
蓝莓叶斑病病原菌鉴定及生物学特性
2023-06-30 12:50:23罗其鑫杨莹李思尚晓静侯瑞
江苏农业科学 2023年5期
罗其鑫 杨莹 李思 尚晓静 侯瑞


摘要:为明确贵州省黔东南苗族侗族自治州麻江县蓝莓叶斑病病原菌种类及其生物学特性,采用组织分离法获得了代表性病原菌株Y2,科赫氏法则证明Y2是引起蓝莓叶斑病的病原菌。形态学和分子生物学综合鉴定菌株Y2为链格孢菌(Alternaria alternate)。生物学特性研究结果表明,病原菌Y2在PDA培养条件下,气生菌丝较发达,菌丝呈白色、密集,基质背面呈淡黄色,分生孢子单生,褐色,卵圆形、倒棒形,有纵横隔膜。该菌菌丝生长的最佳培养基为察氏培养基,产孢最适培养基为WA培养基;菌丝生长的最适温度为25 ℃,产孢最适温度为5 ℃;菌丝对碳源葡萄糖利用效果最好,但麦芽糖能更好地促进Y2产孢;菌丝生长最适氮源为蛋白胨,产孢最适氮源为硫酸铵;全光照适宜菌丝生长,但全黑暗促进Y2产孢;菌丝生长和产孢最适pH值为7。通过Y2菌株生物学特性研究,能够为蓝莓叶斑病防治提供理论依据。
关键词:蓝莓;叶斑病;链格孢菌;生物学特性
中图分类号:S436.639文献标志码:A
文章编号:1002-1302(2023)05-0146-09
蓝莓学名笃斯越橘(Vaccinium uliginosum L.),屬杜鹃花科(Ericaceae)越橘亚科(Vaccination)越橘属(Vaccinium)植物。蓝莓浆果中含有丰富的花青素、鞣花酸等活性物质,可以抗衰老、减少癌症等的发病率[1-3]。随着我国蓝莓种植规模的逐渐扩大,日趋突出的蓝莓病害导致蓝莓产业发展受阻[4]。目前已报道的蓝莓常见病害主要有蓝莓叶斑病[5]、枝枯病[6]、根癌病[7]、灰霉病[8]、间座壳芽枯病[9]和枝干溃疡病[10]等。蓝莓叶斑病已成为蓝莓常见病害,西澳大利亚州报道了在蓝莓上由极细链格孢菌(Alternaria tenuissima)引起的叶斑病[11],Norman等报道了由青枯菌(Ralstonia solanacearum)引起的蓝莓青枯病[12],国内研究报道的有由根癌土壤杆菌(Agrobacterium tumefaciens)引起的蓝莓根癌病[7]、灰葡萄孢菌(Botrytis cinerea)引起的蓝莓灰霉病[13]和极细链格孢引起的蓝莓叶斑病[4]。蓝莓叶斑病致病菌复杂多样,严雪瑞等研究发现引起蓝莓叶穿孔病的致病菌为链格孢菌(Alternaria alternate)[14],张国辉等在贵州省黔东南州台江县发现了巨腔茎点霉(Phoma macrostoma)为病原的蓝莓叶斑病[15]。明确致病菌种类及其生物学特性有助于该病害后续的综合防治。
由于蓝莓叶斑病病原菌的多样性,导致不同地域叶斑病致病菌的生物学特性不同,使蓝莓叶斑病的防治较为困难。本试验对引起贵州省黔东南苗族侗族自治州麻江县蓝莓叶斑病的病原菌进行鉴定和生物学特性研究,明确其菌丝生长的最适条件和产孢情况,研究结果可为防治蓝莓叶斑病提供理论依据。
1 材料与方法
1.1 试验材料
供试发病蓝莓叶片在贵州省黔东南苗族侗族自治州麻江县蓝莓栽培生产基地采集,塑封袋封装带回实验室后于4 ℃冰箱保存备用。
1.2 试验方法
1.2.1 蓝莓叶斑病病原菌的分离及致病性测定
将分离纯化后的菌株Y2接种至健康叶片上,致使健康蓝莓叶片感病,结果发病蓝莓的叶片上应能得到与初始接种的菌株性状一致的菌落,对照组不会得到与初始菌株一致的菌落。接种前先用75%乙醇对叶片表面进行消毒,接着用无菌水反复冲洗3次,移入5%(体积分数) NaClO溶液中灭菌2 min,用无菌水清洗后晾干备用。
离体接种:用消过毒的接种针在消过毒的健康蓝莓叶片表面避开叶脉刺4个小孔,于小孔上贴直径5 mm的菌饼,覆上灭菌的湿脱脂棉,再套保鲜袋保湿,试验每张蓝莓叶片接种2个菌饼,每个菌株处理5个重复,1个对照组。
1.2.2 蓝莓叶斑病病原菌的鉴定
形态学鉴定:超净操作台中,用内径为5 mm的打孔器在2种病原菌Y2的菌落边缘打出菌饼,将Y2菌株的菌饼接种在PDA培养基上并置于28 ℃培养箱中2、4、6 d进行培养皿观察拍照,观察其菌落特点(菌丝形态、颜色、基质颜色、菌丝生长速度、菌丝密度等)。在光学显微镜下参照《真菌鉴定手册》对菌丝的分生孢子梗及分生孢子进行形态学观察,记录其形态特征并测量分生孢子大小。
分子生物学鉴定:病原菌Y2用PDA平板进行扩繁后,刮取PDA平板上活化好的Y2菌株菌丝,用Fungal DNA Midi Kit真菌DNA提取试剂盒(OMEGA)提取病原菌Y2的DNA作为模板,采用真菌通用引物ITS1(5′-TCCGTAGGTGAACCTGCGG-3′) 和ITS4(5′-TCCTCCGCTTATTGATATGC-3′)对提取的DNA进行PCR扩增,扩增产物经1.2%琼脂糖凝胶电泳检测条带后,切胶回收PCR目的条带送至重庆擎科兴业生物技术有限公司进行测序。将病原菌Y2的18S RNA序列与NCBI数据库相近序列进行比对。利用MEGA7.0的Clustal W进行多序列比对,再采用邻接法(neighbor joining,NJ)进行菌株系统发育树的构建。
1.2.3 蓝莓叶斑病病原菌生物学特性研究
无菌环境下,截取直径5 mm的病原菌新鲜菌饼,置于7个不同的碳源(葡萄糖、D-果糖、乳糖、D-麦芽糖、可溶性淀粉、甘露醇、蔗糖)培养基,不同的氮源(硫酸铵、尿素、甘氨酸、蛋白胨、酵母粉、牛肉膏、硝酸钠)培养基,6个不同营养基质(PDA、燕麦、PSA、蓝莓煎叶、WA、C)中培养,第3天量取菌落直径,第5天时对培养皿拍照,30 d后测定产孢量;接种在pH值初始值分别为 5、7、9、11的PDA平板中央,第3天量取菌落直径,第5天时对培养皿拍照,30 d后测定产孢量;置于不同温度(5、15、25、28、35 ℃)下培养3 d,测量菌落直径,在第5天时对培养皿拍照,30 d 后测定产孢量;置于3个光照处理(全黑暗:24 h 黑暗;全光照:24 h光照;半光照:光照12 h/黑暗 12 h)下培养,第3天量取菌落直径,第5天时对培养皿拍照,30 d 后测定产孢量。以上所有处理均设置3个重复,菌落直径测量采用十字交叉法,产孢量用血球计数板测定。
1.3 数据分析
利用Excel 2010、SPSS 19进行数据处理和Photoshop作图。
2 结果与分析
2.1 蓝莓叶斑病病原菌的分离及鉴定
分离得到编号为Y2的病原菌。在28 ℃恒温环境下,病原菌Y2在PDA培养基上生长 6 d 时,病原菌的生长速度较慢,产孢,气生菌丝较发达,菌丝呈白色、密集,基质背面呈淡黄色(图1)。分生孢子单生,褐色,卵、圆形倒棒形,有纵横隔膜(图2)。
结合病原菌Y2的菌落形态和显微特征,初步将病原菌Y2判定为链格孢属(Alternaria)菌物。对病原菌Y2进行rDNA-ITS测序,将获得的序列在NCBI数据库进行BLAST同源性比对,比对结果表明,病原菌Y2序列与登录号为MN756011.1的Alternaria alternate同源性最高,根据MEGA7.0中邻接法构建病原菌Y2的系统发育树(图3)。系统发育树结果表明,病原菌Y2与链格孢菌(Alternaria alternate) MN756011.1菌株的亲缘关系最近,结合病原菌的形态特征和ITS序列特征,故最终将病原菌Y2鉴定为链格孢菌(Alternaria alternate)。
2.2 蓝莓叶斑病病原菌Y致病性的测定
将病原菌Y2的菌饼接种到针刺伤的蓝莓叶片伤口处,以无菌PDA培养基的菌饼作为对照,培养 3 d 时取下菌块,5 d后观察到:接种病原菌Y2的叶片在接种处有少量的气生菌丝,叶片伤口处呈褐色且呈较规则的圆形,菌丝呈白色(图4-A),与采集的田间病叶发病症状相似(图4-C),未接种的叶片未出现发病症状(图4-B),根据科赫氏法则,从发病的叶片病健交界部位组织分离得到一种分离物,和Y2为同一菌株。
[FK(W10][TPLQX44.tif][FK)]
2.3 不同碳源对菌丝生长的影响
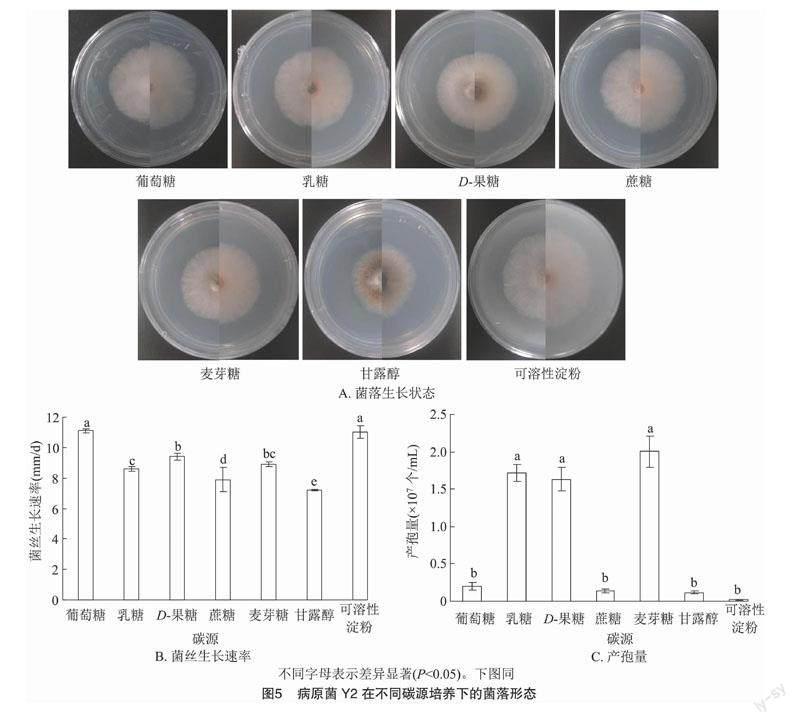
病原菌Y2在7种不同碳源培养基中的菌落形态都呈较规则的圆形,菌丝密集,有少量气生菌丝;在甘露醇作碳源时产生较多的黄色色素,其次,D-果糖、麦芽糖2种碳源培养基培养时有少量黄色色素,其余不同碳源培养基均有微量黄色色素产生(图5-A)。病原菌Y2在7种不同碳源培养基中菌丝生长速率存在显著差异(P<0.05,下同),在葡萄糖、可溶性淀粉作为碳源时的菌落状态最好,菌丝的繁殖速度最快,生长速率为11.1、11.0 mm/d;显著高于其他碳源的生长速率;生长速率最慢的是碳源为甘露醇,为7.2 mm/d(图5-B)。30 d后,测定其孢子浓度(图5-C)。病原菌Y2在麦芽糖、乳糖和D-果糖为碳源时产孢量显著高于其他碳源,麦芽糖作为碳源的培养皿中产孢子数量最多,为 2.003×107个/mL,以可溶性淀粉、葡萄糖、蔗糖和甘露醇为碳源时病原菌Y2的产孢量无显著差异,以可溶性淀粉为碳源时最少,为0.022×107个/mL,以葡萄糖、蔗糖和甘露醇为碳源时分别为0.202×107、0.138×107、0.119×107个/mL。
2.4 不同氮源对菌丝生长的影响
病原菌Y2在7种不同氮源培养基中菌落形态较规则,均为圆形,气生菌丝较密集,在以蛋白胨、甘氨酸、硫酸铵、尿素为培养条件下,菌落圆心部位菌絲高于周围菌丝,形成圆形凸起,其中氮源为蛋白胨时凸起较明显;菌落在以酵母粉、甘氨酸、尿素、牛肉膏为氮源培养时均产生黄色色素,其余条件下无色素产生,以酵母粉培养时产生较大量的黄色色素与褐色色素(图6-A)。不同氮源处理下菌丝生长速率存在显著差异,在以蛋白胨为氮源时菌丝生长速率显著高于其他氮源,其菌落状况最好,繁殖最快,生长速率为13.3 mm/d,生长速率最小的是以尿素为氮源时,其生长速率为7.7 mm/d,以硝酸钠、甘氨酸为氮源时菌丝生长速率无显著差异,均为 10.5 mm/d (图6-B)。不同氮源处理下病原菌Y2产孢量有显著差异(图6-C),在以硫酸铵作为氮源时的病原菌Y2所产孢子量显著高于其他氮源,为21.667×107个/mL,说明硫酸铵能诱导病原菌Y2萌发孢子,是病原菌Y2产孢的最佳的氮源。
2.5 不同pH值对菌丝生长的影响
病原菌Y2在pH 值为 5、7、9、11的环境下都能形成较规则的圆形、菌丝较为密集的菌落,但以在pH值=7环境中的菌落直径最大,生长状态最佳,当pH值从5到11变化时,菌丝逐渐密集,且都有黄色素产生(图7-A)。病原菌Y2在pH值=7时生长速率显著高于其他pH值梯度,为10.5 mm/d;生长速率最小的是在pH值=11时,为8.7 mm/d;在pH值=5、pH值=9时,其菌丝生长速率分别为9.9、9.8 mm/d,二者间无显著差异(图7-B),病原菌Y2在酸性条件下更适宜生长。过酸、过碱的环境均不利于病原菌Y2产孢, 而pH值=7时酸碱适中,可以促进Y2产孢,产孢量显著高于其他pH值,为2.573×107个/mL,pH值=11时产孢量最少,只有0.074×107个/mL(图7-C)。
2.6 不同温度对菌丝生长的影响
病原菌Y2在5个温度值下培养5 d后均有菌丝长出,且菌落均呈规则圆形,气生菌丝密集,均有黄色素产生(图8-A)。当温度降至 5 ℃,菌株几乎停止生长,在25 ℃下最适宜病原菌Y2生长,生长速率显著高于其他温度,为8.9 mm/d;在5 ℃下最不适宜生长,生长速率为2.0 mm/d;在15、28、35 ℃ 下菌落生长速率分别为5.4、7.4、3.4 mm/d (图8-B)。低温可以促进病原菌Y2产孢,5 ℃时产孢量显著高于其他温度,为4.067×107个/mL,在温度为15 ℃和35 ℃时产孢量无显著差异,产孢量很少或几乎不产孢,当温度为25 ℃和28 ℃时产孢量较多,分别为2.077×107个/mL和0.435×107个/mL(图8-C)。
2.7 不同光照对菌丝生长的影响
在全光照、全黑暗和光暗交替3种光照条件下,病原菌Y2都能正常生长,菌落均呈规则圆形,且均有黄色色素产生,全黑暗培养下产生较大量的黄色、褐色色素;菌落气生菌丝均致密(图9-A)。病原菌Y2在全光照条件下生长速率显著高于全黑暗和光暗交替条件下,为12.4 mm/d;生长速率最小是在光暗交替条件下,为10.2 mm/d;在全黑暗条件下菌丝生长速率为10.9 mm/d (图9-B)。计算不同光照处理下致病菌Y2所产孢子量,不同光照对病原菌Y2产孢有显著影响。全黑暗处理的病原菌Y2所产孢子数量是3个处理组中最高的,为1.62×107个/mL,光暗交替处理下的Y2孢子数目最少,为0.012×107个/mL(图9-C)。
2.8 不同培养基对菌丝生长的影响
病原菌Y2在6种不同培养基中,28 ℃下培养5 d 后,观察到病原菌Y2在6种不同培养基中均呈较规则圆形,气生菌丝在PDA、PSA、察氏(C)培养条件下致密,在WA、蓝莓煎汁培养基、燕麦琼脂培养基中菌丝较稀薄;菌落在PDA、PSA、WA和燕麦琼脂培养基中有黑色色素产生,在察氏培养基中有黄色色素产生,蓝莓煎汁琼脂培养基中无色素产生(图10-A)。病原菌Y2在察氏培养基中生长速率显著高于其他培养基,为17.0 mm/d;生长速率最小的是在WA培养条件下,生长速率为7.0 mm/d;在PDA、PSA、蓝莓煎汁琼脂和燕麦琼脂培养条件下菌丝生长速率分别为10.6、11.1、8.9、11.9 mm/d (图10-B)。统计病原菌Y2在不同培养基中所产孢子量,发现WA和燕麦培养基中的孢子数显著高于PDA、PSA、察氏和蓝莓煎汁培养基,其中WA中孢子数最多, 为4.495×107个/mL,在察氏中Y2孢子数目最少,仅仅为0.014×107个/mL,在燕麦中产孢量较多,为4.233×107个/mL,在其他基质下孢子数量差异不显著(图10-C)。
3 讨论与结论
本研究从贵州省人工栽培蓝莓园内发现典型蓝莓叶斑病植株,通过组织分离法进行病原分离,科赫法则证明其致病性,根据菌落形态特征及现代分子生物技术,最终鉴定蓝莓叶斑病病原菌Y2为链格孢菌,病原菌Y2侵染离体叶片形成的病斑为红褐色圆形病斑,与链格孢菌侵染蓝莓叶片形成的病斑形似;链格孢菌作为主要的蓝莓叶斑病病原菌,在蓝莓叶部病害中较为常见。链格孢菌还可引起多种植物病害,如烟草赤星病[16]、高粱叶斑病[17]、梨黑斑病[18]、花椒穗枯病[19]、番茄黑斑病[20]、枣果黑斑病[21]、荞麦叶枯病[22]和玉米叶斑病[23]等。
本研究发现蓝莓叶斑病病原菌-链格孢菌Y2,菌丝生长的最适pH值为7,与周银丽等[24]和卢文洁等[22]研究结果一致;黎研研等[25]、周银丽等[24]、卢文洁等[22]和刘晓琳等[21]的研究结果均表明菌丝生长的最适温度为25 ℃,本研究得到最适氮源为蛋白胨与黎研研等[25]、王宏等[18]和刘晓琳等[21]的研究结论相同,卢文洁等[22]的研究表明,全光照条件下菌丝生长最快,本研究也得到了相同的结论。关于链格孢菌的生物学特性有很多不同的报道。乔镜澄等研究了番茄黑斑病样品中链格孢菌的生物学特性,结果显示,该菌株的最适温度为28 ℃,最适pH值为6,最佳碳源为乳糖,最佳氮源为酵母膏,PDA为其最适宜生长的培養基,菌丝在完全黑暗条件下扩展最快[20];杨晓平等报道了湖北梨黑斑病病原菌链格孢菌菌丝生长的最佳条件是PSA培养基[26];陈凤美等报道了江苏银杏内生链格孢菌最适pH值为5~9,最适氮源为硝酸钠[27];郑肖兰等报道了海南省南繁区玉米链格孢叶斑病链格孢菌的最适温度为30 ℃,最适pH值为5,最适培养基是PDA+玉米绿叶煎汁,最适的氮源是缬氨酸和甘氨酸。
本试验研究了不同处理下病原菌Y2的产孢情况,结果表明,病原菌Y2产孢的最适碳源是麦芽糖、最适氮源是硫酸铵,最适温度为5 ℃、pH值为7,以WA为营养基质有利于病原菌Y2产孢,全黑暗促进Y2产孢。胡中会等报道了柑橘褐斑病病原菌为链格孢菌,其产孢的最适温度为21 ℃,最适碳源为甘露醇,最适氮源为甘氨酸[28];邹凤莲等报道了番红花链格孢菌的最适温度为28 ℃,最适碳源为蔗糖,最适氮源为牛肉浸膏[29];袁高庆等报道了毛叶枣黑斑病病原菌的最适温度为30 ℃,最适pH值为5,PDA培养基最适宜病菌产孢[30]。说明适宜链格孢菌产孢的条件随着地理环境、寄主的变化而产生较大差异。本研究对该菌菌丝生长和产孢的生物学特性的研究结果可为后续该菌的致病机制研究以及科学有效的防治提供重要参考。
本研究明确了贵州省蓝莓叶斑病的病原菌为链格孢菌,并分析了其生物学特性。中国蓝莓叶斑病的病原菌种类复杂多样,病害防控工作的基础原则是准确鉴别病原菌;同一病原菌的生物学特性在不同环境、不同寄主中存在差异,因此,研究病原菌Y2的生物学特性对进一步阐述叶斑病病害的发病规律和综合防治具有的重要指导意义。
参考文献:
[1]胡雅馨,李 京,惠伯棣. 蓝莓果实中主要营养及花青素成分的研究[J]. 食品科学,2006,27(10):600-603.
[2]陈介甫,李亚东,徐 哲. 蓝莓的主要化学成分及生物活性[J]. 药学学报,2010,45(4):422-429.
[3]李亚东,张志东,吴 林. 蓝莓果实的成分及保健机能[J]. 中国食物与营养,2002,8(1):27-28.
[4]余 磊,赵建荣,徐胜光,等. 云南蓝莓叶斑病的病原菌鉴定[J]. 植物病理学报,2015,45(2):216-219.
[5]冯 璐,栾雨时,范永强,等. 越橘叶斑病病原菌的鉴定[J]. 东北农业大学学报,2007,38(5):614-618.
[6]余 磊,赵建荣,Impaprasert R,等. 蓝莓枝枯病病原菌鉴定[J]. 植物病理学报,2013,43(4):421-425.
[7]傅俊范,彭 超,严雪瑞,等. 蓝莓根癌病发生调查及病原鉴定[J]. 吉林农业大学学报,2011,33(3):283-286,292.
[8]严雪瑞,赵睿杰,周 源,等. 蓝莓灰霉病菌差异性比较及蓝莓品种抗病性鉴定[J]. 果树学报,2014,31(5):912-916.
[9]严雪瑞,王 旭,胡梦琼,等. 蓝莓间座壳芽枯病病原菌鉴定及其生物学特性[J]. 植物病理学报,2015,45(5):556-560.
[10]徐成楠,周宗山,张红军,等. 蓝莓枝干溃疡病病原鉴定[J]. 植物病理学报,2012,42(5):532-535.
[11]You M P,Lanoiselet V,Wang C P,et al. First report of Alternaria leaf spot caused by Alternaria tenuissima on blueberry (Vaccinium corymbosum) in western Australia[J]. Plant Disease,2014,98(3):423.
[12]Norman D J,Bocsanczy A M,Harmon P,et al. First report of bacterial wilt disease caused by Ralstonia solanacearum on blueberries (Vaccinium corymbosum) in Florida[J]. Plant Disease,2018,102(2):438.
[13]董克锋,姜惠铁,高海霞. 冬暖棚有机蓝莓园蓝莓灰霉病发生及预防措施[J]. 中国果树,2012(5):67-69,79.
[14]严雪瑞,周 源,赵睿杰,等. 蓝莓叶穿孔病菌鉴定及其生物学特性研究[J]. 北方园艺,2014(16):123-127.
[15]张国辉,刘德波,宋盛英,等. 黔东南州蓝莓叶部病害的种类调查和病原鉴定[J]. 中国森林病虫,2017,36(2):42-46.
[16]彭希文,劉光珍,杨永柱,等. 云南省烟草赤星病(Tobacco brown spot)病原研究及其防治药剂的筛选[J]. 西南农业大学学报,2000,22(2):153-156.
[17]赵艳琴,于华荣,石 凯,等. 高粱链格孢叶斑病病原鉴定[J]. 植物病理学报,2017,47(2):282-285.
[18]王 宏,常有宏,陈志谊. 梨黑斑病病原菌生物学特性研究[J]. 果树学报,2006,23(2):247-251.
[19]刘 峰,杨文香,张 娜,等. 花椒枯穗病病原鉴定[J]. 植物病理学报,2013,43(3):310-313.
[20]乔镜澄,刘 宇,马敬昊,等. 番茄黑斑病病原菌的鉴定及生物学特性研究[J]. 江苏农业科学,2017,45(10):94-97.
[21]刘晓琳,刘 玉,马 荣,等. 新疆枣果黑斑病病原菌鉴定及生物学特性[J]. 西北林学院学报,2015,30(3):132-138.
[22]卢文洁,罗志明,李春花,等. 云南省荞麦叶枯病病原菌鉴定及其生物学特性[J]. 植物保护学报,2019,46(5):1065-1072.
[23]郑肖兰,赵 爽,韩小雯,等. 海南省南繁区玉米链格孢叶斑病病原菌鉴定及其生物学特性[J]. 江苏农业科学,2018,46(6):82-87.
[24]周银丽,白建波,胡先奇,等. 芒果叶斑病病原菌的鉴定及其生物学特性[J]. 江苏农业科学,2013,41(5):108-109.
[25]黎妍妍,杨 涛,贾欣欣,等. 湖北省烟草赤星病菌生物学特性研究[J]. 湖北农业科学,2018,57(22):27-31.
[26]杨晓平,胡红菊,王友平,等. 梨黑斑病病原菌的生物学特性及其致病性观察[J]. 华中农业大学学报,2009,28(6):680-684.
[27]陈凤美,刘 群,蒋继宏,等. 银杏内生链格孢菌GI009生物学特性研究[J]. 西北林学院学报,2005,20(4):112-114,121.
[28]胡中会,赵立华,张 铁. 柑橘褐斑病菌菌丝生长和孢子萌发研究[J]. 云南农业大学学报(自然科学),2015,30(1):9-12.
[29]邹凤莲,汪志平,卢 钢. 番红花链格孢菌的分离及其生物学特性研究[J]. 浙江大学学报(农业与生命科学版),2006,32(2):162-167.
[30]袁高庆,赖传雅,黄丽华. 毛叶枣黑斑病的病原鉴定及其生物学特性研究[J]. 中国农学通报,2003,19(1):44-47.
收稿日期:2022-04-12
基金项目:贵州省科技计划项目(编号:黔科合支撑[2017]2567、黔科合平台人才[2018]5781);贵州大学SRT计划[编号:贵大SRT(2019)252号]。
作者简介:罗其鑫(1999—),女,贵州贵阳人,硕士研究生,研究方向为森林病理学。E-mail:452450608@qq.com。
通信作者:侯 瑞,博士,副教授,主要从事森林病理学等研究工作。E-mail:jiayouhourui123@163.com。
猜你喜欢
中国农业科学(2022年16期)2022-09-19 03:36:38
河北果树(2021年4期)2021-12-02 01:15:08
甘肃教育(2021年10期)2021-11-02 06:14:26
音乐教育与创作(2020年2期)2020-06-19 06:45:46
商周刊(2017年5期)2017-08-22 03:35:22
作文周刊·小学一年级版(2017年26期)2017-08-10 20:31:15
现代园艺(2016年7期)2017-01-09 22:53:28
现代农业科技(2016年19期)2016-12-12 16:15:50
农业与技术(2016年19期)2016-12-12 03:12:12
农业与技术(2016年20期)2016-12-08 19:26:26
