
不同土壤含水量下沙棘叶片解剖结构研究
2023-07-07 02:02崔晓晗叶丽红贾文龙东保柱
北方农业学报 2023年2期
崔晓晗 ,叶丽红 ,2,贾文龙 ,东保柱 ,张 敏 ,宋 阳 ,2
(1.内蒙古农业大学园艺与植物保护学院,内蒙古呼和浩特 010010;2.内蒙古自治区设施园艺工程技术研究中心,内蒙古呼和浩特 010010;3.内蒙古淳点实业有限公司,内蒙古鄂尔多斯 017000;4.固阳县农村社会事业发展中心,内蒙古固阳 014200)
沙棘(Hippophae rhamnoides L.)是一种胡颓子科沙棘属落叶灌木,广泛分布于我国华北、东北、西北等地区,为内蒙古、陕西、辽宁等地创造了巨大的经济效益[1]。沙棘果实营养丰富,含有多种维生素、脂肪酸、类黄酮等营养物质,其种子、叶片等其他器官中亦含有活性物质,为药食同源植物,极具药用价值、营养价值及经济价值,因此近年来愈发受到人们关注,发展前景广阔[2]。沙棘对环境适应性强,具有耐旱、抗风沙、耐盐碱、耐瘠薄等优质特性,常被用于水土保持、防风固沙等生态环境治理中[3]。水是贯穿植物整个生命周期中不可或缺的因素,植物长期处于水分过多或亏缺环境中皆会受到不可逆转的伤害。沙棘作为对环境适应能力较强的植物,在旱涝环境中可做出积极响应,快速适应环境。研究发现,随着干旱程度加重,沙棘叶片中超氧化物歧化酶及过氧化物酶活性先升高后降低,可溶性糖及脯氨酸积累增加,这些变化表明沙棘可在一定程度上抵御干旱[4]。南吉斌等[5]比较西藏3种沙棘属植物干旱胁迫下的光合特性得出,干旱胁迫使3种沙棘属植物光合效率均有不同程度的降低,其中江孜沙棘光合作用受干旱抑制最小,抗旱性最强。胡建忠等[6]比较5个沙棘无性系对干旱的响应发现,其中3个品种的长期水分利用率及抗氧化酶活性在重度干旱下依然持续升高,表现出对长期干旱较强的抗逆性。目前,关于沙棘在干旱下光合及生理生化方面和不同品种抗旱性评价的报道较多,但缺乏有关不同土壤含水量下沙棘叶片结构的研究。本试验通过观察不同土壤含水量下沙棘叶片解剖结构的变化情况,研究沙棘叶片结构对不同土壤含水量的响应,以期为进一步深入研究沙棘抗旱耐涝机制提供参考。
1 材料和方法
1.1 试验材料
以一年生中国沙棘(Hippophae rhamnoides L.subsp.sinensis Rousi)扦插苗为试验材料。
1.2 试验方法
试验于2021年4—9月,在内蒙古农业大学教学基地日光温室中进行。4月10日,将一年生枝条扦插于塑料花盆中(内径15 cm、深20 cm),填充砂壤土。常规管理(每隔3 d浇水2 L)至6月30日后开始处理。设置干旱(采取自然干旱方式,每隔7 d浇水2 L)、水涝(每日浇水2 L,花盆底部置于装水托盘中,并使托盘内水面高度始终保持3 cm以上)2个处理,以常规灌水(每隔3 d浇水2 L)为对照(CK),每个处理10株。处理至8月30日结束,选取长势一致、品质较好的植株,取每一株同一层枝条上靠近顶端生长点展平的上层叶片作为幼叶,以及下层靠近茎基部的老叶。每个处理老叶与幼叶各取10片,参照温国等[7]的制片方法,叶片中部取样并切成包含主叶脉的5 mm×5 mm小方块置于FAA固定液中进行2次低压固定,放置48 h后,进行石蜡切片制片。根据常规石蜡切片流程,依次进行脱水、透明、浸蜡、包埋、修块、切片、展片、脱蜡、染色、封片。用番红-固绿染色,加拿大树胶封片。完成制片后用光学显微镜(OLYMPUS BX53,日本)观察并照相。
1.3 项目测定
用OLYMPUS cellSens Standard测量叶片厚度、上表皮厚度、下表皮厚度、栅栏组织厚度、海绵组织厚度、主叶脉直径、维管束直径、木质部导管最大直径,并计算栅海比、叶片组织结构紧密度(CTR)、叶片组织结构疏松度(SR)[8]。各指标选10张切片测量,取平均值。
栅海比=栅栏组织厚度/海绵组织厚度 (1)
CTR/%=(栅栏组织厚度/叶片厚度)×100 (2)
SR/%=(海绵组织厚度/叶片厚度)×100 (3)
1.4 数据处理
用Excel 2007软件整理数据,用IBM SPSS Statistics 26.0统计学软件进行统计分析。
2 结果与分析
2.1 不同土壤含水量下沙棘叶片及其表皮结构变化
由图1可知,沙棘叶片由表皮、叶肉及叶脉构成,上、下表皮细胞均为单层,近似长方形排列于叶肉细胞外侧。上表皮具有角质,下表皮着生表皮毛。叶肉细胞分化为栅栏组织和海绵组织,栅栏组织位于近轴端,排列整齐紧密。海绵组织位于远轴端,排列松散不规则,其中具有空腔气室。叶脉中含有单一维管束,由木质部、韧皮部、厚角组织及薄壁组织构成。维管系统健壮发达;木质部导管位于上表皮一端,径向排列;韧皮部位于木质部外侧,靠近下表皮。干旱条件下的沙棘叶片表皮细胞排列较紧密,可减少叶片内部水分散失,而水涝条件下的表皮细胞排列略疏松。

图1 不同土壤含水量下沙棘幼叶及老叶的叶片结构Figure 1 Leaf structure of seabuckthorn young and old leaves under different soil moisture content
由图2可知,干旱条件下沙棘幼叶与老叶的叶片厚度、上表皮厚度及下表皮厚度与CK相比均显著(P<0.05)增加。水涝条件下沙棘幼叶的叶片厚度显著(P<0.05)小于CK,上、下表皮厚度均显著(P<0.05)大于CK;老叶的上表皮厚度显著(P<0.05)大于CK,而叶片及下表皮厚度与CK相比无显著差异(P>0.05)。CK的老叶叶片厚度、上表皮厚度和下表皮厚度在CK下分别较幼叶增长7.28%、56.10%、33.54%,在干旱条件下分别较幼叶增长10.45%、33.66%、3.98%;在水涝条件下分别较幼叶增长33.65%、42.88%、3.92%。可见沙棘叶片及其表皮在干旱条件下均表现出增厚趋势,但其增长量小于CK,说明干旱使沙棘叶片及表皮细胞发育受到抑制。水涝条件下幼叶变薄,但表皮增厚,老叶仅上表皮增厚变化显著,叶片厚度增幅大于CK,说明水涝在一定程度上促进沙棘叶片生长发育。在3种水分条件中,幼叶与老叶的叶片厚度、上表皮厚度及下表皮厚度均表现为干旱条件下最大。
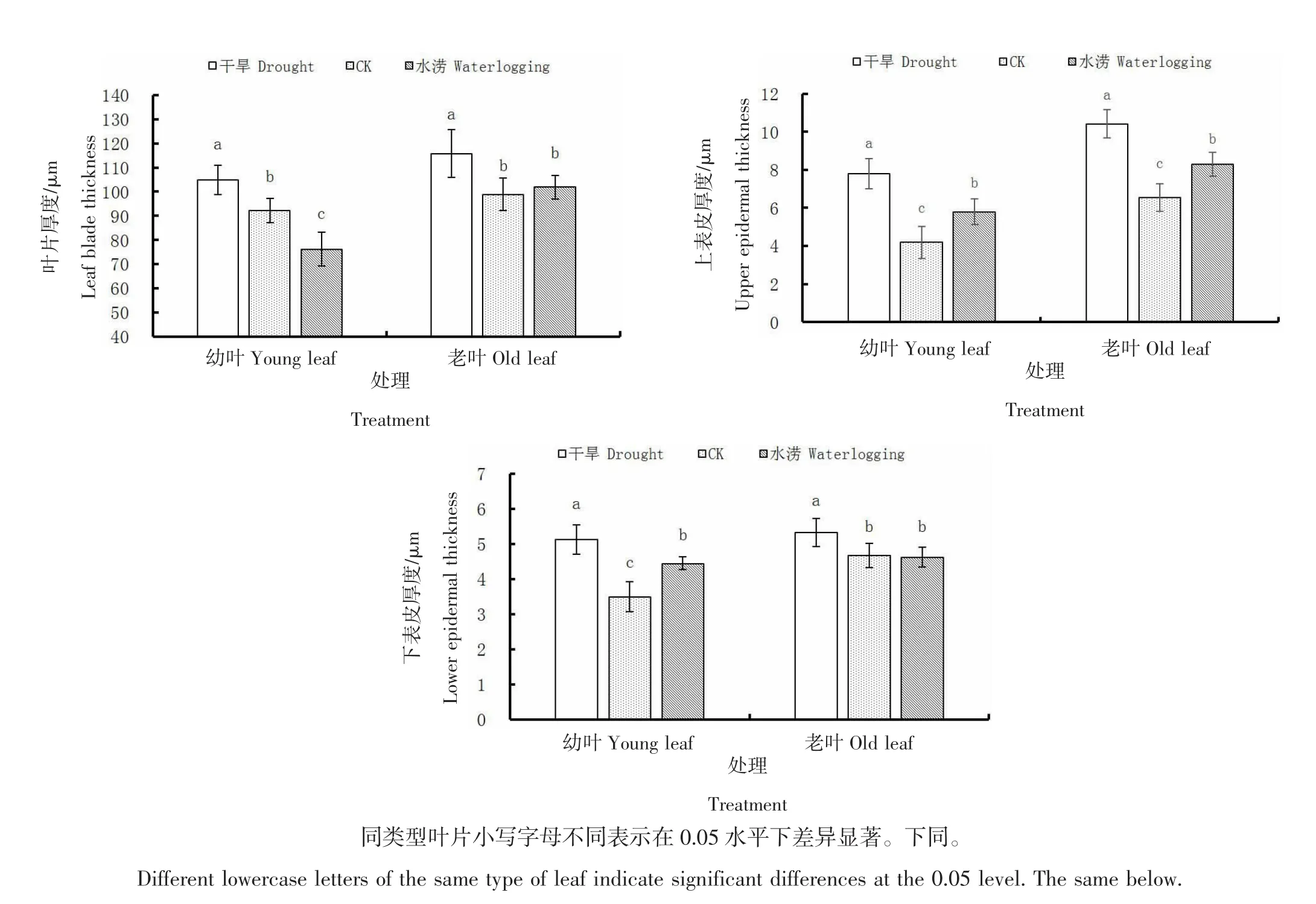
图2 不同土壤含水量下沙棘叶片及表皮厚度变化Figure 2 Changes of leaf and epidermal thickness of seabuckthorn under different soil moisture content
2.2 不同土壤含水量下沙棘叶肉结构变化
由图3可知,干旱条件下沙棘幼叶栅栏组织厚度、栅海比和CTR显著(P<0.05)大于CK,SR显著(P<0.05)小于CK,海绵组织厚度与CK差异不显著(P>0.05);老叶海绵组织厚度和SR显著(P<0.05)大于CK,栅海比和CTR显著(P<0.05)小于CK,栅栏组织厚度与CK无显著差异(P>0.05)。水涝条件下沙棘幼叶与老叶的栅栏组织厚度、栅海比和CTR均显著(P<0.05)小于CK,海绵组织厚度和SR均显著(P<0.05)大于CK。可见干旱条件下沙棘幼叶栅栏组织发达,叶肉较紧实,但干旱使栅栏组织发育退化,老叶海绵组织增厚,叶肉疏松,栅海比和CTR均较幼叶显著降低。而水涝条件对栅栏组织和海绵组织发育均有促进,但幼叶及老叶均表现为海绵组织更为发达,栅海比和CTR均为3种水分条件中最低。
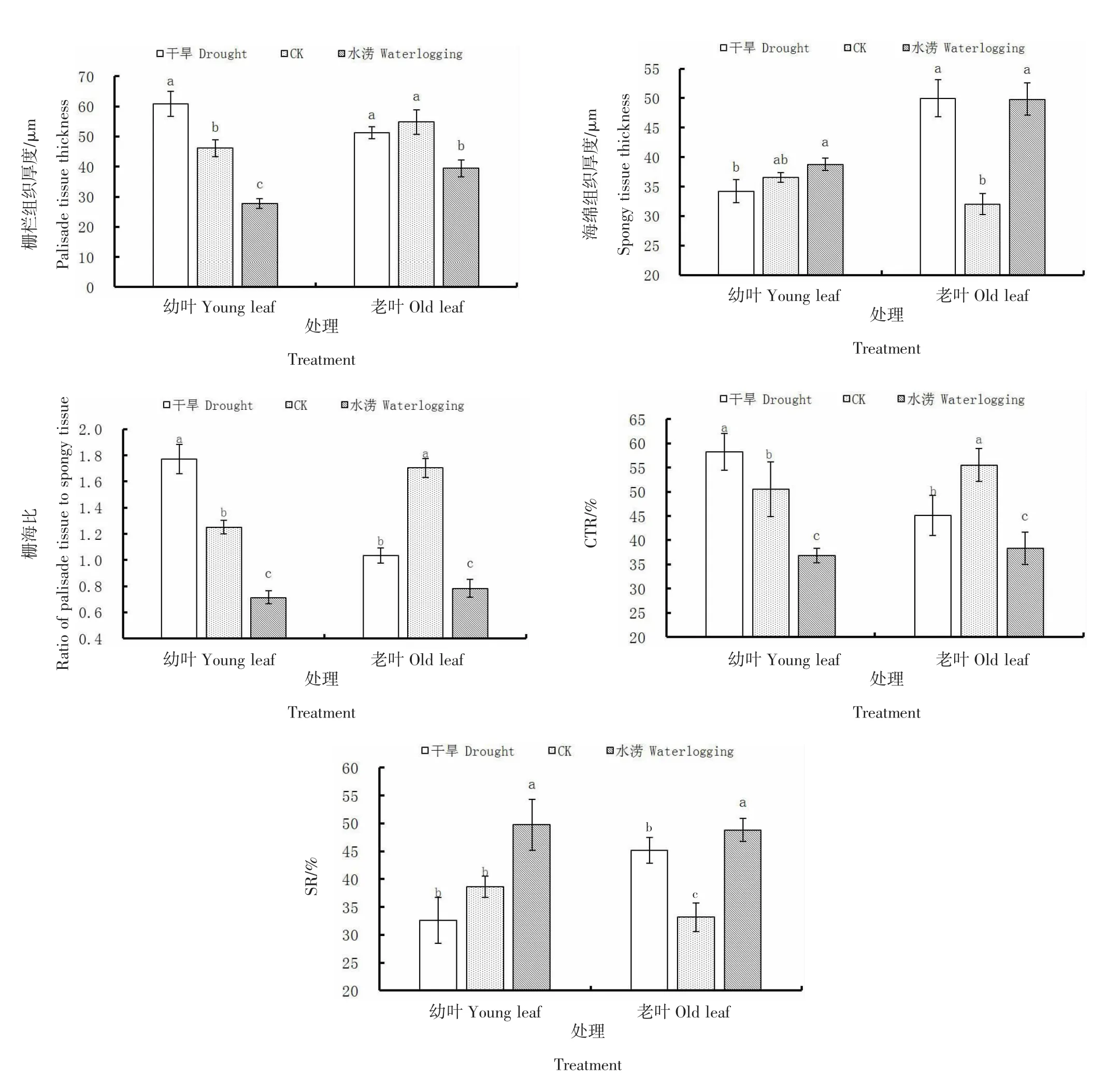
图3 不同土壤含水量下沙棘叶肉结构变化Figure 3 Structural changes of seabuckthorn mesophyll under different soil moisture content
2.3 不同土壤含水量下沙棘叶脉结构变化
由图4可知,干旱条件下沙棘叶片的主叶脉直径、维管束直径和木质部最大导管直径均表现为幼叶与CK相比无显著差异(P>0.05),老叶显著(P<0.05)大于CK。水涝条件下幼叶的维管束直径显著(P<0.05)小于CK,主叶脉直径和木质部最大导管直径与CK无显著差异(P>0.05);老叶主叶脉直径、维管束直径和木质部最大导管直径均显著(P<0.05)大于CK。CK的沙棘老叶主叶脉直径、维管束直径和木质部最大导管直径分别较幼叶增长2.19%、-6.39%、3.81%,在干旱条件下分别较幼叶增长35.51%、15.05%、38.76%,在水涝条件下分别较幼叶增长22.88%、42.52%、34.10%。可见干旱和水涝条件均使沙棘叶片的主叶脉及维管系统生长量较CK增加,且使老叶的主叶脉及维管系统显著增粗,但对幼叶无显著影响。这可能是由于幼叶距离茎基部较远,维管系统传输能力及效率较老叶弱,故对水分响应没有老叶敏感。
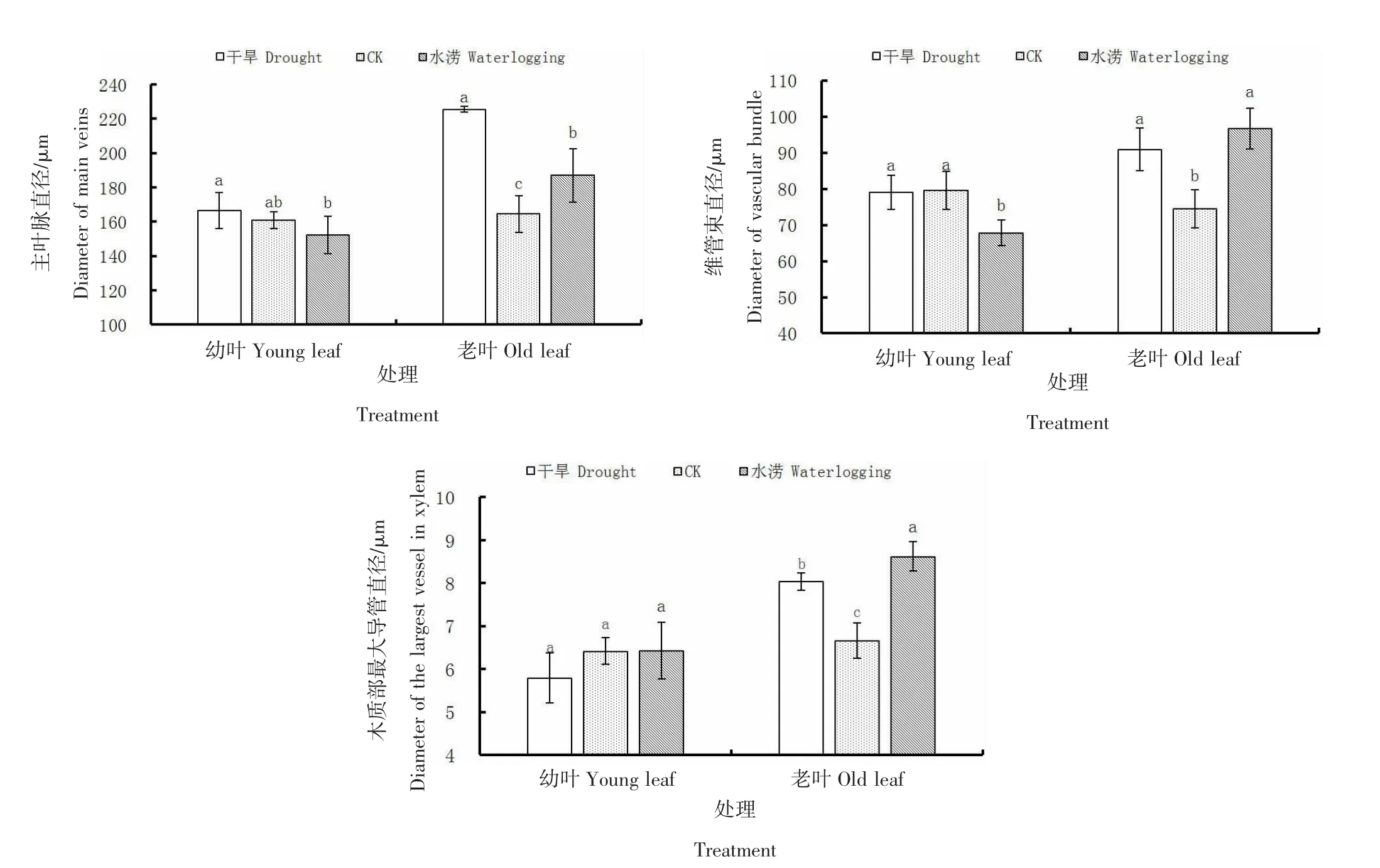
图4 不同土壤含水量下沙棘叶脉结构变化Figure 4 Changes of seabuckthorn leaf vein structure under different soil moisture content
2.4 土壤含水量和叶片类型对沙棘叶片解剖结构双因素方差分析
由表1可知,土壤含水量和叶片类型对叶片厚度、上表皮厚度及下表皮厚度均有极显著影响(P<0.01),二者交互作用对叶片厚度和下表皮厚度有显著影响(P<0.05),对上表皮厚度无显著影响(P>0.05)。叶片类型对栅栏组织厚度及海绵组织厚度有显著影响(P<0.05),对栅海比、CTR和SR无显著影响(P>0.05)。土壤含水量及二者交互作用对沙棘叶片栅栏组织厚度、海绵组织厚度、栅海比、CTR和SR均有极显著影响(P<0.01)。土壤含水量、叶片类型及其交互作用均对沙棘主叶脉直径、维管束直径和木质部最大导管直径存在显著影响(P<0.05)。
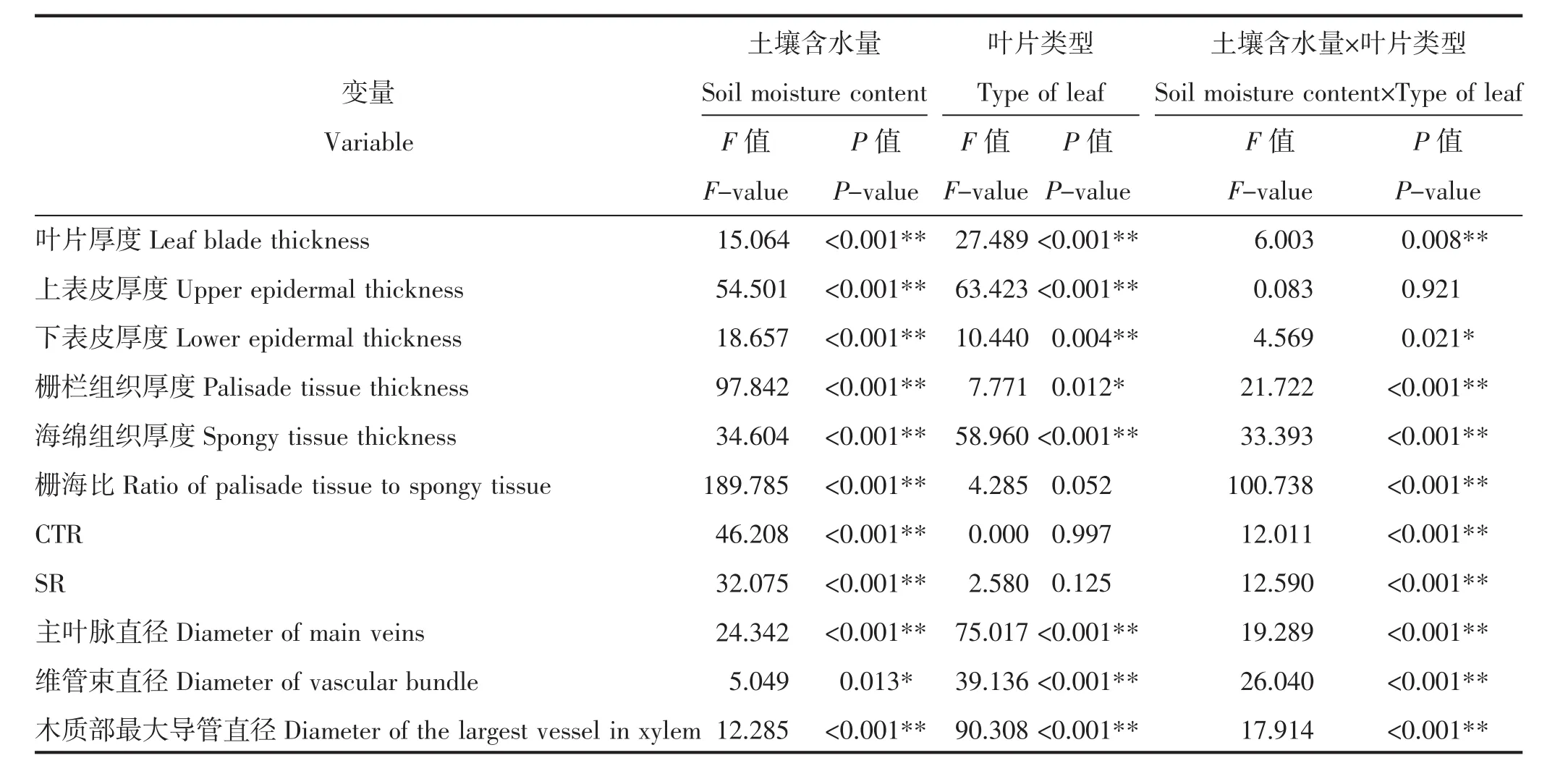
表1 土壤含水量和叶片类型对沙棘叶片解剖结构双因素方差分析Table 1 Soil moisture content and type of leaf on the anatomical structure of seabuckthorn leaves by two-way ANOVA
3 讨论与结论
叶片是植物生长过程中直接暴露于环境中的器官,故对环境温度、湿度、光照、CO2和O2浓度等变化极为敏感,且具有较强的可塑性,尤其在植物遭受水分胁迫时,叶片特征变化最为直观。当植物处于不同水分环境中,其叶片解剖结构变化可有效体现出植物对不同水分条件的响应[9-11]。
表皮是控制叶片吸收光量子的关键因素,因此在光合作用和蒸腾作用中具有重要意义[12-13]。研究表明,表皮在干旱环境下表现增厚,能够减少植物体内水分散失,使植物适应干旱环境[14-15]。刘红茹等[16-17]研究发现,表皮细胞越厚,越利于提高植物的抗旱性。较厚的表皮细胞具有一定的贮水能力[18]。徐扬等[19]分析苹果属植物叶片解剖结构发现,表皮细胞较厚的品种具有更强的抗旱能力。本试验结果表明,无论是沙棘幼叶还是老叶,处于干旱条件下,其叶片及上、下表皮均较CK显著增厚,且在3种水分条件中最大,这与于海燕等[20]研究结果一致。水涝条件下沙棘幼叶显著变薄,表皮增厚;老叶厚度无显著变化,仅上表皮增厚。
叶片栅栏组织与海绵组织的比例可间接反映环境中的水分状态[21]。发达的栅栏组织可避免叶肉细胞遭受强光灼伤,且能高效利用衍射光进行光合作用,提高光合效能,减少植物体内水分耗散[22]。栅栏组织增厚,海绵组织变薄,栅海比及CTR增加等变化,利于提高叶片光合效率,提高植物保水能力及抗旱性[21]。本试验中,观察各处理下沙棘叶片解剖结构可以发现,幼叶的栅栏组织厚度在3种水分条件下表现为干旱>CK>水涝,干旱条件下栅栏组织细胞层数多于CK,水涝条件下与CK基本一致;老叶的栅栏组织厚度在干旱条件下与CK相比差异不显著,水涝条件下显著小于CK。沙棘幼叶的海绵组织厚度在3种处理下无显著差异;老叶的海绵组织在干旱和水涝条件下均较CK增厚。观察沙棘解剖结构可见干旱条件下老叶的栅栏组织细胞排列较幼叶疏松,栅栏组织在干旱条件下较幼叶有所退化,海绵组织较幼叶增厚;水涝条件下老叶栅栏组织及海绵组织均较幼叶增厚。沙棘幼叶在干旱下栅栏组织增厚、细胞层数增加的现象体现出对干旱环境的适应;而老叶在干旱下海绵组织增厚的表现与唐军荣等[23]对干热河谷牛角瓜的研究结果相似,海绵组织细胞间隙较大有利于贮藏水分,并增加叶片内表面积,可促进光合作用的气体交换,从而提高光合效率。水涝条件下沙棘幼叶及老叶的栅栏组织厚度均显著小于CK,海绵组织厚度高于CK,以弱化栅栏组织,强化海绵组织发育适应水涝条件。栅海比能反映植物的抗旱能力,其值越高,抗旱能力越强[24-25]。本试验中,干旱条件下沙棘幼叶栅海比显著升高,老叶却显著降低。幼叶通过升高栅海比减少叶片蒸腾失水,但老叶对干旱的响应没有幼叶积极。水涝条件下幼叶和老叶的栅海比均为3种处理下最低,可见沙棘在水分过于充足的环境下,能够通过提高海绵组织比例加速叶片蒸腾作用,散失多余水分。CTR和SR值体现了叶片厚度、栅栏组织厚度与海绵组织厚度之间存在一定的相互制约关系,使其比值能够保持相对稳定[26]。干旱条件下沙棘幼叶的CTR显著高于CK,SR显著低于CK;老叶相反。水涝条件下沙棘幼叶和老叶的CTR均为3种处理中最低,SR最高。水涝环境下叶片结构疏松,利于气体在叶肉细胞之间自由流动;干旱下幼叶结构紧密,有助于植物保存叶片内部水分;老叶结构疏松,植物抗旱能力减弱。
发达的维管系统可提高植物水分和养分的运输效率,减缓干旱对叶片造成的萎蔫损伤[27-28]。于海燕等[20]研究发现,文冠果在干旱环境中叶脉维管系统明显更加发达,导管明显增粗。本试验结果中,沙棘幼叶仅维管束直径表现出水涝条件下显著小于CK,主叶脉直径和木质部最大导管直径在3种水分条件下无显著差异;老叶在干旱和水涝条件下的主叶脉直径、维管束直径和木质部最大导管直径均显著大于CK。这可能是幼叶距茎基部较远,故叶脉和维管组织对不同水分条件反应不敏感;而老叶相对靠近茎基部,水分运输路径较短,所以可积极发育,以加速水分由根系向叶片输送,适应干旱和水涝环境,干旱环境下有助于叶片抵御缺水萎蔫,水涝环境下与疏松程度较高的叶肉组织配合加速蒸腾。另外,沙棘幼叶相较于老叶,感受强光辐射及高温更为敏感,叶片蒸腾量大于老叶,故叶片厚度及叶肉组织相关指标对干旱和水涝的反应强于老叶。
综上所述,对辐射及高温较敏感的沙棘幼叶通过叶片、表皮和栅栏组织显著增厚,栅海比和CTR显著升高,SR显著降低等表皮和叶肉结构变化适应干旱环境;通过叶片、栅栏组织显著变薄,栅海比和CTR显著降低,SR显著升高等叶肉结构变化适应水涝。靠近顶端距茎基部较近的沙棘老叶则主要通过叶脉维管组织显著增粗适应不同水分条件。
猜你喜欢
动漫界·幼教365(大班)(2021年11期)2021-11-20
金山(2020年9期)2020-11-10
发明与创新(2020年26期)2020-06-27
小学生学习指导(低年级)(2019年9期)2019-09-25
数学小灵通(1-2年级)(2017年9期)2017-10-13
东坡赤壁诗词(2017年3期)2017-07-05
红领巾·萌芽(2017年2期)2017-03-09
语文世界(初中版)(2017年1期)2017-03-06
小说月刊(2015年2期)2015-04-23
大江南北(2014年3期)2014-11-23
