
中国荷斯坦奶牛LBP基因多态性分析及其与乳品质性状相关性
2023-07-08 12:34:52李嘉灵陈佳怡于海滨赵志辉
中国兽医学报 2023年5期
李嘉灵,陈佳怡,于海滨,姜 平,赵志辉
(广东海洋大学 滨海农业学院,广东 湛江 524088)
中国荷斯坦奶牛是我国重要的奶牛品种,经过十多年的选种选育,现已是我国重要的高产乳用品种之一。张胜利等[1]指出,至2020年,我国奶牛单产水平已达8 300 kg,接近欧洲一些发达国家,在奶牛育种工作加持下,奶业迅速发展。但是在奶牛单产提升的同时,目前我国对牛奶质量的研究并不深入。马广旭等[2]于2020年以6省的城乡居民为调研对象进行居民的牛奶消费状况调查,提出我国应研发低乳脂和高乳脂的乳制品,针对性开发适合特殊人群饮用的乳产品,以提高居民牛奶消费量。因此,对奶牛的相关经济性状进行深入研究具有重要的经济价值。
LBP是脂质转移蛋白家族成员之一,由中心结构域连接的N端和C端结构域组成[3]。1986年TOBIAS从急性期兔血清中纯化得到LBP。LBP的合成场所主要在肝细胞,并与细胞表面受体结合,启动先天宿主反应。脂多糖(lipopolysaccharide,LPS)通过作用于动物细胞而发挥其毒性,LBP与LPS结合形成高亲和力复合物控制LPS的反应[4]。白细胞分化抗原14(cluster of differentiation 14,CD14)可与LPS/LBP复合物结合,介导LPS对细胞的刺激作用[5]。LPS-LBP-CD14进一步激活Toll样受体4(Toll-like receptors 4,TLR4),释放促炎因子和趋化因子,诱导炎症反应[6]。SONG等[7]通过建立小鼠脓毒症模型验证得出LBP通过LBP-PPARα-CYP4a2信号通路使得肝线粒体免受LPS诱导的损伤。已有多种研究表明,LBP是内毒素血症的标志物[8-10]。
对LBP早期的研究多集中于免疫、炎症反应方面,随着对LBP的深入研究,发现LBP还影响着机体许多代谢活动。LBP是BPI家族的重要成员,参与脂代谢,从而参与心血管疾病的发生机制[11]。MORENO-NAVARRETE等[12]通过脂质组学分析敲除LBP后脂肪细胞的脂质组学特征,发现敲除LBP后脂肪细胞的脂质体发生急剧变化。LATORRE等[13]通过手术干预注射shRNA-LBP慢病毒颗粒,构建特定脂肪组织LBP基因敲低的模型小鼠,发现基因敲低后,小鼠血浆游离脂肪酸、瘦素、甘油三酯降低,脂肪增加相关基因表达量减少。MOLINARO等[14]发现下调LBP可以调节非肥胖小鼠的全身葡萄糖稳态。SUN等[15]通过调查发现肥胖受试者的体内LBP水平显著高于体质量正常受试者,LBP水平的升高与2型糖尿病、代谢综合征及肥胖有关。
目前的研究表明LBP多态性与炎症反应息息相关。ZENG等[16]研究发现rs2232618多态性与败血症和多器官功能障碍的更易感呈显著相关。CASTANO-RODRIGUEZ等[17]研究发现LBP多态性与幽门螺旋杆菌感染的胃癌有明显关联。程结南[18]发现LBP多态性与荷斯坦奶牛临床型乳腺炎发病率呈显著相关。然而与奶牛奶品质性状的多态性研究鲜有报道。因此本研究以荷斯坦奶牛为群体,采用直接测序法研究LBP单核苷酸多态性,并分析SNPs位点与奶牛乳品质性状之间的相关性,为后期验证基因功能奠定基础,同时提供优质奶牛选种选育分子标记。
1 材料与方法
1.1 样品采集饲养环境相同的133头中国荷斯坦奶牛来自黑龙江某牛场,牛颈静脉采血(5 mL/头)于抗凝管中,置于-80℃保存。
1.2 主要试剂血液DNA提取试剂盒购自天根生化有限公司;2×Taq Master Mix(Dye Plus)、50×TAE电泳缓冲液购自南京诺唯赞生物科技股份有限公司;琼脂糖购自北京全式金生物技术有限公司;DL2000 DNA Marker购自北京宝日医生物技术有限公司。
1.3 基因组DNA提取根据试剂盒说明书提取荷斯坦奶牛血样DNA,使用Thermo ScientificTMNanoDrop Lite分光光度计检测DNA的浓度和纯度,用1%琼脂糖电泳检测DNA提取效果,-20℃保存。
1.4 引物设计根据Ensembl中公布的牛LBP基因序列(ENSBTAT00000022428.3),利用Primer 5.0软件设计引物,引物序列见表1。设计好的引物由生工生物工程(上海)股份有限公司合成。

表1 LBP基因扩增目的片段引物序列及相关信息
1.5 PCR扩增目的条带PCR反应体系:2×Taq Master Mix(Dye Plus)10 μL,上、下游引物各0.5 μL,基因组DNA 1 μL,ddH2O 8 μL。PCR反应程序:95℃预变性5 min;95℃变性30 s,60℃退火30 s,72℃延伸1 min,进行30个循环;72℃延伸5 min。PCR产物置于4℃保存。PCR产物经1.5%琼脂糖凝胶电泳检测扩增效果,若条带单一、片段大小正确,送生工生物工程(上海) 股份有限公司测序。
1.6 性状测定根据中华人民共和国农业行业标准中《中国荷斯坦牛生产性能测定技术规范》采集样品,牛乳中脂肪、蛋白、乳糖、体细胞、干物质的测定采用乳成分分析仪进行检测。

2 结果
2.1 PCR扩增LBP基因目的片段以基因组DNA为模板,通过PCR扩增LBP目的片段,产物经1.5%琼脂糖凝胶电泳检测,结果见图1,2,可见PCR产物条带单一清晰、特异性好。参考DL2000 DNA Marker标识,所示条带长度与目的条带相符。

M.DL2000 DNA Marker;1~7.分别为不同个体PCR扩增目的片段

M.DL2000 DNA Marker;1~2.分别为不同个体PCR扩增目的片段
2.2 LBP目的片段测序结果测序中共发现3个明显套峰,分别位于第二内含子5′端第1 095个碱基,由G突变为A;第五内含子5′端第142和199位碱基,第142位碱基由T突变为C,第199位碱基由A突变为G。这3个SNP为I2-1095 G>A、I5-142 T>C、I5-199 A>G(图3); 3个SNPs不同基因型测序结果见图4,5,6。

图3 牛LBP基因混池DNA扩增目的片段测序结果

图4 牛LBP基因I2-1095 G>A位点3种基因型测序结果

图5 牛LBP基因I5-142 T>C位点3种基因型测序结果
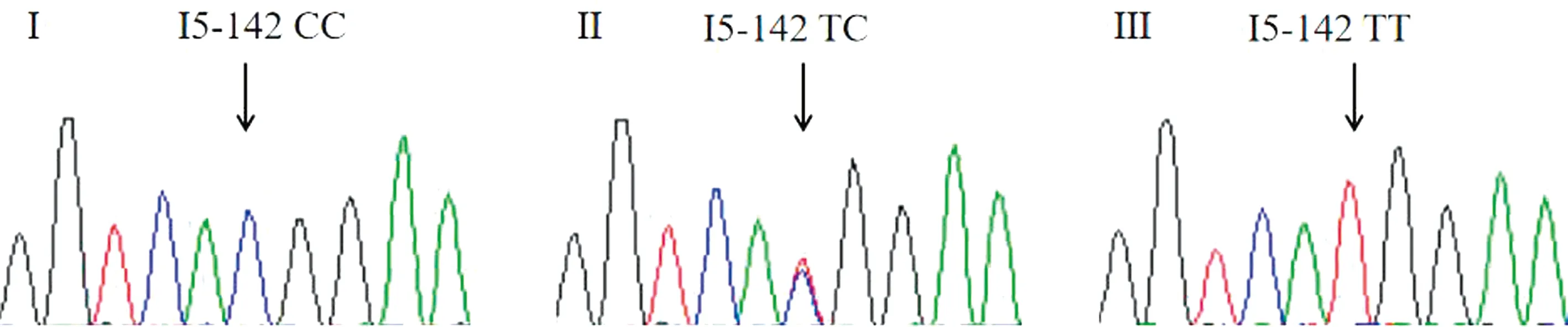
图6 牛LBP基因I5-199 A>G位点3种基因型测序结果
2.3 LBP基因3个SNP位点的群体遗传特性由表2可知,I2-1095 G>A位点的优势基因为G,其频率为0.654 1,该位点优势基因型为GG,频率为0.443 6;I5-142 T>C位点的优势基因为T,其频率为0.556 4,该位点的优势基因型为TC,其频率为0.496 2;I5-199 A>G位点的优势基因为G,其频率大于0.518 8,该位点的优势基因型为AG,其频率为0.481 2。 3个SNPs的PIC和H均介于0.25与0.50之间,说明该SNPs在群体中处于中度多态。χ2验证结果显示, 3个SNPs位点在荷斯坦奶牛群体中处于Hardy-Weinberg平衡状态(P>0.05)。
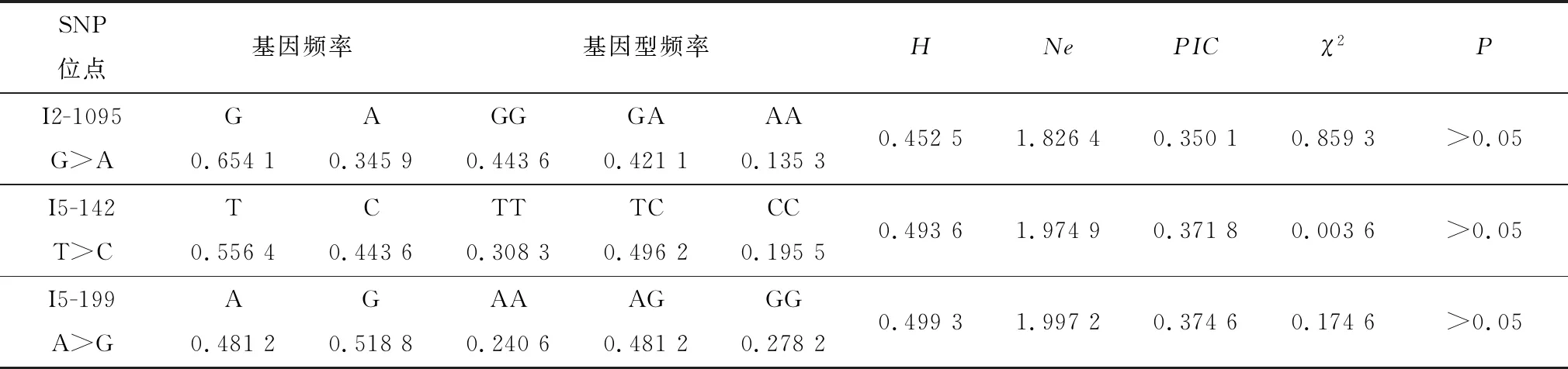
表2 牛LBP基因SNPs群体遗传特性
2.4 LBP基因SNPs位点不同基因型与中国荷斯坦奶牛乳品质性状的相关性分析在133个待测奶牛样品中, 3个SNPs位点不同基因型的产奶量、乳脂、乳蛋白、乳糖、干物质、体细胞、尿素氮、校正奶8个指标的数据平均值如表3。结果显示,I2-1095 G>A位点与乳品质性状无显著相关(P>0.05),I5-142 和I5-199的SNPs与奶牛乳品质性状中的乳糖、乳脂、体细胞、校正奶都呈显著相关(P<0.05),此外,I5-142 T>C位点还与产奶量呈显著相关(P<0.05)。I5-142 T>C、I5-199 A>G 2个SNPs与蛋白、干物质均无显著性差异(P>0.05)。
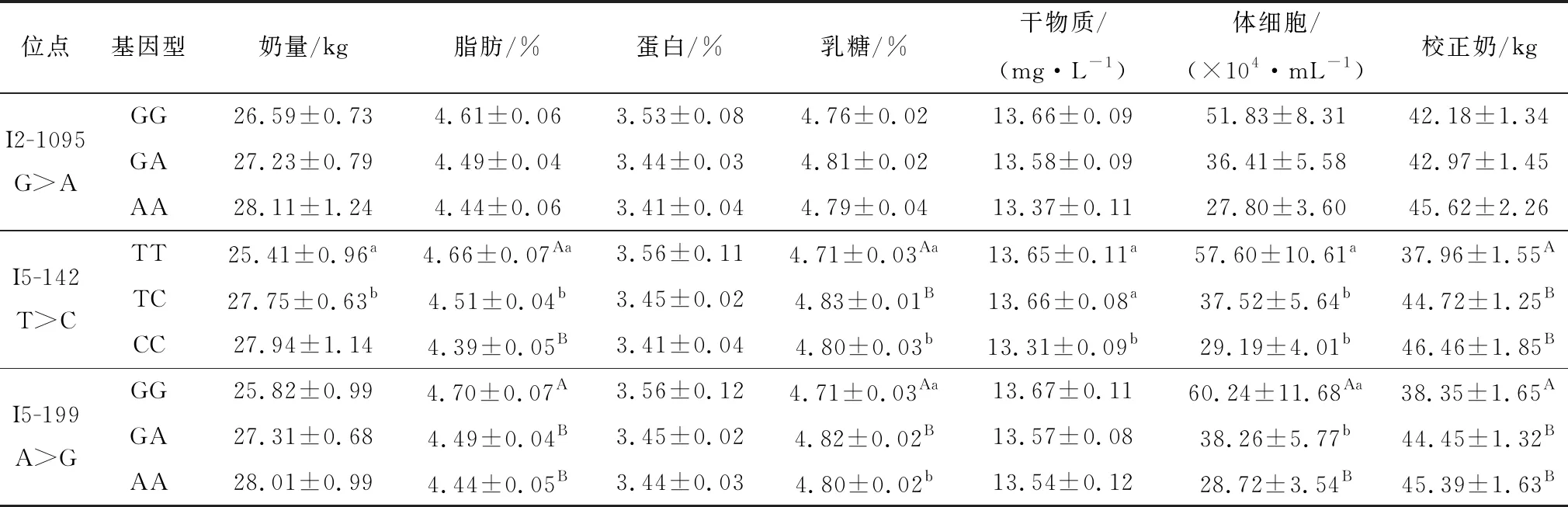
表3 牛LBP基因SNPs 3种基因型与中国荷斯坦奶牛生产性状的相关性分析
I5-142 T>C位点TC基因型个体的产奶量显著高于TT基因型(P<0.05);脂肪含量方面,TT型个体乳脂含量显著高于TC型个体(P<0.05),极显著高于CC型(P<0.01);TT型个体乳糖及校正奶含量极显著小于TT、TC型个体(P<0.01);牛奶中体细胞的数量提示奶牛乳房感染情况,TT基因型个体的体细胞数量显著高于TC、CC型个体(P<0.05)。
I5-199 A>G位点GG基因型个体的乳脂含量极显著高于GA、AA型个体(P<0.01);GA基因型的乳糖含量最高,极显著高于GG型个体(P<0.01);体细胞方面,GG型个体体细胞数最高,显著高于GA型(P<0.05),极显著高于AA型(P<0.01);GG型个体的校正奶含量极显著小于GA、AA型。
2.5 LBP基因单倍型与中国荷斯坦奶牛乳品质性状的相关性分析对LBP 3个SNPs进行单倍型分析,单倍型频率见表4,共有5种单倍型组合,分别为H1H1、H1H2、H1H3、H2H3、H2H2。对 3个SNPs位点进行连锁不平衡分析,发现I5-142 T>C、I5-199 A>G 2个SNPs存在强连锁遗传,见图7。把5种单倍型与奶牛乳品质性状进行相关性分析(表5),结果显示H1H2和H2H2单倍型个体产奶量显著高于H1H3(P<0.05);H1H1型乳脂含量极显著高于H1H2、H2H2型单倍型(P<0.01),显著高于H1H3型个体(P<0.05);H1H2型个体含乳糖最高,极显著高于H1H1型(P<0.01);H2H3型个体干物质含量最高,极显著高于其他单倍型(P<0.01);3种单倍型中,H1H1型个体含有数量最多的体细胞,显著高于H1H2、H2H2单倍型(P<0.05);H1H2型个体的校正奶含量极显著高于H1H1型(P<0.01),显著高于H1H3型(P<0.05)。3种单倍型与蛋白含量无显著相关(P>0.05)。
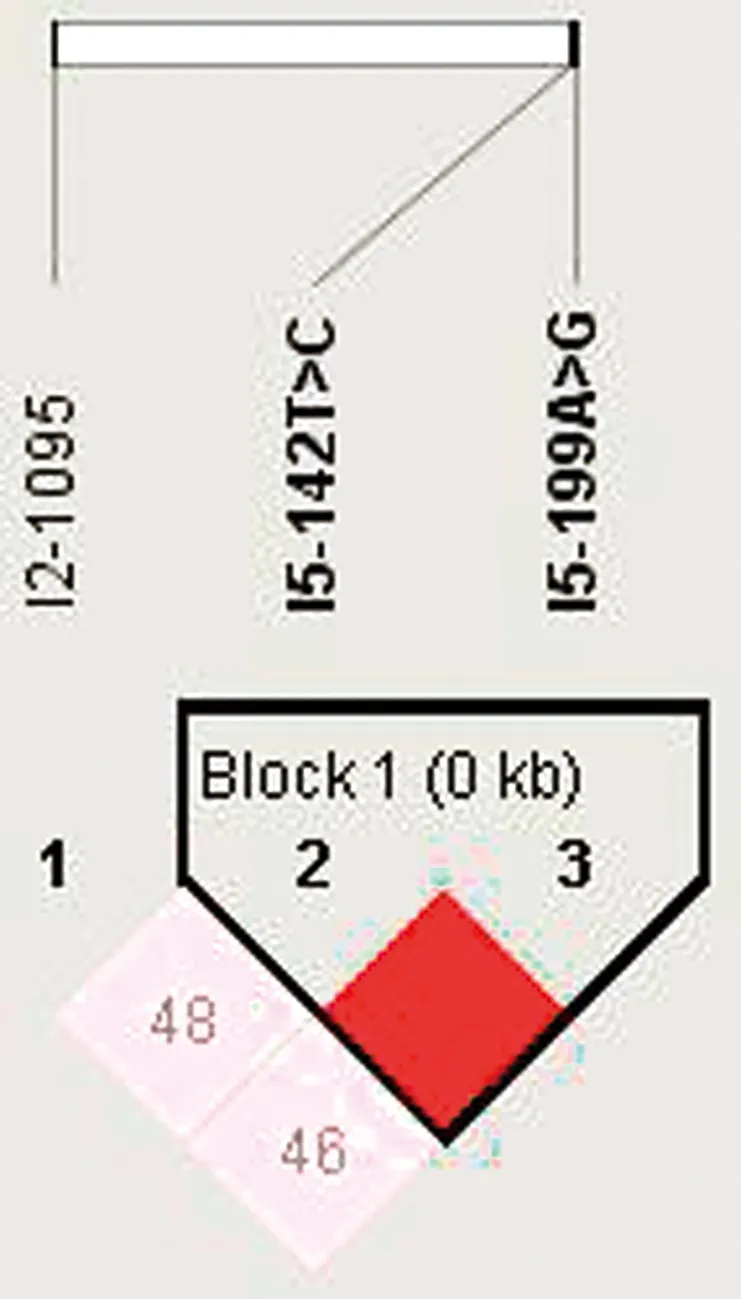
图7 牛LBP基因SNPs位点连锁分析

表4 牛LBP基因单倍型频率

表5 牛LBP基因单倍型与中国荷斯坦奶牛生产性能相关性分析
3 讨论
我国对高质量奶的需求与日俱进,同时对进口乳制品的依赖愈演愈烈。刘馨蔚[19]指出,我国奶业消费水平持续增长,但增长份额更多来自国外生鲜乳原料供应,截止2020年,我国奶源自给率仅为66%。这将使我国奶业竞争力置于不利地位,提高我国乳制品质量及产量从而提升我国消费者对乳制品的信心尤为重要。
LBP来源于肝脏,是一种急性期蛋白,主要参与调节革兰阴性菌引起的免疫反应。而革兰阴性菌是奶牛临床乳腺炎常见的病原体,感染后可降低牛奶质量与产量,导致不可估量的经济损失。本研究发现LBP 2个SNPs均与目标奶牛群体的乳脂、乳糖、体细胞、校正奶含量呈显著相关关系,其中I5-142 T>C位点还与产奶量显著相关。目前关于牛LBP多态性的报道多以乳腺炎、乳房炎为主。CHENG[20]采用PCR-SSCP方法对奶牛LBP基因进行SNP分析,发现g.4619G → A位点GA基因型个体临床乳腺炎发病率显著高于其他两种基因型。SUN等[21]发现牛重组LPS结合蛋白(BRLBP)可消减LPS诱导的BMEC炎症损伤。ZENG等[22]发现患有临床乳腺炎奶牛的牛奶中体细胞数量是健康奶牛牛奶体细胞的9倍,且牛奶中LBP含量高于健康群体,血液中LBP含量显著高于健康组。牛奶中的体细胞数提示着乳房或乳腺的炎症反应程度,固LBP可能与体细胞数有关,这也印证了本研究中LBP多态性与体细胞呈显著相关。
TSUGAMI等[23]和KOBAYASHI等[24]用LPS处理BMECs,发现LPS处理后会降低β-酪蛋白、乳糖和甘油三酯的分泌,并且LPS处理后细胞表达STAT3和NF-κB显著增加,STAT3和NF-κB信号通路的激活会抑制乳腺产奶量,从而说明LPS对产奶量有影响。LBP通过与LPS形成高亲和力复合体参与机体的调节活动,与LBP结合是LPS转移级联的基本步骤[3]。LBP与LPS的结合能力也从侧面提供了LBP与产奶量、乳脂、乳糖的相关性信息。WANG等[26]发现TLR4 c.-226 G >C和 c.2021 C >T这2个SNPs位点与奶牛产奶量有显著关系。CHRISTINE等[27]发现 CD14-1908 A 等位基因与乳脂和蛋白质产量存在相关关系。根据TLR4、CD14是LBP下游基因的关系,推断LBP多态性与产奶量、乳脂也存在联系。目前尚无研究报道牛LBP基因是否影响乳糖调节,LBP参与糖代谢的研究集中在葡萄糖方面。MOLINARO等[14]使用CRISPR/CAS9技术敲低了肝脏中LBP,发现LBP可调节机体葡萄糖稳态。本研究首次研究牛LBP多态性对奶牛生产性状的影响,并发现LBP 2个SNPs与产奶量、乳脂、乳糖、体细胞、校正奶有显著影响。
对LBP基因 2个SNPs进行连锁不平衡分析,结果显示SNPs强连锁遗传,共有3种单倍型,且与奶牛生产性状显著相关。把SNPs 3种单倍型与生产性状进行相关性分析,结果提示体细胞数与脂肪、乳糖、校正奶呈负相关,与干物质呈正相关。H1H1型个体体细胞数超过《中国奶业D20标准生牛乳》中的体细胞数量,提示H1H1型个体患有乳房疾病的风险增加。H1H2、H2H2型个体在乳脂、乳糖、干物质、校正奶方面并没有显著性差异,结合各项指标的平均值,提示H1H2单倍型可能为最优选择。该单倍型可作为奶牛选种选育的分子标记,可提高奶牛生产性能。
本研究发现牛LBP基因I2-1095 G>A、I5-142 T>C、I5-199 A>G 3个SNPs对中国荷斯坦奶牛部分经济性状产生显著影响,分别有产奶量、乳糖、乳脂、校正奶和体细胞数。这些SNPs为将来奶牛的生产性状辅助选择提供有价值的分子标记,并丰富了奶牛选种选育的基因资源,亦为日后对LBP进行深入的功能研究奠定了基础。
猜你喜欢
现代畜牧科技(2021年11期)2021-12-21 06:11:16
今日农业(2020年24期)2020-12-15 16:16:00
科学(2020年4期)2020-11-26 08:27:10
中国乳业(2019年12期)2020-01-17 08:47:40
分析化学(2018年4期)2018-11-02 10:16:04
中西医结合心脑血管病杂志(2016年20期)2016-03-01 04:20:45
上海农业学报(2016年5期)2016-02-10 06:52:55
中国粮油学报(2016年5期)2016-01-23 02:44:57
华南农业大学学报(2015年5期)2015-12-04 03:04:38
畜牧兽医学报(2015年3期)2015-07-05 08:22:16
