
节律性听觉刺激下的神经振荡同步化及其应用*
2023-06-20 04:42李经纬杨皓宇吕雪靖
生物化学与生物物理进展 2023年6期
李经纬 杨皓宇 胡 理 吕雪靖
(1)中国科学院心理研究所,中国科学院心理健康重点实验室,北京 100101;2)中国科学院大学心理系,北京 100049)
1924 年,Hans Berger 第一次利用脑电技术记录到人脑内的节律性电活动,并将观测到的以10 Hz 频率振荡为主要特征的脑电活动称为alpha波[1]。随后,研究者陆续发现了其他频段的脑电活动,并逐渐意识到这类脑电活动是大脑行使功能的一种基本活动模式[2],源于神经元重复的节律性放电活动在细胞集群中的整合。这种在皮层水平表现为一定频率范围内的节律性电活动被称为神经振荡(neural oscillation)[3]。
在安静无外界刺激时,大脑活动并非完全静止,神经元仍保持活动并维持相互之间的交流。如当个体在闭眼且安静状态下,可以在其枕叶处观察到明显的alpha 振荡(8~12 Hz),这反映了一种皮层的默认活动模式[1,4]。当处理信息或受到外界刺激时,脑内的默认神经振荡模式也会相应发生变化,反映了某些与刺激加工相关的特定心理过程。例如,视皮层alpha 振荡的能量和相位与个体对视觉刺激的探测能力有关[5-6],通过调控个体视皮层alpha 振荡,可以显著改善视力受损个体的视野探测缺陷,提高对视觉刺激的探测能力[7-8]。此外,记忆与theta 频段、认知功能与gamma 频段神经振荡之间的联系也被许多研究证实:theta 频段的神经振荡在工作记忆、情景记忆、陈述性记忆的形成中都起着关键作用,在海马、海马旁回等与记忆相关的皮质区域也可以观察到theta 频段的局部场电位活动[9];gamma 振荡被认为是大脑行使认知功能的一般活动模式[10],信息处理速度、工作记忆、抽象推理等认知功能的行使往往伴随着高频gamma 活动,在精神分裂症、阿尔茨海默病等认知功能受损的个体中也观察到gamma 活动的异常[11-12]。这些神经振荡模式与特定功能之间的联系以及神经振荡的可调控性,提示或许可以通过某些外界干预的方式影响与某些功能相关的振荡模式,调控行为任务表现,甚至在某些疾病的治疗上提供帮助。例如,通过呈现特定频率的外界刺激,可以引起大脑内相应频率的神经振荡的变化,达到内部振荡与外界刺激节律同步的效果,这种现象在神经生理学中被称为节律同步化(entrainment),也可以称为同步(synchronization)或相位锁定(phase locking)[13]。
目前,基于外界节律性刺激调控大脑神经振荡的方式主要有3 类,侵入性刺激(如深部脑刺激、光遗传技术等)、非侵入性刺激[14](如经颅电刺激、经颅磁刺激、超声刺激等)和节律性感觉刺激(如由单音组成的声音序列)。前两类是通过电/磁刺激等直接改变皮层神经元的活动模式,虽更加直接,但由于两者均采用外部刺激直接作用于皮层,难以还原信息在外周神经和皮层间的传递过程,而且还可能引起神经元活动的额外变化,难以确定其效果是否完全与诱发的神经振荡变化有关。此外,该类手段的实施依赖于专业的刺激器,设备成本高、操作复杂[15]。相比之下,节律性感觉刺激简单便携、可操纵性更强、成本更低,能还原信息传递的过程且不会引起额外的神经元变化,具有较好的应用前景。例如,利用节律性视觉刺激诱发神经振荡节律同步化,可以有效提高个体对特定相位出现目标刺激的探测正确率[16-17]。类似地,节律性听觉刺激诱发的神经振荡节律同步化也被证实可以有效调控认知行为和情绪状态等心理过程[18-19]。视觉、触觉、听觉等不同感觉通道的节律性刺激都能诱发节律同步化,相比之下,听觉刺激的时间分辨率更高,刺激期间被试活动更为方便,占据注意资源少,越来越得到研究者的关注[20]。
为明确外源节律性感觉刺激对神经元活动的作用机制及其对认知行为和情绪状态的调控效果,本文以不同类型的听觉刺激为例,在介绍神经振荡与节律同步化现象的基础上,综述节律同步化的电生理特点及其与行为的联系,讨论听觉刺激诱发神经振荡节律同步化的相关生理机制及应用前景,并对这一领域的后续研究方向提出建议。
1 听觉刺激诱发的节律同步化现象
无论是呈现间隔固定的序列刺激,还是振幅或者频率以一定规律变化的持续性调幅或调频刺激,或是左右声道由不同频率声音构成的双耳差频刺激,都可以看作是节律性听觉刺激,能够诱发大脑节律同步化现象,并影响个体的行为表现与情绪状态,但不同类型的刺激在不同频段的调控作用存在差异(表1)。在实际使用时,多采用被试感受舒适的刺激强度(如50 dB/SPL)呈现刺激。

Table 1 The regulatory effect of rhythmic auditory stimulations at different frequencies表1 不同刺激类型在不同频段的调控作用
1.1 序列刺激诱发的节律同步化
序列刺激以固定的间隔重复呈现某种声音刺激(如点击声),形成特定的呈现频率[21](图1a)。研究显示,各个频段的序列刺激都可以诱发大脑节律同步化,即在特定脑区的皮层水平观测到神经振荡频率与外界刺激频率趋同的现象[22]。从节律同步化产生的位置来看,当刺激呈现频率低于60 Hz时,序列刺激诱发的节律同步化现象主要可以在听觉皮层被观测到,并可随作用时间延长扩展至其他皮层区域[23-25],当刺激呈现频率高于60 Hz时,节律同步化现象则难以在皮层观察到,但在脑干及低位中脑水平可见[26]。
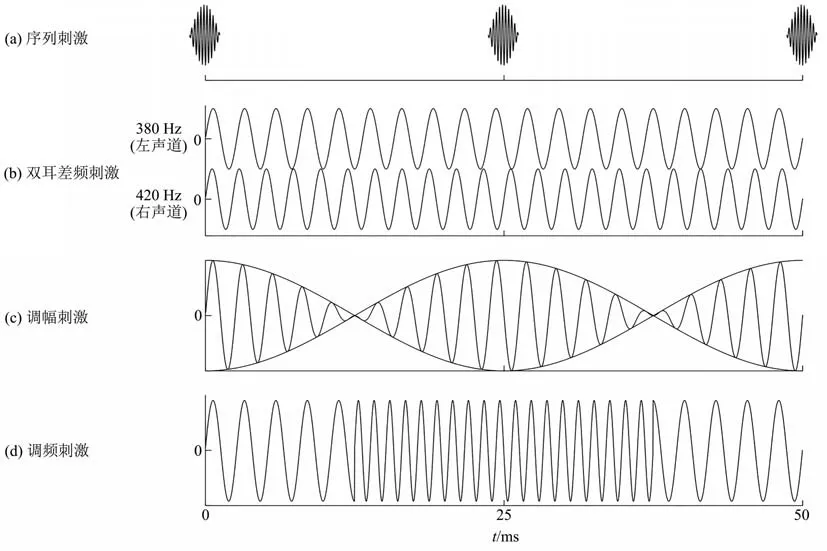
Fig. 1 The illustration of rhythmic auditory stimuli图1 节律性听觉刺激示例
低频呈现序列刺激的一个重要特性是可以引起注意产生周期性波动并影响知觉加工,在序列刺激的引导下,个体会对刺激出现的位置产生预期并将注意力集中。一个典型的例子来自Jones 等[27]的研究,研究者首先向被试呈现一个150 ms的纯音,然后以1.6 Hz 的频率序列呈现多个60 ms 的纯音。在声音序列的最后,呈现一个与第一个声音持续时间相同(150 ms)但频率不同的纯音作为目标刺激,该目标刺激可能出现在声音序列的不同相位。任务要求被试比较目标刺激与序列的第一个纯音的音高。结果显示,被试对在不同相位出现的目标刺激音高判断准确率有显著差异,表现为对预期相位出现的目标刺激音高判断准确率最高。类似地,当向被试以delta 频率(1~4 Hz)呈现序列刺激并同步记录脑电图(electroencephalography,EEG)数据时,Stefanics 等[28]不仅观察到大脑delta 频段的神经振荡显著增强,也发现目标刺激的探测反应时与目标刺激出现时皮层delta振荡的相位显著相关。这些结果提示,低频呈现序列刺激在注意调节中发挥着重要作用,其诱发的节律同步化与注意波动有极为明显的相位一致性,即节律性刺激可以影响神经元的同步活动并引导注意向预期下一个刺激出现的时间点集中,使得个体对刺激的探测速度增加,并提高阈限附近刺激的探测准确率[29-31]。
此外,研究者发现,以theta 频段(4~8 Hz)呈现的低频序列刺激,有助于改善个体的记忆表现,这主要是由于海马的theta振荡在记忆的编码、巩固和提取中均发挥着重要作用[32-33]。在一项再认任务中,被试在记忆的编码阶段分别暴露在白噪声、5.5 Hz 和14 Hz 的节律性声音刺激中,并同步记录EEG 数据,结果显示暴露在节律性刺激中的被试均表现出对应频段的节律同步化,且5.5 Hz组的被试在记忆任务中的表现显著优于其他组[34]。这种低频刺激诱发的同步化对记忆的影响也表现在记忆的整合能力中,当视觉刺激和听觉刺激同时以4 Hz 频率呈现时,个体可以更好地将两种刺激联系起来,形成更为牢固的情景记忆[35],且即使在睡眠阶段施加外部序列刺激,也会对记忆整合能力产生积极的影响[36]。
相比于低频呈现,高频呈现的序列刺激(尤其是40 Hz)可以诱发更强的同步化响应[25]。相较于瞬时的听觉诱发电位,这种持续性的刺激更加稳定,更易观测。因此,40 Hz节律同步化现象最早被观察到并被称为40 Hz 听觉稳态响应(auditory steady state response,ASSR)[37-38]。与低频呈现的序列刺激相比,40 Hz呈现的序列刺激诱发的节律同步化易受个体生理状态的影响。例如,在睡眠或麻醉状态下,丘脑对听觉信息进行上行抑制,节律性刺激诱发的同步化响应显著弱于清醒状态,且在40 Hz频段变化最大[39-41]。此外,高频刺激诱发的节律同步化还受个体认知状态[42]以及选择性注意和工作负荷等认知因素[43-44]的影响,因此,ASSR也可能反映了个体一般性的认知功能网络连接的密切程度[12]。
综上所述,低频序列刺激及其诱发的神经节律同步化响应在调节注意、记忆等认知功能方面有重要作用。相比之下,高频序列刺激诱发下的神经节律同步化响应则多作为个体生理状态的有关指标发挥指示性作用。
1.2 双耳差频刺激诱发的节律同步化
双耳差频刺激(binaural beats)是通过向双耳呈现不同频率的声音(如左声道380 Hz,右声道420 Hz),形成以差频(如40 Hz)为主要听觉感受的听觉刺激(图1b)。换言之,在主观感受下,人们不仅会感受到一个载波(380 Hz),还可以感知到一个差频波(40 Hz)的声音。当差频小于2 Hz时,人们会感知到一个在左右耳之间往复移动的声音。随着差频的提高,这种往复运动的感觉消失,但仍可以感知到一个出现在头部中央的持续声音刺激[45]。在脑响应上,低频的双耳差频刺激在听皮层、顶叶、额叶会表现出明显的对应频率及其谐波频率的神经振荡同步化[46-47],与序列呈现的听觉刺激诱发同步化的区域没有显著差异[48]。
双耳差频刺激最早被用于缓解患者术前焦虑和手术后的疼痛。例如,在手术前使用30 min delta频段的双耳差频刺激可以有效降低患者的术前焦虑及术后的止痛药物用量[49]。近年来的研究也显示,delta频段、alpha频段的双耳差频刺激在牙科手术、膝关节置换术、白内障手术中有显著减少止痛药用量、缓解焦虑的作用[50-52]。除此之外,在睡眠中对健康个体施以delta 频段的双耳差频刺激可以影响睡眠状态,使个体更容易进入深度睡眠,并使睡眠循环中深度睡眠时间延长[53],这提示双耳差频刺激的治疗作用还可以应用于普通人群。后续研究也发现,不同频段的双耳差频刺激与不同生理功能有密切的联系。例如,施以theta 频段的双耳差频刺激可以诱导个体进入与冥想类似的皮层活动状态,并影响自主神经系统活动,减少焦虑情绪,增强放松感[54-55]。又如,alpha 频段的双耳差频刺激可以在提高认知功能方面发挥作用——减少个体探测听觉刺激和视觉刺激的反应时,并有助于个体发挥创造力,增强发散性思维[56],但对记忆表现没有影响[57]。相比之下,beta 频段(13~30 Hz)的双耳差频刺激对记忆有积极影响,在15 min 的双耳差频刺激后,要求被试对词语进行记忆并在分心任务后自由回忆,结果显示接受了beta频段双耳差频刺激的被试成绩显著好于接受白噪声刺激的被试,而接受较低的theta频段刺激的被试成绩甚至更差[58]。以上结果提示,低频差频刺激下认知功能与情绪状态的改善可能对差频频率具有偏好性。
与序列呈现刺激的方式相同,高频双耳差频刺激下的节律同步化的诱发相较于低频段也更为容易,甚至有研究指出大多数频段的双耳差频刺激都会诱发40 Hz的振荡响应[19]。这些同步化响应最先在听皮层、顶叶以及额叶区域被观测到[59],随作用时间的增加可进一步在全脑观测到同步化响应[23]。与低频双耳差频刺激调控个体情绪状态相比,高频双耳差频刺激对个体的影响更多的表现在对认知功能的调控上。在行为上,高频差频刺激可以提高个体的认知任务表现,这可能与大脑高频振荡与认知功能的普遍联系有关[12]。例如,40 Hz高频双耳差频刺激可以提高个体在视觉和听觉探测任务的反应速度[57]和在词汇回忆任务的表现[60];在注意瞬脱实验过程中施加40 Hz双耳差频刺激,经睡眠巩固后,可以在第2 天有效改善注意瞬脱现象,但在施以较低频段(16 Hz)的双耳差频刺激后则没有这种效果[59]。
双耳差频刺激诱发节律同步化响应还依赖于外界刺激呈现时长。例如,有研究分别向被试呈现多个频段的双耳差频刺激,但每个刺激仅呈现3 min,结果显示所有频段刺激均不能诱发节律同步化响应,也对个体情绪没有影响[61]。类似地,一段时间内间隔施加双耳差频刺激(如持续1 min,间隔1 min,共20 min 的双耳差频刺激)也同样无法诱发节律同步化现象[62-63]。相比之下,持续6 min左右的刺激可以在听觉皮层及其附近区域诱发一定程度的节律同步化,持续10 min 左右的双耳差频刺激则可以诱发较大范围甚至扩展至全脑的节律同步化活动[23,54,64-65],这可能是由于随时间延长会有更多的神经元活动被调节至刺激频率,使得节律同步化范围扩大,提示刺激时长对节律同步化的影响与节律同步化诱发的生理机制有关。
由此可见,低频双耳差频刺激多用于情绪调控,高频双耳差频刺激则多用于认知调控,而且调控效果在一定程度上依赖于调控时长,表现为持续一段时间的双耳差频刺激才能诱发稳定的神经节律同步化响应,从而影响个体的情绪状态和认知表现。
1.3 调幅和调频刺激诱发的节律同步化
调幅刺激(amplitude modulated beats)是将听觉信号的振幅以一定规律调节产生的刺激(图1c)。除了将振幅以不同频率的正弦函数调制,调幅刺激也可通过将两个有一定频率差的声音刺激合成为单声道刺激(monaural beats)实现。在已有文献中,调幅刺激和单声道刺激经常会相互使用,虽然名称不同,但其物理学本质相同。除此之外,将声音频率按照一定规律调节可以产生调频刺激(frequency modulated beats)(图1d)。调频刺激也可以影响皮层的神经振荡模式[66-67],大脑对调频刺激的响应区域与对调幅刺激也无显著差异[68]。调幅刺激与调频刺激组合,加上停顿、节奏等便成为了音乐。音乐在情绪调控、改善认知方面的作用已被广泛证明,本团队的一项元分析研究也表明[69],音乐通过对注意、情绪的调节作用,可以对实验性的疼痛产生中等程度的镇痛效果,可以作为疼痛管理的辅助工具。由于音乐本身是一种复杂的声音刺激在时间维度的延展,且具有强烈的个人偏好,因此不作为本文重点讨论。
与序列和双耳差频声音刺激一样,调幅刺激同样可以诱发神经节律同步化,其活动最强的区域主要位于听觉皮层,并可扩展至顶叶、额叶、枕叶乃至全脑[23]。有研究对比了3、6、40 Hz等多频段下调幅刺激和双耳差频刺激的节律同步化响应,其结果显示两种刺激诱发的节律同步化的头皮分布上均没有区别[48,70],但调幅刺激诱发的神经响应更强[71]。目前针对调幅和调频刺激调节个体的情绪和认知的研究较少。与序列刺激类似,低频的调幅刺激也有对注意的节律性调节作用。对于出现在不同相位的探测目标,个体的探测能力有明显的差异,施加调幅刺激可以诱发节律同步化并引起个体的注意波动,提高个体对出现在特定相位刺激的探测能力[72-73]。此外,theta、alpha 和gamma 频段的调幅刺激可以缓解焦虑情绪,但对工作记忆、长时记忆、警觉性的作用效果不显著[74],甚至会降低短时记忆能力[75]。40 Hz的调幅刺激也可以增强个体的认知能力,减少被试在注意任务中的反应时[76]。频率不断变化的调频刺激难以诱发特定频段的节律同步化,较少作为神经调控的工具调控个体的行为表现,但调频刺激在听觉系统加工中被认为有特殊意义。如调频刺激在听力测试中有较高的敏感性,可用于测试更大范围的听觉能力[77],通过定制个性化的调频系统,可以帮助听觉加工系统受损患者提高语义理解能力等[78]。
综上所述,调幅刺激本身有着与序列刺激和双耳差频刺激类似的作用,可用于诱发多频段的节律同步化,调频刺激也在听觉系统中有特殊的作用,但两者在神经调控方面的研究较为初步,需要更多的证据来进一步证明其效果。
2 听觉刺激诱发节律同步化的机制
2.1 节律性感觉刺激诱发大脑神经振荡节律同步化理论
研究者们提出了不同理论解释外界刺激如何逐步影响皮层的神经振荡模式,如相位重置理论、叠加理论等,其中最具影响力的是Thut等[13]提出的节律同步化模型。该模型认为,神经元活动有其自发的、可持续的振荡模式,在没有外界刺激的情况下,神经元以该模式自发活动,但当外界节律性刺激作用于神经元,神经元的自发振荡模式就会改变,使神经元的振荡模式与外界刺激越来越一致,直至与外界刺激完全一致。当受影响的神经元越来越多,这一细胞集群便表现为与外界刺激一致的活动 模 式 , 并 可 以 被 EEG、 脑 磁 图(magnetoencephalography,MEG)等神经科学手段观测到。因此,通过调节外部刺激的参数可以影响神经元的活动,表现为通过改变外部刺激的振幅、相位、呈现频率和作用时间等调节相应神经元的活动。
2.2 节律性感觉刺激诱发大脑神经振荡节律同步化的生理过程
从生理过程来看,不同的节律性听觉刺激都通过听觉系统发挥作用,在听觉皮层表现出节律同步化响应,并扩展至顶叶、额叶等区域。然而,在皮层下水平,不同类型的刺激初步产生节律性神经反应的结构水平并不相同。具体而言,个体的听觉系统利用外化的耳廓等结构稳定地捕捉到声音的频率信息,声音信号经耳膜、听小骨、纤毛等结构转换为电信号,进一步从耳蜗听神经传至耳蜗腹前核,并经由轴突连接上橄榄核。位于脑干两侧的上橄榄核对于声音的相位信息尤其敏感,并相互影响,例如当双耳捕获到的声音频率不同时,上橄榄核产生的膜电位便受到调制[79]。上橄榄核的放电模式进而影响其轴突连接的下丘神经元,对双耳差频刺激而言,下丘神经元可整合两耳听觉信息以反映双耳差频刺激的差频特性[45]。于是,双耳听觉刺激在脑中被合成为类似调幅刺激的听觉感知,并进一步影响大脑皮层,产生可以在皮层观察到的节律同步化现象[71]。由此可见,双耳差频刺激与调幅刺激和序列刺激的差异在于:前者产生调节作用诱发节律性活动的位置在上橄榄核,而后者在耳蜗水平便可产生节律性的神经活动[74]。
2.3 节律性感觉刺激诱发大脑神经振荡节律同步化的认知过程
从认知过程来看,个体在处理不同类型的节律性听觉刺激时所涉及的加工过程并不相同。以注意的节律性波动为例,许多研究均表明注意的节律性波动与刺激诱发的节律同步化有密切关系[13,72]。然而,任何一个被主观意识到的外界刺激都会与注意机制有关,提示节律性感觉刺激诱发的神经振荡同步化不仅受输入刺激本身的属性影响,还可能受自上而下的注意调控。例如,注意可以影响诱发的节律化响应的强度。有研究在猴视觉皮层内插入电极,同时向其呈现节律性视觉刺激和听觉刺激,并训练猴注意特定的感觉通道,颅内脑电结果显示,在仅注意听觉通道时,视觉节律性刺激诱发的同步化响应会显著降低,而在注意视觉通道时,响应则明显提高[80]。此外,个体可以主动加工刺激产生特定频段的节律同步化。对一串序列刺激,有音乐训练经验的个体可以根据指导语主动组合刺激,将两个纯音或三个纯音组合为一组,形成二拍或三拍的节律,由此将2.4 Hz的节律性刺激知觉为1.2 Hz或0.8 Hz。在3 种条件下,被试会有不同的节律同步化响应模式:不进行主观加工条件下仅能观察到顶叶、颞叶2.4Hz的节律同步化活动,而主观加工条件下,除了2.4 Hz,也可以在顶叶、额叶观察到1.2 Hz和0.8 Hz的节律同步化活动[81]。
3 总结与展望
节律性听觉刺激诱发神经振荡同步化是一种广泛的现象。目前已有大量研究关注了该现象的生物标记作用。例如,ASSR已被广泛应用于婴幼儿听力测试,作为评估听觉系统的客观指标。此外,由于40 Hz节律同步化响应易受生理状态影响,在不同状态下有较大差异(麻醉、睡眠状态相较于清醒状态),可将其作为麻醉状态的监测指标[82-83]。类似地,在40 Hz节律同步化响应还可以作为神经分裂症的生物标记,精神分裂症患者等认知功能受损的个体通常会表现出同步化响应功率减小、同步及去同步响应延迟现象[12,84]。
更重要的是,节律性听觉刺激具备调控大脑神经振荡模式,从而影响个体的行为表现的潜力,因此可以被看作是一种非侵入性的神经调控手段。虽然上文提到的节律性刺激对情绪和认知功能的调控尚处于实验室探索阶段,但目前已有多项研究将节律性刺激应用于临床治疗,尤其在运动功能康复上效果显著。研究表明,节律性刺激可以在帕金森病、卒中、创伤性脑损伤、多发性硬化等多种因素导致的运动功能损伤康复中发挥重要作用[85],例如,在帕金森病患者的运动训练过程中给予节律性刺激,经一段时间的训练后,相比于不接受节律性刺激的对照组,在训练中接受节律性听觉刺激可以有效提高步行速度和步长,增强步行稳定性,防止摔倒并减少步态冻结[86-87]。除运动康复方面,动物研究也表明,gamma 频段的节律性听觉刺激和节律性视听联合刺激可以减轻小鼠阿尔茨海默病的相关表现,改善空间记忆和再认记忆,并引起脑内神经元活动的相应变化,减少海马、额叶乃至全脑的淀粉样蛋白沉淀[88]。由调幅、调频等刺激组合的音乐刺激也被证明有在执行功能和注意功能方面有显著的康复作用[89],提示了节律性听觉刺激在更为广阔的领域的应用潜力。
然而,目前节律性刺激诱发神经振荡同步化的机制仍有待进一步研究。首先,不同频段的听觉刺激诱发的节律同步化与个体行为表现的联系仍不清晰。虽然目前的研究提示低频刺激与情绪调节、高频段与认知功能之间存在联系,但这种联系似乎并不稳定。这可能是由于实验中采取了不同的刺激参数,导致了结果的不一致性。因此未来的研究需要探究不同刺激参数下的调控效果,构建节律性刺激与神经振荡同步化以及行为表现之间的关联模型。其次,以往研究表明序列刺激、双耳差频刺激、调频/调幅刺激三类刺激在皮层水平可以诱发相似的生理响应,但调控效果却不完全相同。这可能是因为研究者在探究不同类型刺激的调控效果时,所关注的调控内容不同。例如,低频调幅刺激和序列刺激都可以引起注意的节律性波动,但现阶段仍缺乏对低频双耳差频刺激调控注意的研究。大量研究显示施加高频双耳差频刺激和调幅刺激对认知功能有影响,却鲜有研究探索高频序列刺激对认知功能的影响,而是主要将其作为相关功能的生物标记信号。因此,未来研究应进一步统一相关证据或阐明各类刺激调控效果的差异,这将有助于加深对听觉刺激诱发节律同步化的认识。
综上所述,不同频段的节律性听觉刺激可以诱发相应的节律同步化活动,改变个体的神经振荡模式,进一步影响个体的疼痛感受、情绪、记忆和认知等功能。相较于其他侵入性或非侵入性的电/磁刺激,节律性听觉刺激有着成本低、易控制、简便易得等优势,是一种具有应用潜力的神经调控方式。未来有望在广泛人群中通过施加节律性听觉刺激对大脑神经振荡进行调控,以达到调控情绪与认知功能、辅助治疗相关疾病的目的。
猜你喜欢
家庭科学·新健康(2022年1期)2022-02-02
家庭科学·新健康(2021年11期)2021-11-23
紫禁城(2020年5期)2021-01-07
舰船电子工程(2019年6期)2019-07-08
电子制作(2019年9期)2019-05-30
测控技术(2018年6期)2018-11-25
传感器与微系统(2017年7期)2017-08-09
家庭科学·新健康(2016年11期)2016-11-23
新疆农垦科技(2016年10期)2016-06-15
河北大学学报(自然科学版)(2015年6期)2016-01-29
