
黄檀扦插生根过程中叶片营养物质含量变化研究
2023-06-30 07:13黄光卫肖京武胡育文陈乔松谢昱昀
湖南林业科技 2023年3期
黄光卫,肖京武,张 斌,陆 佳,胡育文,陈乔松 ,谢昱昀
(1.永州市林业科学研究所,湖南 永州 425045; 2.中南林业科技大学林学院,湖南 长沙 410004; 3.中南林业科技大学水土保持与荒漠化防治湖南省高等学校重点实验室,湖南 长沙 410004)
黄檀(Dalbergiahupeana)是中国特有的植物,因展叶较晚,又名“不知春”。这种豆科(Fabaceae)黄檀属(Dalbergia)的落叶乔木为深根性树种,广泛分布于亚热带和亚热带—暖温带过渡区域,属于泛热带分布区类型[1]。黄檀的适应能力强,可以耐受干旱和贫瘠的土壤。在《湖南省主要栽培珍贵树种参考名录(2020年版)》中,黄檀位列第11位,是湖南省重点推荐栽培利用的乡土珍贵优质硬木树种。
天然林中黄檀的种子繁殖力较弱,树体生长速度缓慢。受到人为砍伐破坏和逐年恶劣的自然环境影响,野生黄檀资源持续减少[2-3]。因此,开展黄檀扦插繁育技术研究,对保护和利用黄檀资源,不断扩大栽培面积,增加资源总量,充分挖掘其经济、生态和社会效益具有十分重要的理论和现实意义。
目前对交趾黄檀(Dalbergiacochinchinensis)、印度黄檀(Dalbergiasissoo)、降香黄檀(Dalbergiaodorifera)等的研究较多,主要集中在抗逆性、栽培、病虫害防治、化学成分等方面,如钟栎等[4]发现,采用IAA、NAA和抗坏血酸速蘸15~20 s的处理组合,降香黄檀扦插成活率最高达到85.2%。还有一些学者对影响降香黄檀扦插生根的生长调节剂[5]、插条粗度和萌芽度[6]等进行了初步研究,发现这些因素均显著影响降香黄檀的扦插成活率。
植物在扦插过程中需要大量的营养物质,插穗内的营养物质是插穗生根的重要条件。插穗内所存储的可溶性糖作为渗透调节物质和抗氧化剂可起到保护插穗的作用。淀粉可水解为可溶性糖,在一定程度上补充消耗掉的可溶性糖。可溶性蛋白是植物生根的营养物质,同时还能调节代谢、增强插穗的抗逆性。磷是细胞的组成成分,在蛋白质、脂肪和糖代谢中发挥着重要作用。缺磷时,细胞分裂受到影响,蛋白质合成受阻,植物生长缓慢。有关研究表明,插穗生根所需要的营养物质主要是碳水化合物和含氮化合物,可溶性糖含量高、氮素达到一定水平时,扦插生根成活率高[7-8]。
目前对黄檀扦插过程中营养物质含量的研究尚未见报道。本文研究黄檀扦插生根过程中叶片内可溶性糖、淀粉、可溶性蛋白质、全氮、全磷等营养物质的动态变化规律,旨在为黄檀的扦插繁殖提供理论依据。
1 试验地概况
本试验在湖南省岳阳市平江县加义镇境内的芦头实验林场内进行。芦头实验林场地理坐标为113°51′—113°58′E、28°31′—28°38′N。实验林场地处中亚热带向北亚热带过渡的气候带,属湿润的大陆性季风气候,四季分明,日照充足,降水充沛,年平均气温为15.9 ℃,年平均日照1 930 h,平均空气湿度为82%,年平均降水量为1 450.8 mm,雨季为3—7月,雨季降水量占全年降水量的66.8%。土壤以红壤、山地黄壤、山地黄棕壤为主,成土母岩多样化。
2 材料与方法
2.1 试验材料
2021年3月21日,从林场内3年生黄檀实生苗上选择生长健壮、无病虫害、带有腋芽的1年生枝条,剪取插穗13~15 cm,每插穗保留2~3个腋芽,剪口靠近腋芽下方、呈45度斜切面。
2.2 试验方法
扦插容器选用32孔黑色穴盘,扦插基质为体积比为1∶1的珍珠岩和泥炭土混合基质。扦插前3 d,基质装盘后用0.1%KMnO4溶液喷淋消毒并覆盖塑料薄膜,扦插前1 d揭除薄膜。扦插当天剪好插穗,基部对齐捆好,然后将插穗基部2~3 cm蘸满泥浆。用小木棍在基质中打孔,将黄檀插穗插入孔中并压实周围基质,共扦插150株。搭建宽约1.5 m、高约0.7 m的拱架,架上覆盖2.5 m宽的塑料薄膜,扦插好的穴盘放置拱棚内,薄膜四周用土压实。扦插后根据基质干湿情况,每隔3~4 d浇水1次。插穗出芽后光照强时加盖遮阳率为85%的遮阳网;气温达到32 ℃以上时,将拱棚两端的薄膜揭开通风降温。当芽生长到3~5 cm时,揭掉薄膜,只盖遮阳网。每隔7~10 d交替用0.1%多菌灵溶液和0.3%KMnO4溶液进行消毒。
2.3 指标测定方法
扦插后定期观察,扦插60 d后为大量生根期,此时母树叶片处于展叶期;70 d后为根系伸长期,此时母树叶片处于全叶期。分别于大量生根期、根系伸长期随机取扦插插穗15棵,随机采取叶片共10 g,设三次重复,同时取母树叶片作为对照,用液氮处理后放入-80 ℃的超低温冰箱中备用。可溶性糖、淀粉含量的测定采用蒽酮比色法[9],可溶性蛋白含量的测定采用考马斯亮兰G-250染色法[10],全氮、磷含量的测定采用硫酸-高氯酸消煮法[11],各项指标的测定均三次重复。
2.4 数据统计与分析
使用SPSS 25.0和Excel 2010对试验数据进行统计分析。用Origin 2018软件绘图。
3 结果与分析
3.1 黄檀插穗叶片可溶性糖含量的变化
可溶性糖是植物体生命活动重要的能量来源,为插穗不定根的生长提供营养,有利于插穗的根系生长。如图1所示,大量生根期和根系伸长期插穗叶片中的可溶性糖含量分别为106.44 mg·g-1和80.01 mg·g-1,大量生根期的明显高于根系伸长期的;而母树叶片展叶期的可溶性糖含量低于全叶期的。黄檀扦插从大量生根期到根系伸长期,插穗呼吸作用逐渐增强,大量不定根的伸长需要消耗能量,导致可溶性糖含量降低,这与陈洁等[12]的研究一致。从展叶到全叶期,黄檀母树叶片可溶性糖含量上升。

图1 插穗及母树叶片内可溶性糖含量Fig.1 Soluble sugar content in the leaves of cuttings and parent tree
3.2 黄檀插穗叶片淀粉含量的变化
如图2所示,在大量生根期和根系伸长期,插穗叶片中的淀粉含量均高于母树叶片中的淀粉含量。插穗的淀粉含量在大量生根期和根系伸长期分别为31.17 mg·g-1和44.35 mg·g-1,大量生根期的淀粉含量低于根系伸长期的,可能是因为在根系伸长期,插穗新梢逐渐舒展,光合作用增强,使得淀粉的合成速率加快;同时,不定根出现后插穗吸收能力增强,能够吸收水分和营养,所以淀粉含量出现了小幅度的增加。

图2 插穗及母树叶片内淀粉含量Fig.2 Starch content in the leaves of cuttings and parent tree
3.3 黄檀插穗叶片可溶性蛋白含量的变化
如图3所示,在大量生根期和根系伸长期,插穗叶片中的可溶性蛋白含量均高于母树叶片中的可溶性蛋白含量。插穗在大量生根期和根系伸长期的可溶性蛋白含量分别为12.37 mg·g-1和29.22 mg·g-1,根系伸长期的可溶性蛋白含量是大量生根期的2倍,可能是因为大量生根后,插穗开始抽梢,插穗光合能力逐渐恢复,促进了可溶性蛋白的积累,植株对蛋白质的消耗减少,蛋白质的含量上升。

图3 插穗及母树叶片内可溶性蛋白含量Fig.3 Soluble protein content in the leaves of cuttings and parent tree
3.4 黄檀插穗叶片全氮含量的变化
如图4所示,插穗大量生根期和根系伸长期全氮含量分别为35.31 mg·g-1和20.15 mg·g-1。在大量生根期,插穗叶片中的全氮含量低于母树叶片中的含量,说明此阶段插穗对氮素的需求不高,低浓度的氮素含量有利于插穗不定根的形成。在根系伸长期,插穗叶片中的全氮含量稍高于母树叶片中的全氮含量。而根系伸长期插穗叶片的全氮含量低于大量生根期的,说明大量生根后,植株对氮素需求增加,总氮快速消耗有利于不定根的伸长。
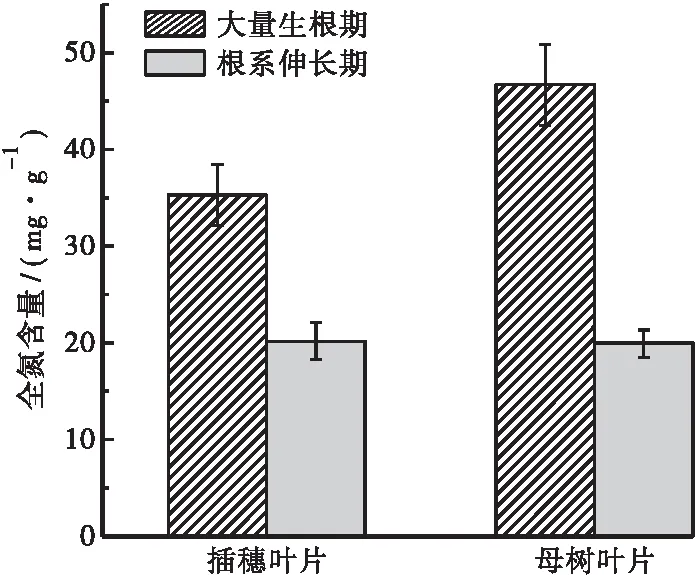
图4 插穗及母树叶片内全氮含量Fig.4 Total nitrogen content in the leaves of cuttings and parent tree
3.5 黄檀插穗叶片全磷含量的变化
磷是细胞的组成成分,在蛋白质代谢、脂肪代谢和糖类代谢中发挥着重要的作用。缺磷时,蛋白质合成受阻,新的细胞质和细胞核形成减少,细胞分裂受到影响,生长缓慢。黄檀插穗叶片全磷含量的变化如图5所示。在大量生根期和根系伸长期,插穗叶片全磷含量均高于母树叶片的全磷含量。插穗在大量生根期和根系伸长期的全磷含量分别为6.67 mg·g-1和4.34 mg·g-1,根系伸长期的全磷含量低于大量生根期的,说明大量生根后,植株对磷素需求增加,全磷含量下降。

图5 插穗及母树叶片内全磷含量的变化Fig.5 Total phosphorus content in the leaves of cuttings and parent tree
4 讨论与结论
氮是植物体内维生素、氨基酸和能量系统的组成部分,而且能增加植物中蛋白质的含量[7]。磷是细胞的组成成分,在糖类代谢、蛋白质代谢和脂肪代谢中起着重要的作用;缺少磷时,蛋白质合成受阻,新的细胞质和细胞核形成减少,从而影响细胞分裂,导致植物生长缓慢;磷元素可被重复利用[8]。蛋白质在生物体内参与构成细胞、调节代谢等多种功能[13]。可溶性糖及淀粉同样也对植物的生长发育及扦插生根起调控作用[14-15]。在插穗不定根的形成过程中,许多活动需要消耗营养物质,例如愈伤组织的形成、大量根的产生和根系的伸长[16-17]。从大量生根期到根系伸长期,需要消耗大量的营养物质,而插穗在生根前无法从土壤中吸收营养物质,因此掌握这个阶段插穗中的营养物质变化,能为促进插穗的生根及其根系的伸长提供理论依据。
两个时期插穗叶片中的可溶性蛋白、淀粉、可溶性糖、全磷的含量均高于母树叶片中的,而大量生根期插穗叶片中的全氮含量明显低于母树叶片中的。其原因有可能是愈伤组织、不定根出现后插穗具有了吸收外界营养的能力,插穗产生大量不定根,进而形成一个完整独立的植株,可以进行一定程度的光合作用,可溶性糖等的含量较母树叶片出现了不同程度的增加。然而,插穗叶片中的全氮含量仅在大量生根期低于母树叶片中的,在根系伸长期与母树叶片中的相差不大,这表明在生根过程中稀释了氮素的浓度,大量生根期氮素含量的降低有利于黄檀插穗不定根的形成,高含量的氮素不利于黄檀生根;这与詹亚光等[18]研究白桦(Betulaplatyphylla)、葛文志等[19]研究胡桃楸(Juglansmandshurica)时发现的营养物质含量变化规律比较类似。
从大量生根期到根系伸长期,插穗叶片中可溶性糖、全氮、全磷的含量逐渐降低,而可溶性蛋白、总淀粉的含量逐渐升高,可能的原因是在扦插时插穗需要消耗大量能量来克服逆境环境进行生长和皮孔萌动,这些能量是从扦插枝条本身储存的营养物质中获取的[20-21]。从大量生根期到根系伸长期,插穗叶片逐渐舒展并且光合作用效率提高,使得可溶性蛋白和淀粉的合成速率加快;此时,根系开始形成,插穗具有了吸收外界营养的能力,能够吸收水分和营养,因此,可溶性蛋白和淀粉的含量逐渐在插穗基部和叶片上积累,并呈现一种逐渐上升的趋势[22]。本试验对可溶性蛋白含量、淀粉含量的研究结果与杜伟等[23]对桑(Morusalba)、王小玲等[24]对四倍体刺槐(Robiniapseudoacacia)的研究结果相似。可溶性糖、全氮、全磷在大量生根期的含量都高于在根系伸长期的,说明根系的生长需要消耗大量的可溶性糖、氮、磷,这与陈洁等[12]、曾炳山等[25]的研究一致。
综上所述,在大量生根期,氮素含量的降低有利于黄檀插穗不定根的形成,高含量的氮素不利于黄檀生根。黄檀插穗从大量生根期到根系伸长期,可溶性蛋白、总淀粉的含量随着扦插时间延长逐渐升高,说明插穗枝条叶片逐渐舒展后,光合作用增强,可以吸收基质中的营养物质,故可溶性蛋白、淀粉的含量逐渐在基部及叶片上积累,会呈现一个上升的趋势。而其全氮、全磷、可溶性糖的含量从大量生根期到根系伸长期逐渐降低,说明黄檀插穗根系的生长需要消耗大量的可溶性糖、氮、磷。因此,黄檀的扦插生根过程与插穗自身的营养水平密切相关且营养物质对黄檀的扦插生根和根系伸长起着至关重要的调节作用。
黄檀插穗在大量生根期,仅全氮的含量低于同期母树叶片的,其他营养物质的含量均高于同期母树叶片的,说明低浓度的氮素含量有利于其插穗不定根的形成。可溶性糖是植物体生命活动最直接的能量来源,在根系伸长期,黄檀的主要能量来源为可溶性糖。插穗从大量生根期到根系伸长期,可溶性糖含量显著下降,全氮、全磷的含量也有所降低,说明根系的伸长对可溶性糖、氮和磷的需求较大,可通过外部施用糖溶液、氮素和磷素来促进根系的生长。
猜你喜欢
林业与生态(2022年5期)2022-05-23
山东林业科技(2021年4期)2021-08-31
福建林业科技(2018年1期)2018-04-09
山东林业科技(2017年1期)2017-06-29
现代农业(2016年3期)2016-04-14
广东药科大学学报(2016年1期)2016-04-11
广西林业科学(2016年2期)2016-03-20
土壤与作物(2015年3期)2015-12-08
郑州大学学报(理学版)(2012年4期)2012-03-25
浙江林业科技(2011年2期)2011-05-09
