蛋白、氨态氮和异丁酸含量均极显著高于试验2 组、试验3 组(P<0.01),试验1组乙酸、丁酸含量均显著高于试验3组(P<0.05)。
2.3 日粮精粗比对瘤胃菌群Alpha多样性的影响
2.3.1 犊牛瘤胃细菌菌群OTU分析(见图1)
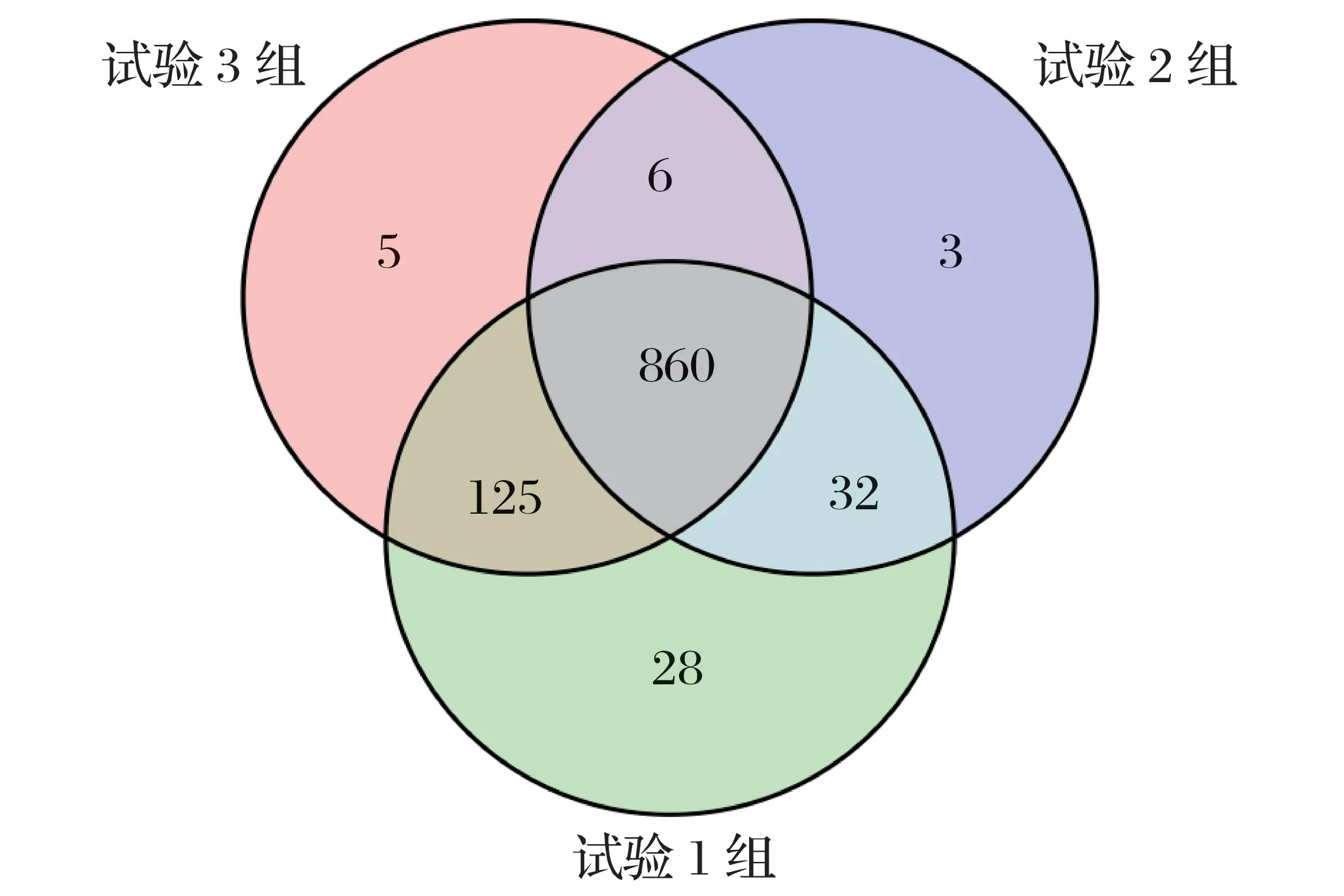
图1 犊牛瘤胃细菌菌群OTU韦恩图
选用相似水平为97%的OTU样本表制成Venn图。
由图1 可知,试验1 组犊牛瘤胃细菌OTU 数为1 045,特有的OTU 数为28,明显高于试验2 组、试验3 组。
2.3.2 不同日粮精粗比对瘤胃菌群多样性指数的影响(见表4)
由表4可知,本试验菌群检测的物种覆盖率为100%。试验1 组、试验3 组Ace、Chaol、Shannon 指数均极显著高于试验2组(P<0.01)。
2.4 不同日粮精粗比对犊牛瘤胃细菌菌群结构的影响
2.4.1 日粮精粗比对犊牛门水平上瘤胃微生物的影响(见表5)

表5 日粮精粗比对犊牛门水平上瘤胃微生物的影响 单位:%
由表5 可知,试验3 组犊牛瘤胃拟杆菌门、后壁菌门、黏胶球形菌门、螺旋菌门、纤维杆菌门的相对丰度极显著高于试验1 组(P<0.01)。各组瘤胃蓝藻门、放线菌门、软壁菌门、嗜热球菌门差异不显著(P>0.05)。
2.4.2 日粮精粗比对犊牛属水平上瘤胃微生物的影响(见表6)

表6 日粮精粗比对犊牛属水平上瘤胃微生物的影响 单位:%
由表6可知,试验1组犊牛瘤胃普雷沃氏菌属、密螺旋体属、螺旋菌属、丁酸弧菌_2 属的相对丰度极显著高于试验2 组、试验3 组(P<0.01)。各组犊牛瘤胃的瘤胃球菌科-NK4A214群差异不显著(P>0.05)。
3 讨论
3.1 不同精粗比对犊牛生长性能的影响
犊牛出生后7 d可进行早期补饲,哺乳期适时饲喂固体颗粒饲料能满足犊牛生长发育所需的营养成分,可通过对瘤胃组织的生理刺激,逐步实现饲料从液态至固态的过渡[6]。日粮结构会影响反刍动物瘤胃微生物多样性和组成[7]。日粮精粗比会明显影响动物体重。研究发现,育肥后期日粮精粗比为65∶35时,牦牛总增重和平均日增重下降[8]。饲粮中的精粗比例与反刍动物饲料有效利用有很大关系[9]。李斌昌[10]研究发现,在日粮精料水平提高30%~50%,9~15月龄的后备奶牛消化率显著提高。徐文慧等[11]研究发现,泌乳新疆褐牛全混合日粮精料比为50∶50,可增加产奶量,延长反刍时间。本研究表明,在不同日粮精粗比例下,犊牛体重、采食量均会发生变化,但试验1 组犊牛日增重显著高于试验2 组、试验3 组。综合分析,高精粗比日粮可增加哺乳期犊牛采食量,提高增重。
3.2 日粮精粗比对犊牛瘤胃发酵参数的影响
瘤胃利用微生物降解粗饲料,通过产生挥发性脂肪酸合成微生物蛋白,为机体提供能量和蛋白质,促进动物生长发育[12]。日粮中蛋白质在瘤胃中的主要降解产物为氨态氮(NH3-N),NH3-N 可为瘤胃中的微生物提供18%~100%的氮,保证瘤胃液中微生物蛋白产量正常。饲草料在瘤胃进行发酵会产生脂肪酸,主要有乙酸、丙酸、丁酸、异丁酸、异戊酸和戊酸。王安思[13]研究发现,饲喂不同日粮精粗比的犊牛,其瘤胃组织的形态和颜色会有明显变化。沈芳等[14]研究发现,舍饲组高精料日粮饲喂条件下,牦牛犊牛瘤胃液NH3-N、总挥发性脂肪酸浓度均极显著提高,因此高精料日粮可以有效促进牦牛犊牛的瘤胃发酵。李岩等[8]研究发现,瘤胃液中异丁酸含量随着日粮中精料的增加而升高,异戊酸含量随着日粮精料增加先升高再降低。本研究中,高精料日粮饲喂下试验1 组犊牛瘤胃微生物蛋白、氨态氮和异丁酸浓度极显著高于试验2、试验3组,乙酸、丁酸含量显著高于试验3组,与上述研究结果一致。
3.3 日粮精粗比对犊牛瘤胃细菌菌群多样性的影响
在反刍动物幼龄时期,消化道中的微生物定植会对其生长发育产生影响,对机体具有长期健康防护作用。研究表明,反刍动物的瘤胃真菌在出生后1 周左右进行初期定植,在出生3 周后初步完成定植。日龄的变化会对反刍动物瘤胃中菌属群落产生较为明显的影响,因此不同日龄反刍动物瘤胃中主导的优势菌属也不同[6]。
利用16S rDNA 对瘤胃微生物进行高通量测序时,Shannon指数、Chao1指数和覆盖度是衡量瘤胃菌群多样性的评定指标。Shannon指数显示瘤胃组织中微生物分布和多样性,Ace指数显示物种丰富程度,Chao1指数反映物种的总数[14]。在本研究中,3组瘤胃测序数据的覆盖度为100%,表明样本符合要求(测序样本的覆盖面超过0.97),能够真实反映3个OTU的覆盖率和序列深度,表明数据可科学充分反映犊牛瘤胃菌群的实际情况。在本试验条件下,试验1组、试验3组Ace、Chaol和Shannon指数显著高于试验2组,与沈芳等[14]研究结果一致,印证了瘤胃菌群结构会随日粮的不同而发生变化,不同的饲料精粗比会在一定程度上改变犊牛瘤胃的菌群数量和种类。
反刍动物出生后开始与外界微生物接触,其瘤胃内的微生物受动物日龄、品种以及饮食结构的影响。瘤胃中的微生物主要有厌氧细菌、古生菌、真菌和原虫,其中细菌数量最多。成年反刍动物每克瘤胃液中细菌约有1 010~1 011 个,反刍动物通过瘤胃细菌对饲草料的消化分解消化来获取充足的营养和能量。黄金峰等[15]研究发现,瘤胃微生物组成很大程度上受饲料组成和采食模式影响。冶文兴等[16]研究发现,犊牛开食料中添加5 g甘露寡糖会大幅增加变形菌门、互养菌门、琥珀酸弧菌属、新月形单胞菌属的丰富程度,杆菌属和优杆菌属的含量明显下降。徐晓锋等[7]研究发现,日粮精粗比会对瘤胃微生物的数量和种类具有明显影响。孙光明等[17]研究表明,日粮中精料水平提高,瘤胃中氨态氮和总氮含量会明显增加,但微生物蛋白含量明显下降,因此,低精料日粮能够促进氨态氮的利用并能够有效调节瘤胃氮平衡。远德龙等[18]研究发现,饲喂含有枣粉和梨渣的日粮可以显著提高奶牛瘤胃总挥发性脂肪酸含量。郝文君等[19]研究发现,不同GI 与CBI组合日粮对牦牛瘤胃放线菌门具有显著差异,对牦牛瘤胃细菌区系属水平影响不显著。张俊等[20]通过Meta 分析探究发现,日粮精粗比与后备奶牛的干物质采食量和瘤胃丙酸比例具有显著的一元一次回归关系;在细菌门水平上,以拟杆菌门和纤维杆菌门为主要优势菌;在属水平上,共涉及124 个属,其中优势微生物菌属包含普雷沃氏菌属、密螺旋体属、琥珀酸弧菌科-UCG-001 属、纤维杆菌属、瘤胃球菌属等。普雷沃氏菌属属于普雷沃氏菌科,普雷沃氏菌对促进饲料中淀粉、蛋白质和纤维素在瘤胃中的消化代谢[21],瘤胃球菌会促进动物机体蛋白质代谢[22],其相对丰度在高精料日粮饲喂的湖羊瘤胃具有较高表达[23]。拟杆菌门对饲草料碳水化合物的消化吸收具有积极作用,尤其是对饲草料植物细胞壁有明显的降解作用[24],表明试验1组犊牛体重的显著增加与瘤胃中主要优势菌有关,在瘤胃中主要优势菌促进饲草料消化,增加机体营养成分的吸收,为犊牛生长发育提供能量。
4 结论
在本试验条件下,哺乳期犊牛饲喂日粮精粗比为60∶40 有利于犊牛早期生长发育,改善瘤胃发酵参数,瘤胃细菌在门水平上影响最为明显。
猜你喜欢
中国饲料(2021年17期)2021-11-02
当代水产(2021年2期)2021-03-29
传染病信息(2021年6期)2021-02-12
科学(2020年2期)2020-08-24
中成药(2018年10期)2018-10-26
现代检验医学杂志(2016年1期)2016-11-12
湖南畜牧兽医(2016年3期)2016-06-05
兽医导刊(2016年12期)2016-05-17
烟草科技(2015年8期)2015-12-20
动物营养学报(2015年10期)2015-12-01
