
广西香椿半同胞家系子代测定及早期选择
2023-06-29 01:17舒应东蒙兰杨吴远媚潘涛罗筱娥潘永光
热带农业科学 2023年4期
舒应东 蒙兰杨 吴远媚 潘涛 罗筱娥 潘永光
(南宁市林业科学研究所 广西南宁 530107)
香椿(Toona sinensis)为楝科(Meliaceae)香椿属落叶乔木,为我国特有珍贵速生用材树种,具有树干通直、生长迅速、木材花纹美丽和芳香耐腐等特点,有“中国桃花心木”之称,多用于家具、装修和车船制造等领域[1-3]。目前有关香椿优良家系或无性系的选择研究主要在四川[4-7]、福建[8-10]和河南[11-12]等地,广西还未见报道。广西各地均有香椿分布,但缺乏良种,材用良种选育工作还处于起步阶段[13-16]。为此,南宁市林业科学研究所自2018 年以来在广西南宁市、河池市和百色市等地开展香椿种质资源调查及材用优树选择工作,收集了马山县、隆安县和德保县等种源的优树种子。本研究以广西10 个种源的33 个香椿优树的半同胞子代为研究材料,营建子代测定林,造林后连续2 年测定树高、胸径并计算材积,用来进行生长性状分析、遗传参数估算、变异系数和遗传增益计算,以期筛选出早期优良家系,为广西香椿的良种选育奠定基础。
1 材料与方法
1.1 材料
1.1.1 试材于2018 年10 月至12 月采用3 株优势木对比法在广西区内开展香椿材用选优工作,选择马山县、隆安县和德保县等10 个种源的33株优树(表1),采集优树种子繁育家系苗木营建的子代测定林。2019 年1 月将33 个家系的香椿种子放在沙床上催芽,2019 年2 月将已长出2片真叶的幼苗转移到育苗杯(规格为宽13 cm,高15 cm),育苗基质为97%泥土+3%过磷酸钙[有效磷(P2O5)≥12%],2019 年10 月营建子代测定林。
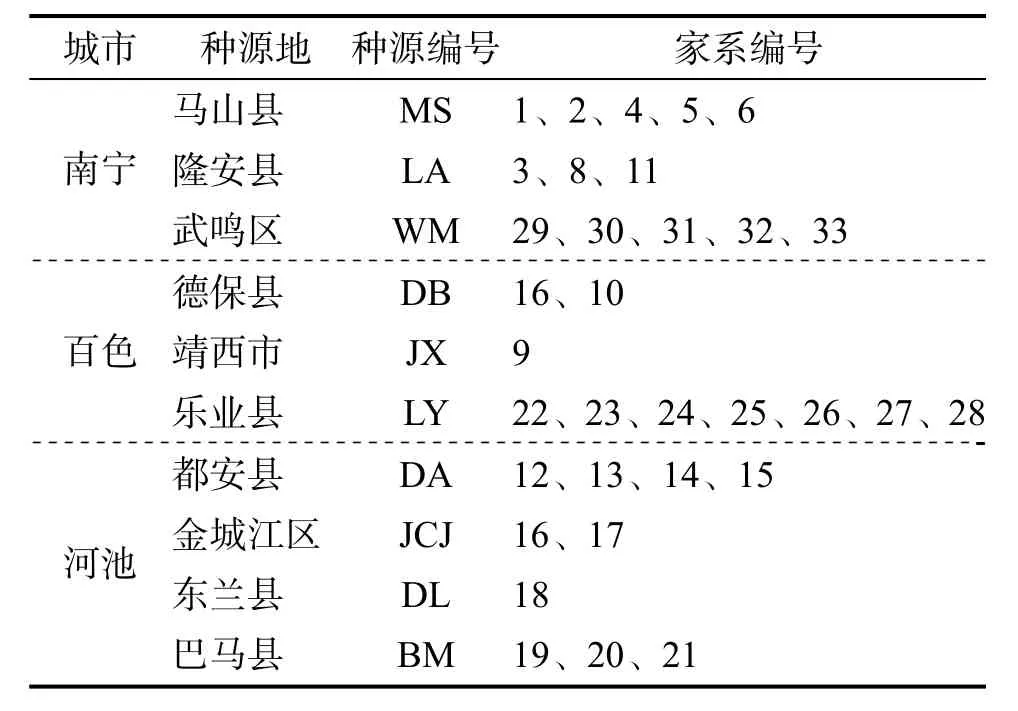
表1 香椿种源和家系
1.1.2 试验地概况试验地位于南宁市林业科学研究所(E107°59′,N23°10′),属石灰岩峰林间缓丘宽谷台地地貌,海拔100~159 m,属低山和丘陵地带,坡度在5~15°,地势平坦。气候为北热带北缘季风气候,四季温和,冬暖夏凉,年平均气温21.5℃,≥10℃的年平均积温7 697.8℃。1月份最冷,月平均温度为12.3℃,极端最低气温为-3℃;7 月份最热,平均气温28.2℃,极端最高温度40.6℃。年平均降雨量1 250 mm,雨季多在4—8 月,年均蒸发量1 613.8 mm,年平均相对湿度79.0%,有霜日年平均3~5 d。
土壤为第四纪红土发育成的中至厚层赤红壤,有机质0.499%~3.146%,pH 4.5~5.5,速效氮7.5~70 mg/kg,速效钾40~55 mg/kg,全氮0.036%~0.147%,土质粘性小,土壤保水性差、渗透强,比较干旱[17]。
1.2 方法
1.2.1 试验林营建子代测定林位于南宁市林业科学研究所第2 林班,地势平坦,面积2.3 hm²。采用随机区组试验设计造林,每个重复设置33 个种植小区,每个种植小区安排一个家系单行种植20 株,设置4 个重复。种植株行距3 m×2 m,试验林每重复间设1 行保护行。
造林地前期为柑橘林,于2019 年6 月份完成林地清理工作(包括对林地杂灌进行砍除,炼山清理,中耕机一犁一耙)。按长宽深为40 cm×40 cm×40 cm 的规格挖坎,每坎施放0.5 kg 过磷酸钙后拌土回坎。
2019 年10 月挑选33 个香椿家系生长良好、无病虫害和长势均匀的子代苗80 株,用于营建子代测定林。苗木造林前在容器袋中下部沿3 个不同方向纵切约5 cm 的切口。造林时首先把坎内的回填土挖出约1/2,再把容器苗垂直放入种植坎内,容器口与种植坎口持平或略低;然后回土填入种植坎,沿容器外围踩紧填土,最后覆盖一层松土(略高于苗木容器1~2 cm),做成中间低四周高的定植树盆,以便浇水、撒施肥料[18-19];植苗后立即浇足定根水。
造林后根据杂草的生长情况,每年对全林地割除杂草3 次,每年5 月开沟施复合肥(N∶P∶K=15∶15∶15)1 次,施0.5 kg/株。定期或不定期观察植株生长情况,防病虫害及人畜破坏。
1.2.2 数据收集于2020、2021 年12 月分别对子代测定林的每株家系苗木进行一次生长量测定,测定的指标有胸径和树高,其中胸径使用钢围尺测量,树高使用测杆测量。香椿单株材积采用V=0.000 052 764 291D1.882161H1.0093166公式计算,其中D为胸径、H为树高[8,11]。
1.2.3 数据分析使用Excel 2007 进行数据录入和整理,使用Origin Pro 8 软件制图。采用SAS 8.1进行方差分析和方差分量估算[20]。
性状分析使用混合线性模型Yijk=U+Bi+Fj+BFij+Eijk,其中Yijk为第i个区组第j个家系第k个单株观测值;U为群体平均效应;Bi为第i个区组效应;Fj为第j家系效应;BFij为第i家系和第j区组的互作效应;Eijk为机误。其中区组为固定效应,其余为随机效应[21-22]。
单株遗传力为hs2=4σF2/(σE2+σ2FB+σF2),家系遗传力为hs2=σF2/(σE2/(nhb)+σ2FB/b+σF2),其中σF2为家系方差分量,σE2为误差方差分量,σ2FB为家系和区组互作的方差分量,nh为小区调和平均数,b为区组数[20]。
表型变异系数为CVp=σp/x×100%,遗传变异系数为其中σp为性状的表型标准差,σg2为遗传方差分量,x为性状的群体均值[21]。
现实增益遗传为G=(Xi-x)/x×100%,遗传增益为ΔG=(Xi-X)hs2/x×100%,其中Xi为入选家均值[23]。
采用综合育种值评选法评选香椿优良家系,育种值为Z=x+hs2(Xi-x)[23-24]。
2 结果与分析
2.1 香椿家系生长性状变异
香椿家系间造林1、2 a 的树高、胸径和材积都存在极显著差异(p<0.01)(表2)。造林1 a树高、胸径和材积的均值分别为3.59 m、4.49 cm和 0.003 581 m3,最大家系分别是最小家系的1.52、1.32 和2.28 倍;造林2 a 树高、胸径和材积的均值分别为7.03 m、7.68 cm 和0.018 672 m3,最大家系分别是最小家系的1.36、1.21 和1.86 倍。这说明不同香椿家系间的生长存在较大的差异,为优良家系的选择提供了基础。本研究中香椿造林2 a 的树高、胸径和材积均值分别是造林1 a 的1.96、1.71 和5.21 倍,说明材积的增长率最大,其次为树高,再次为胸径。
通过统计生长性状方差分量百分比(表3)可知,造林1 a 香椿生长性状的家系效应平均占比为10.94%,其中树高12.60%、胸径8.93%、材积11.30%;造林2 年香椿生长性状的家系效应平均占比为10.76%,其中树高15.06%、胸径6.88%、材积10.35%;造林1、2 a 年,家系效应均表现为树高>材积>胸径。
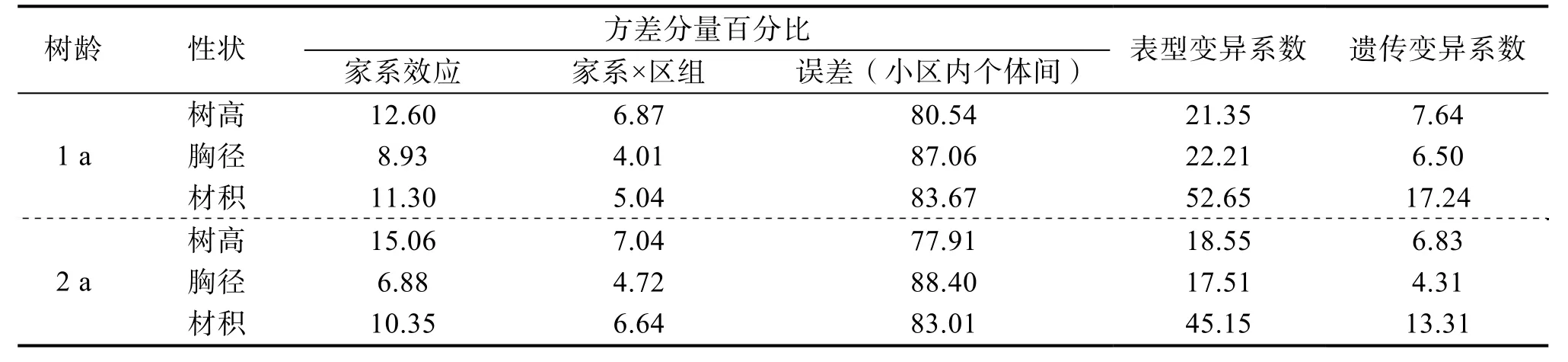
表3 香椿家系性状方差分量百分比和变异系数 单位:%
造林1 a 香椿的树高、胸径和材积的表型变异系数分别为21.35%、22.21%和52.65%,遗传变异系数分别为7.64%、6.50%和17.24%;造林2 a香椿的树高、胸径和材积的表型变异系数分别为18.55%、17.51%和45.15%,遗传变异系数分别为6.83%、4.31%和13.31%。造林1、2 a 香椿树高、胸径和材积的表型变异系数均大于遗传变异系数,这有利于香椿速生优良家系的选择。材积的变异系数均远大于树高和胸径,说明材积的性状最不稳定,选择的潜力最大;而树高、胸径的性状较稳定,但选择的潜力较小。
2.2 香椿家系生长性状的遗传力
由图1 可知,造林1 a 香椿的树高、胸径和材积的单株遗传力分别为0.50、0.36 和0.45,家系遗传力分别为0.82、0.80 和0.82;造林2 a 的树高、胸径和材积的单株遗传力分别为0.60、0.28和0.41,家系遗传力分别为0.84、0.74 和0.79。香椿的单株遗传力受中度遗传控制,家系遗传力受高度遗传控制,家系遗传控制强于单株遗传控制,这为香椿优良家系的选择提供了可能。
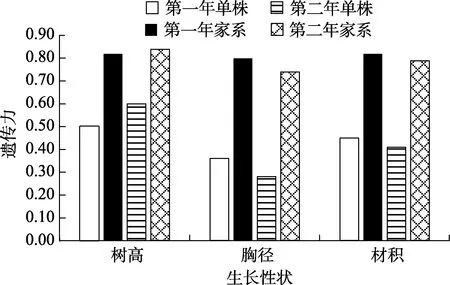
图1 香椿树高、胸径和材积的遗传力
2.3 香椿优良家系选择
采用育种值评估法以20%的入选率分别对造林1、2 a 的香椿进行优良家系选择,分别选出了7 个家系(表4、5),其中家系13、15、6、16、26 重复入选,重复入选率为71.43%,说明这5 个家系造林1、2 a 的生长量均较大,因而初步定为优良家系。入选的优良家系造林1 a 的胸径、树高、材积平均值分别为4.91 cm、4.05 m 和0.004 63 m3,造林2 a 的胸径、树高和材积平均值分别为8.21 cm、7.53 m 和0.022 4 m3。造林1、2 a 最好的家系均为13,造林1 a 的胸径、树高、材积平均值分别为4.94 cm、4.35 m 和0.005 13 m3,造林2 年的胸径、树高、材积平均值分别为8.33 cm、8.06 m和0.024 6 m3。
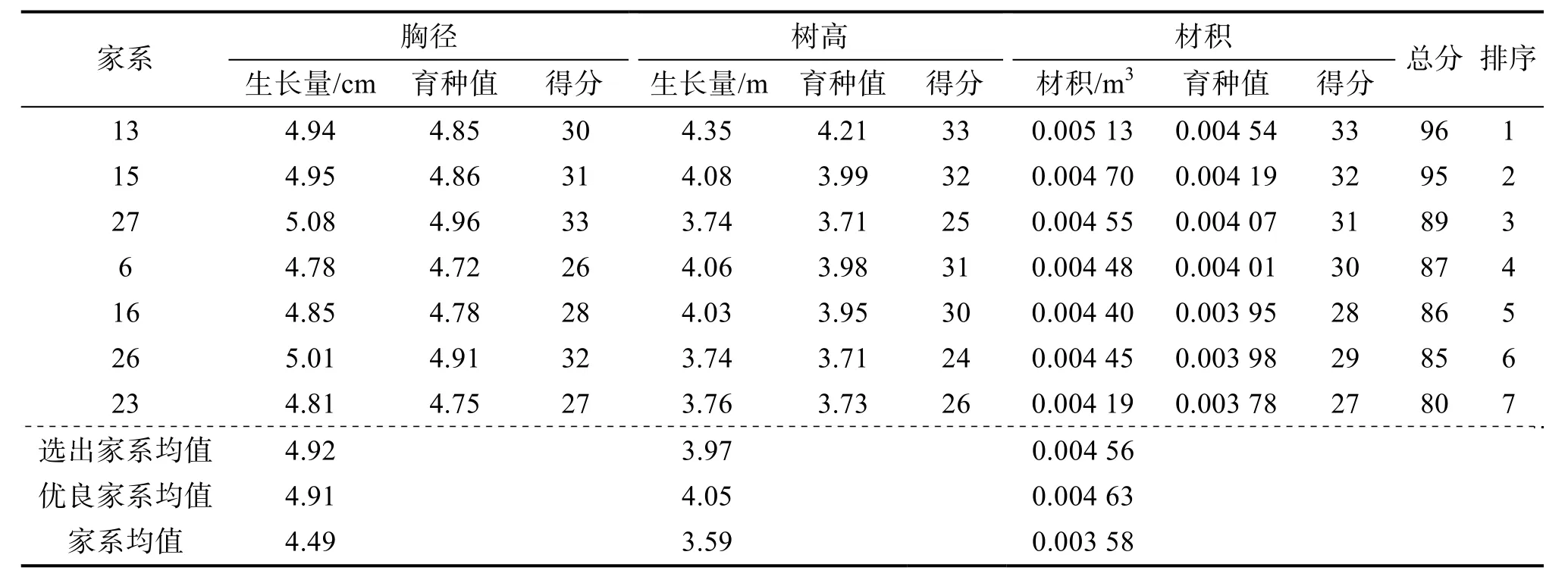
表4 造林1 a 香椿优良家系综合性状育种值分析
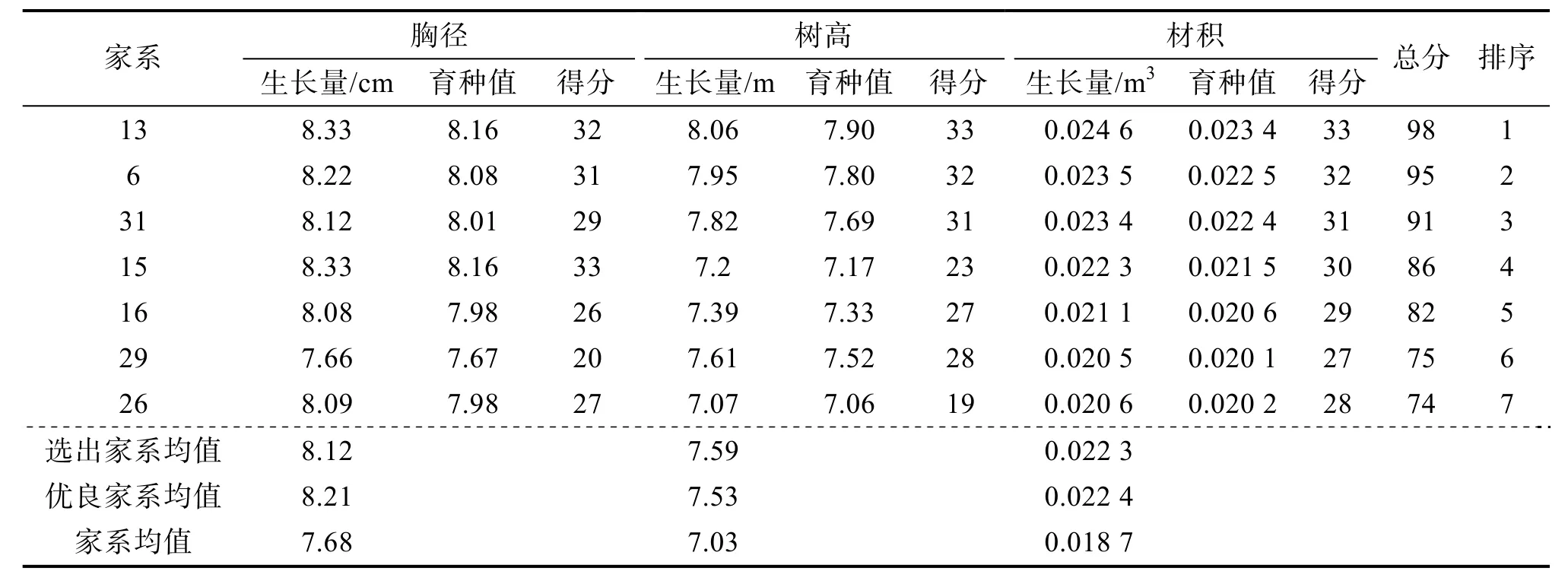
表5 造林2 a 香椿优良家系综合性状育种值分析
2.4 香椿优良家系遗传增益
由表6 可知,入选的5 个香椿优良家系造林第1 年胸径、树高和材积的现实遗传增益均值分别为9.27%、12.87%和29.34%,遗传增益均值分别为7.60%、10.30%和24.06%。家系26 胸径的现实遗传增益和遗传增益最大,分别为11.58%和9.50%;家系13 树高的现实遗传增益和遗传增益最大,分别为21.17%和16.94%;家系13 材积的现实遗传增益和遗传增益最大,分别为43.23%和35.45%。现实遗传增益和遗传增益均表现为材积>树高>胸径。
由表7 可知,入选的5 个香椿优良家系造林第2 年胸径、树高和材积的现实遗传增益均值分别为6.90%、7.17%和19.90%,遗传增益均值分别为5.80%、5.31%和15.72%。家系13、15 胸径的现实遗传增益和遗传增益最大,分别为8.46%和7.11%;家系13 树高的现实遗传增益和遗传增益最大,分别为14.65%和10.84%;家系13 材积的现实遗传增益和遗传增益最大,分别为31.79%和25.12%。现实遗传增益和遗传增益均表现为材积>树高≈胸径。
3 讨论与结论
选用广西33 个香椿优树的半同胞子代在广西南宁市营建试验林,结果表明,不同家系间造林1、2 a 的树高、胸径和材积都存在极显著差异,与代锐等[6]和肖兴翠等[7]研究结论一致;香椿造林1 年的生长指标优于张海燕等[9]研究中香椿造林4 a 的生长指标,造林2 a 的生长指标优于代锐等[6]和胡继文等[11]研究中香椿造林3 a 的生长指标,可能原因为广西气候土壤等条件更有利于香椿速生。香椿造林1~2 a,材积的增长率最大,其次为树高,再次为胸径;家系效应均表现为树高>材积>胸径,与木荷的一致[21]。本研究中,香椿造林1、2 a 树高、胸径、材积的表型变异系数与代锐等[6]和肖兴翠等[7]相似;表型变异系数均大于遗传变异系数,与欧阳天林等[21]对木荷家系的研究一致,这有利于香椿优良家系的选择;材积的表型变异系数和遗传变异系数均大于树高和胸径,而遗传变异系数高往往带来较大的选择响应[25],说明材积选择的潜力最大。
本研究中,香椿造林1、2 a 树高、胸径和材积的单株遗传力在 0.28~0.60,家系遗传力在0.74~0.84,家系遗传力大于单株遗传力,这与在木荷[21]、樟树家系[26]上的研究结果相似,说明香椿的树高、胸径和材积受到较强的遗传控制,这有利于香椿优良家系的选择。
本研究采用育种值评估法最终以15%的入选率,选出造林1 和2 a 重复入选的5 个优良家系,分别为13、15、6、16、26,其中家系13 的表现最好,生长指标(除第一年胸径外)的遗传增益均排在第一。优良家系造林1 年的胸径、树高、材积的遗传增益均值分别为 7.60%、10.30%、24.06%,造林2 年的的胸径、树高、材积的遗传增益均值分别为5.80%、5.31%、15.72%,说明优良家系具有良好的遗传改良潜力。本研究选出的香椿优良家系生长性状的遗传增益均值低于代锐等[6]和肖兴翠等[7]的,其原因为本研究中优良家系的入选率较大,导致选择差较小,遗传增益较小。
本研究中的试验林尚处于幼龄阶段,早期选择可能存在不稳定性,需继续对试验林进行观测,才能做出更全面的评价研究。
猜你喜欢
养猪(2022年4期)2022-08-17
农民致富之友(2017年4期)2017-04-10
现代农业科技(2017年4期)2017-04-10
绿色科技(2017年1期)2017-03-01
西南农业学报(2016年6期)2016-04-16
湖北畜牧兽医(2015年11期)2016-01-11
武夷学院学报(2015年3期)2015-07-18
山东农业科学(2014年1期)2015-03-09
湖北农业科学(2014年3期)2014-07-21
长春工程学院学报(自然科学版)(2013年4期)2013-12-06
