
布鲁菌二联BCSP31-L7/L12-IL-2嵌合病毒样颗粒疫苗候选株的构建与鉴定
2023-07-08 12:34:36邹映雪黄天鸿冯嘉轩丁佳欣陈铭桦陈凯楠郭春红
中国兽医学报 2023年5期
邹映雪,黄天鸿,冯嘉轩,丁佳欣,陈铭桦,陈凯楠,郭春红,张 頔,丁 壮
(吉林大学 动物医学学院 人兽共患病研究教育部重点实验室,吉林 长春130062)
布鲁菌病是由布鲁菌所引起一种慢性、持续性感染的人畜共患传染病[1]。该病自首次被证实以来在世界范围内广泛存在。目前已知可感染布鲁菌的动物达60余种,猪、牛、羊为主要传染源。 该病可引起母畜流产、死胎,人睾丸炎及生殖系统损伤等症状,被世界卫生组织认为是七大被忽视的人畜共患病之一[2]。因此,通过疫苗的接种对于控制并最终消除这种疾病至关重要。目前,布鲁菌病常用的传统疫苗主要为Rev-1、S19及S2等减毒活苗,但上述活苗存在副作用较强、种属局限性较大、接种后干扰血清学诊断及可能出现毒力返强等缺点[3]。由此,针对传统疫苗免疫种属局限性、保护局限性以及单一抗原组分无法引起有效细胞免疫应答,保护效果相对有限的特点,研发一种绿色、安全、高效、“一针多防”、可实现多方位免疫且在各菌种间高度保守的布鲁菌新型疫苗——布鲁菌嵌合病毒样颗粒,对布鲁菌病的防控可发挥重要作用。L7/L12蛋白广泛存在于各个种的布鲁菌中,在各种间高度保守,是布鲁菌优势抗原蛋白之一[4]。BCSP31基因在布鲁菌中均可表达(除了绵羊附睾布鲁菌外),可刺激机体对布鲁菌感染产生保护性免疫应答,上述2种蛋白可以作为开发疫苗的候选抗原[5]。IL-2又名T细胞生长因子(T cell growth factor,TCRF),具有促进T、B淋巴细胞增殖分化,诱导干扰素(interferon,IFN)与肿瘤坏死因子(tumor necrosis factor,TNF)等细胞因子的分泌[6],增强机体免疫力,充当佐剂增强疫苗保护力、降低疫苗副作用等功能,可作为增强新型疫苗保护效力的候选免疫增强因子。
病毒样颗粒(VLPs)是由病毒的一种或几种结构蛋白自行组装而成的颗粒,具有与病毒颗粒类似的空间结构[7]。与传统疫苗相比,VLPs不含病毒核酸,不能进行自主复制,具有极高的安全性,还可通过MHCⅠ和MHCⅡ交叉提呈方式增强抗原提呈细胞的摄取、加工和提呈能力,并通过与病毒粒子相同的途径诱导机体产生较强的天然性免疫与获得性免疫反应[8]。近些年来,VLPs技术的应用逐渐广泛,以VLPs为载体研发疫苗也已成为新的方向。VLPs疫苗作为一种新型疫苗,因其绿色安全的特性及其具有强大的免疫优势,为基因工程疫苗的研究提出了很好的方向,这种绿色生态、安全高效的非全病原疫苗或新型疫苗也已成现在和未来的疫苗研发主流[9]。
因此,本研究基于实验室已成功构建的昆虫杆状病毒表达系统,以实验室保留的基因Ⅶ型NA-1株M的ND VLPs为基础,分别通过GPI锚定策略将IL-2、布鲁菌病保护性抗原蛋白L7/L12、BCSP31引入该VLPs,构建表达布鲁菌L7/L12、BCSP31蛋白的二联嵌合型布鲁菌VLPs,弥补目前使用传统疫苗不足,以期为布鲁菌病的防制提供新策略。
1 材料与方法
1.1 细胞、病毒及载体EscherichiacoliDH10Bac感受态细胞、Sf9昆虫细胞均为本实验室保存;EscherichiacoliDH5α感受态细胞购自北京全式金生物技术有限公司;rBv-M,rBv-BCSP31,pFastBac1-GPI空载体为本实验室保存。
1.2 主要试剂2×M5 HiPer plus Taq HiFi PCR mix购自北京聚合美生物技术有限公司;质粒小提试剂盒、胶回收试剂盒购自Axygen公司;T4连接酶购自全式金公司;胎牛血清购自BI公司;昆虫培养基Sf-900TMⅢ SFM (1×)购自Gibco公司;X-treme GENEHP DNA Transfection Reagent购自ROCHE公司;其他试剂均为国产分析纯。
1.3 引物设计根据GenBank中IL-2(鼠IL-2序列,GenBank:NM-008366.3)、布鲁菌S2型L7/L12(ANH46869)序列分别设计IL-2-F、IL-2-R、L7/L12-F、L7/L12-R鉴定引物以及pFastBac1-GPI通用引物送至上海生物公司合成,序列信息见表1。

表1 引物信息
1.4 目的基因的合成根据GenBank中IL-2(鼠IL-2序列,GenBank:NM-008366.3)、布鲁菌S2型L7/L12(ANH46869)序列,分别在上游添加SalⅠ酶切位点,下游添加KpnⅠ酶切位点,由生工生物工程(上海)股份有限公司合成并克隆至pUC57载体,分别命名为pUC57-L7/L12、pUC57-IL-2。
1.5 分别表达L7/L12、IL-2基因的重组穿梭质粒的构建及鉴定将实验室保存pFastBac1-GPI空载体及公司合成pUC57-L7/L12、pUC57-IL-2分别用SalⅠ、KpnⅠ进行双酶切,通过1%核酸电泳鉴定正确后,分别胶回收产物。使用T4连接酶将pFastBac1-GPI线性载体分别与L7/L12、IL-2连接后转化至EscherichiacoliDH5α感受态细胞。次日挑取单菌落分别使用其特异性引物进行PCR鉴定,取鉴定正确的菌株接种至LB液体培养基过夜培养后提取质粒。利用SalⅠ、KpnⅠ进行双酶切鉴定正确后,送至生工生物工程(上海)股份有限公司测序,将鉴定正确的重组穿梭质粒分别命名为pFastBac1-GPI-L7/L12、pFastBac1-GPI-IL-2。
1.6 分别表达L7/L12、IL-2基因的重组杆粒的构建及鉴定将pFastBac1-GPI-L7/L12、pFastBac1-GPI-IL-2分别转化至EscherichiacoliDH10Bac感受态细胞,次日挑取单菌落分别使用特异性引物及M13引物进行PCR鉴定,特异性引物及M13引物均鉴定为阳性的菌液再次在3抗固体培养基上进行第2次划线筛选。重复上述菌落划线和菌液PCR,将连续3次筛选正确的菌液视为阳性菌液。提取杆粒,将鉴定正确的杆粒命名为rBacmid-GPI-L7/L12、rBacmid-GPI-IL-2。
1.7 分别表达L7/L12、IL-2基因的重组杆状病毒的拯救与鉴定用3.5 cm细胞培养板接种Sf9细胞,待其生长密度为80%~85%时分别将rBacmid-GPI-L7/L12、rBacmid-GPI-IL-2与转染试剂按照1∶3的比例混合均匀并静置15 min后接种细胞,27℃培养96 h,收集上清为P1代。将P1代接入到培养密度为80%~85%且状态良好的Sf9细胞中,27℃培养96 h,再次收集上清视为P2代。盲传4代后,收集P4代杆状病毒,提取基因组,分别用特异性引物及通用引物进行PCR鉴定,将结果正确的样品命名为rBv-L7/L12、rBv-IL-2。
1.8 分别表达L7/L12、IL-2基因的重组杆状病毒滴度的测定使用杆状病毒滴度快速测定试剂盒测定病毒滴度,具体操作见说明书。
1.9 IFA检测将rBv-L7/L12、rBv-IL-2及实验室保留的rBv-M、rBv-BCSP31以MOI=5共感染培养密度为80%~85%且状态良好的Sf9细胞,同时设置未感染组作对照,培养48 h后,用4%多聚甲醛溶液将细胞于4℃固定10 min,再使用Triton-100室温通透10 min,分别以L7/L12、IL-2、NDV M及BCSP31的多克隆抗体为一抗,4℃过夜孵育后,用FITC标记的猪抗兔IgG抗体作为二抗室温孵育1 h,用DAPI于室温下染核10 min,利用倒置荧光显微镜观察各组分蛋白的表达。
1.10 BCSP31-L7/L12-IL-2 cVLPs的制备与鉴定
1.10.1BCSP31-L7/L12-IL-2 cVLPs的制备 将构建成功的P4代rBv-L7/L12、rBv-IL-2与实验室保存的rBv-M、rBv-BCSP31以MOI=5共感染悬浮培养密度为2×106个/mL的Sf9细胞,27℃、200 r/min 转动培养96 h后收集细胞悬液,分别经过20%,30%,60%蔗糖,100 000 ×g离心2 h后获得纯化BCSP31-L7/L12-IL-2 cVLPs。
1.10.2BCSP31-L7/L12-IL-2 cVLPs的鉴定 以L7/L12、IL-2、NDV M及BCSP31的多克隆抗体为一抗,对纯化后获得的BCSP31-L7/L12-IL-2 cVLPs进行Western blot鉴定,利用透射电镜观察其形态特征。
2 结果
2.1 重组穿梭质粒pFastBac1-GPI-L7/L12、pFastBac1-GPI-IL-2的鉴定利用SalⅠ、KpnⅠ对pFastBac1-GPI-L7/L12、pFastBack1-GPI-IL-2进行双酶切鉴定,将产物经1%琼脂糖凝胶电泳鉴定,可见pFastBac1-GPI-L7/L12酶切后产生大小为375,4 936 bp的2条条带,pFastBack1-GPI-IL-2酶切后产生大小为510,4 963 bp的2条条带,与预期大小相符(图1),测序结果显示无突变。
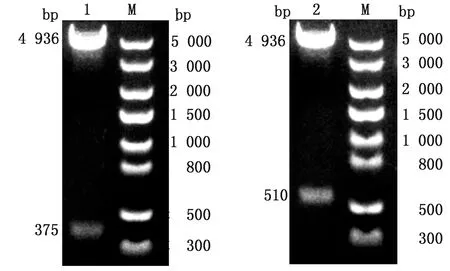
M.DL5000 DNA Marker;1.pFastBac1-GPI-L7/L12双酶切;2.pFastBac1-GPI-IL-2双酶切
2.2 重组杆粒rBacmid-GPI-L7/L12、rBacmid-GPI-IL-2的鉴定3次蓝白斑筛选后,将鉴定正确的菌液接菌扩大培养后提取杆粒,分别使用特异性引物及M13通用引物进行PCR,扩增片段经1%琼脂糖凝胶电泳进行鉴定。结果显示,L7/L12特异性引物扩增产物大小为375 bp,IL-2特异性引物扩增产物大小为375 bp,M13通用引物扩增rBacmid-GPI-L7/L12获得片段大小为2 875 bp,M13通用引物扩增rBacmid-GPI-IL-2获得片段大小为3 010 bp(图2)。

A.特异性引物PCR扩增rBacmid-GPI-L7/L12结果(M1.DL2000 DNA Marker;1.PCR产物 );B.通用引物PCR扩增rBacmid-GPI-L7/L12产物(M2.DL5000 DNA Marker;2.PCR产物);C.特异性引物PCR扩增rBacmid-GPI-IL-2产物(M1.DL2000 DNA Marker;3.PCR产物);D.通用引物PCR扩增rBacmid-GPI-IL-2产物(M2.DL5000 DNA Marker;4.PCR产物)
2.3 重组杆状病毒rBv-L7/L12、rBv-IL-2的鉴定
2.3.1Sf9细胞病变观察 将杆粒rBacmid-GPI-L7/L12、rBacmid-GPI-IL-2转染至Sf9细胞,用光学显微镜观察发现,与正常Sf9细胞相比,转染组细胞生长缓慢,细胞变大、变圆、呈颗粒变性状,且有细胞产生脱落、破碎等典型病变(图3)。
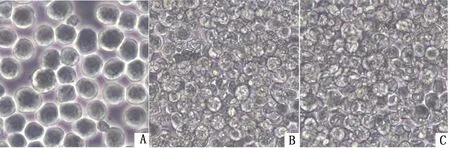
1.正常Sf9细胞;B.rBv-L7/L12感染后细胞;C.rBv-IL-2感染后细胞
2.3.2重组杆状病毒rBv-L7/L12、rBv-IL-2的鉴定 将杆粒rBacmid-GPI-L7/L12、rBacmid-GPI-IL-2转染至Sf9细胞,收集上清盲传4代,提取P4代杆状病毒的基因组,用特异性引物以及M13通用引物进行PCR鉴定,结果显示,扩增条带大小与预期相符(图4)。

A.特异性引物PCR扩增rBacmid-GPI-L7/L12结果(M1.DL2000 DNA Marker;1.PCR产物 );B.通用引物PCR扩增rBacmid-GPI-L7/L12产物(M2.DL5000 DNA Marker;2.PCR产物);C.特异性引物PCR扩增rBacmid-GPI-IL-2产物(M1.DL2000 DNA Marker;3.PCR产物);D.通用引物PCR扩增rBacmid-GPI-IL-2产物(M2.DL5000 DNA Marker;4.PCR产物)
2.3.3IFA检测 利用激光共聚焦显微镜观察cVLPs的各组分蛋白表达情况,可见各试验组与未感染组相比均出现绿色荧光信号,表明各组分蛋白均正确表达(图5)。
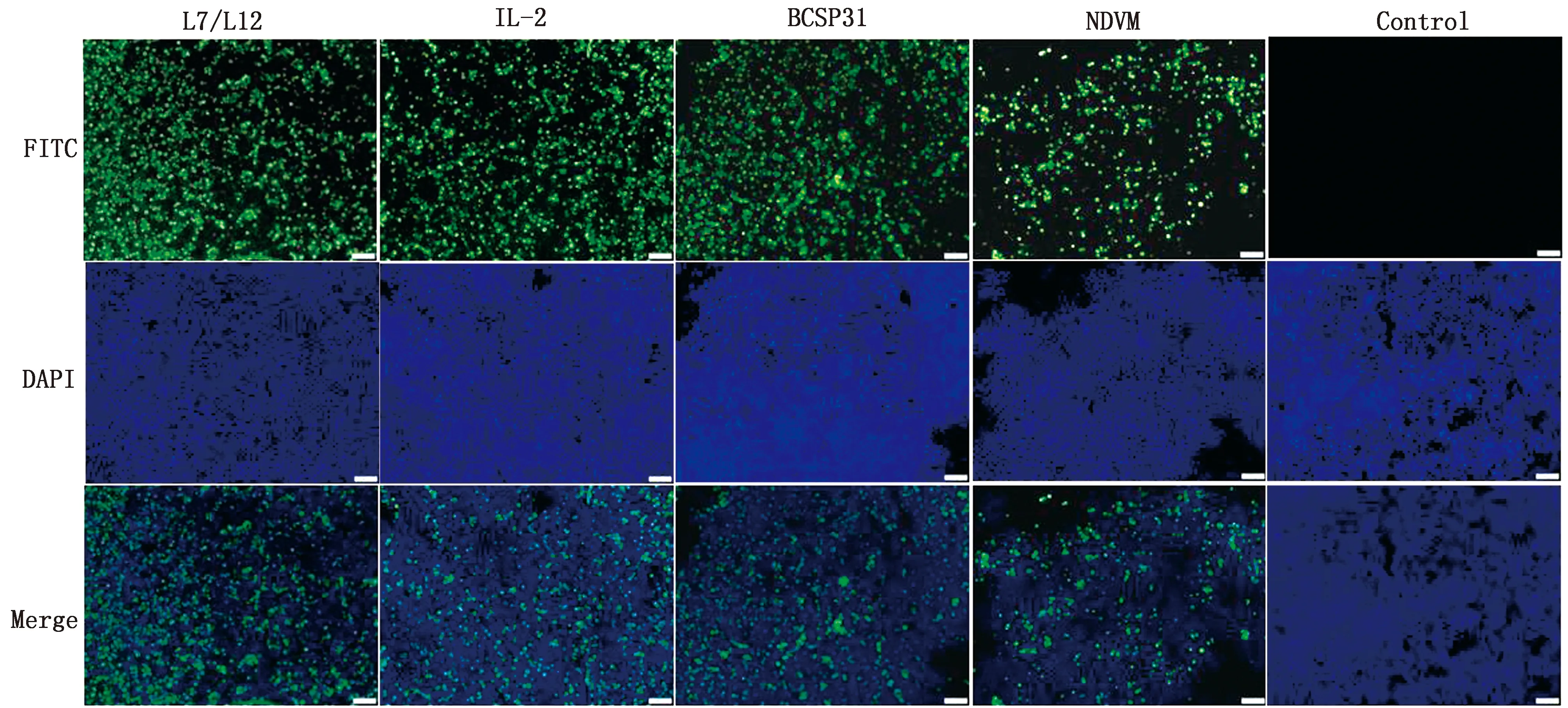
图5 IFA检测各蛋白表达(×200)
2.4 BCSP31-L7/L12-IL-2 cVLPs的鉴定
2.4.1BCSP31-L7/L12-IL-2 cVLPs的透射电镜观察 透射电镜观察纯化后的cVLPs形态,粒子呈球形,大小约为100 nm,表面存在纤突结构,与野生型NDV粒子相似(图6)。
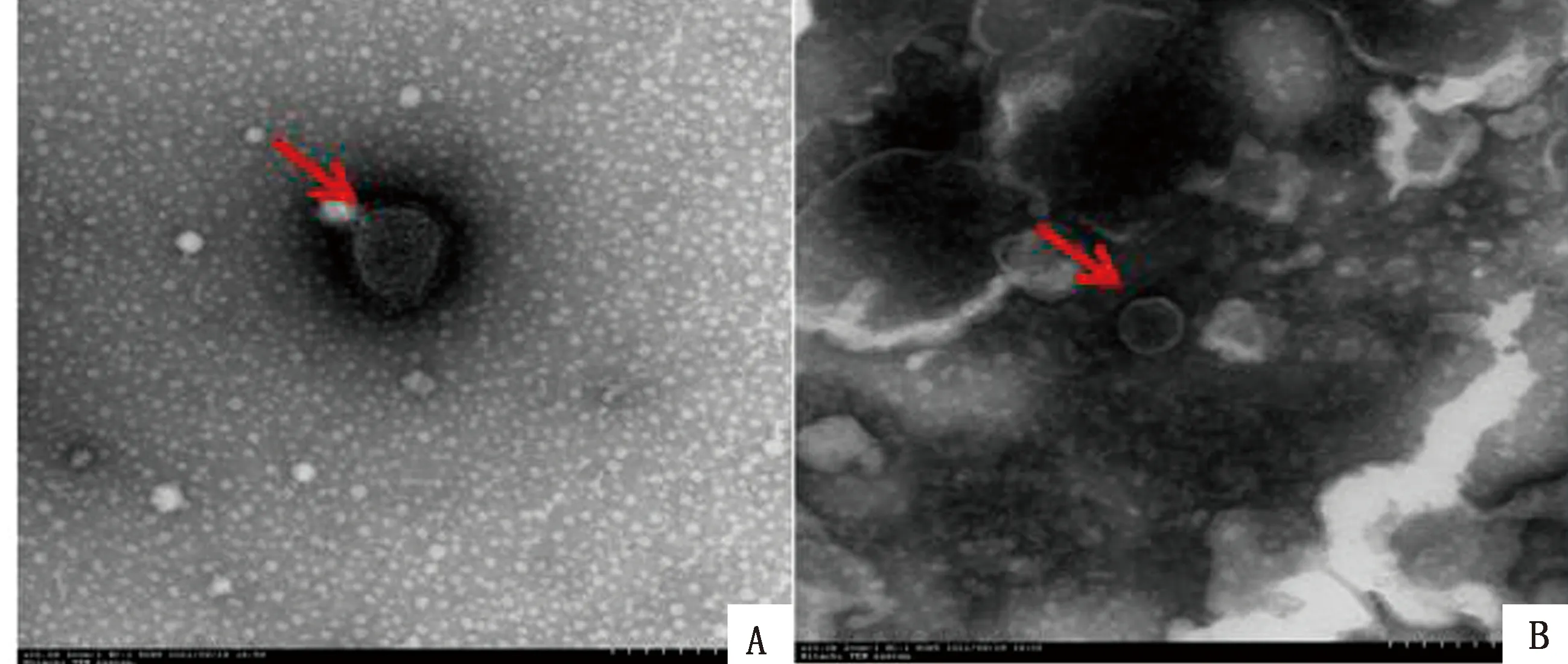
A.野生NDV NA-1株;B.BCSP31-L7/L12-IL-2
2.4.2BCSP31-L7/L12-IL-2 cVLPs Western blot分析 将经过纯化的BCSP31-L7/L12-IL-2 cVLPs 进行Western blot 鉴定,结果显示可分别与NDV M、BCSP31、L7/L12及IL-2多克隆抗体结合且条带与预期的大小相符,NDV M、L7/L12、IL-2、BCSP31蛋白大小分别为 40,20,25,36 kDa(图7)。
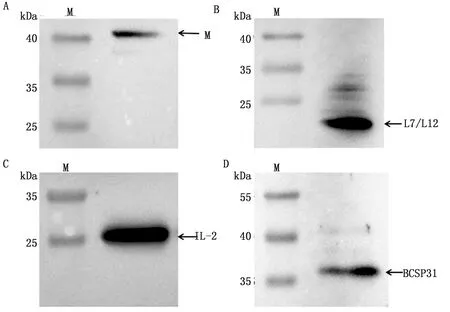
M.蛋白Marker;A.BCSP31-L7/L12-IL-2 cVLPs M蛋白;B.BCSP31-L7/L12-IL-2 cVLPs L7/L12蛋白;C.BCSP31-L7/L12-IL-2 cVLPs IL-2蛋白;D.BCSP31-L7/L12-IL-2 cVLPs BCSP31蛋白
3 讨论
理想的布鲁菌疫苗应具有下述特点[10]:(1)不会产生干扰血清学诊断的抗体;(2) 能诱导机体产生较强的Th1型免疫反应;(3)具有稳定性、易生产和便于操作特点;(4)毒性减弱且可使免疫动物感染,但对人不能有致病性;(5) 能够对妊娠动物产生长期保护效果且不会使其发生空怀、流产、死胎等症状;(6)再次免疫时不会出现血清转换。中国农业科学院哈尔滨兽医研究所联合兰州兽医研究所等单位以传统M5-90疫苗为基础研发了可区分自然感染与野毒感染的bp26基因缺失标记活疫苗产品,但现有常用布鲁菌传统疫苗仍主要是减毒活疫苗,在免疫时存在诸多不足。首先:现常用疫苗A19、S2等是将筛选出的布鲁菌菌株连续传代致弱获得的减毒活疫苗,但这种弱毒株制得的活苗仍具有残余毒力,在免疫过程中可能具有危险性,出现感染人或免疫动物患病的问题;第二,连续传代致弱获得毒株作为疫苗在免疫中易出现毒力返强的问题;第三,现有常用布鲁菌疫苗株在免疫后会刺激机体产生阳性血清[11],引起机体血清中的抗体持续性存在,导致无法区分疫苗感染与野毒感染,进而干扰血清学诊断,对布鲁菌病的确诊造成了不良的影响,不利于流行病学调查,更不利于种畜布病净化;第四,在我国所用的布病疫苗中,使用的菌株在连续传代致弱后仍有毒力,例如M5是目前我国使用布鲁菌疫苗中毒力最强的菌株,这导致在免疫过程中,可能会引起免疫动物流产乃至不孕[12];第五,现常用传统疫苗如羊种Rev-1疫苗菌株分离自羊,主要用来预防羊的布鲁菌病,牛种S19疫苗菌株分离自牛,被广泛用于预防牛的布鲁菌病,尚未存在各种属间通用的布鲁菌疫苗。由此,研发一种绿色、安全、高效、“一针多防”、可实现多方位免疫且在各菌种间高度保守的布鲁菌新型疫苗——布鲁菌嵌合VLPs,对布鲁菌病的防制起着重要作用。
L7/L12蛋白为布鲁菌核糖体蛋白,因其保守性高以及可引起机体产生良好免疫原性[13],常被应用于制作布鲁菌亚单位疫苗中。目前已有研究表示,该蛋白不仅可促使机体产生细胞免疫,同时还可促进Ⅱ型干扰素的转录与表达,发挥吞噬巨噬细胞中细菌的功能[14]。BCSP31蛋白作为布鲁菌表面蛋白,在除绵羊附睾布鲁菌外,其他菌种中均可高度保守表达,同源性高达99.3%[15],且现有研究表明,BCSP31蛋白作为布鲁菌保护性抗原,可引起机体产生保护性免疫应答,已有多种研究将其作为研发亚单位疫苗或开发新型疫苗的候选抗原[16-17]。因此,与布鲁菌传统疫苗相比,以ND VLPs为基础,上述2种蛋白为候选抗原研发的BCSP31-L7/L12-IL-2 cVLPs可以打破诸多布鲁菌传统疫苗局限性问题。首先,VLPs与纳米级病毒大小相似,是一种纳米级颗粒,拥有多种疫苗给药方式,可以通过皮下注射及肌肉注射给药[18],也可以从灌服和滴鼻等方式通过黏膜部位免疫;第二,与全病原疫苗相比,VLPs具有与病毒颗粒类似的空间结构,可以保护自身天然结构不被降解,可作为外源性抗原被树突状细胞等抗原递呈细胞摄取,交由MHCⅡ类分子进行抗原处理和提呈,刺激CD4+T辅助细胞。也可以作为内源性抗原潜伏在树突状细胞胞浆中,由MHCⅡ类分子进行抗原处理和提呈,从而刺激CD8+T辅助细胞[19],这种机制确保了机体能够产生全面且有效的免疫反应;第三,与传统弱毒疫苗相比,VLPs不含核酸,无法进行自我复制[20],不会出现毒力返强、持续性带毒、向外界排毒的现象,杜绝出现感染人或免疫动物患病的问题,不仅是一种高效的疫苗,还是一种安全、绿色、环保的疫苗;第四,VLPs通过引入保护性抗原构建,不使用全病原,可通过建立针对cVLPs不含有的血清学诊断性抗原开发ELISA检测方法,从而区分野毒感染抗体和cVLPs疫苗免疫抗体。
本研究成功构建了在各菌种间高度保守且绿色、安全、高效、可实现多方位免疫的BCSP31-L7/L12-IL-2 cVLPs,为布鲁菌新型疫苗的研发提供了新思路。但本研究构建的二联嵌合VLPs疫苗免疫原性、免疫效果以及引入的IL-2 是否会起到增强免疫效果的作用仍有待进一步研究。
猜你喜欢
山东畜牧兽医(2022年10期)2022-12-06 03:52:34
当代水产(2021年7期)2021-11-04 08:17:54
灾害医学与救援(电子版)(2018年1期)2018-06-05 02:15:36
中国兽医杂志(2016年5期)2016-06-27 01:44:58
兽医导刊(2016年12期)2016-05-17 03:51:29
广东海洋大学学报(2015年4期)2016-01-13 08:39:33
华南农业大学学报(2015年5期)2015-12-04 03:04:38
西南医科大学学报(2015年1期)2015-08-22 13:01:46
医学研究杂志(2015年6期)2015-07-01 17:41:11
癌变·畸变·突变(2015年3期)2015-02-27 06:15:07
