
基于红外相机技术的海南长臂猿分布区林冠层伴生鸟兽多样性的研究
2023-06-15 07:17蒙金超杨雪珂冯悦恒齐旭明
热带生物学报 2023年3期
蒙金超,杨雪珂,冯悦恒,齐旭明,刘 辉
(1. 海南大学 林学院,海口,570208; 2. 海南热带雨林国家公园管理局,海口, 570203)
红外相机监测作为一种非损伤性的野生动物监测方法[1],具有昼夜连续工作、人力投入少、对研究对象干扰小等优势[2-3],目前已被广泛应用于动物生态学的研究[4]。通过分析红外相机所拍摄的动物图像数据,可以获得物种丰富度、空间分布、活动节律和种群密度等信息[5-6],能一定程度弥补传统调查方法的不足,尤其是针对一些夜间活动的、隐蔽的和珍稀物种的调查更为有效[7]。我国最早在云南省高黎贡山地区和台湾地区使用红外相机技术监测野生动物[8-9]。经过20 多年的发展,红外相机技术已发展成为地栖大中型兽类和鸟类的重要常规监测技术[10-11]。但该技术应用于高度树栖的林冠层动物多样性的研究相对较少,且布设相机台数少及安装高度较低,仅5~15 m[12],在林冠层中使用红外相机调查树栖哺乳动物的潜在效果在很大程度上仍是未知的。
海南长臂猿(Nomascus hainanus)为长臂猿科(Hylobatidae)、黑冠长臂猿属(Nomascus),是我国一级重点保护野生动物。由于在历史上受栖息地破坏及狩猎等因素导致海南长臂猿种群数量极低[13],2007 年10 月保护国际(CI)宣布海南长臂猿名列全球25 种濒危灵长类物种之首[14]。我国高度重视该物种的保护,基于自然恢复方法的科学保护使得海南长臂猿成为全球20 种长臂猿中唯一处于数量增长的类群[15],种群数量从20 世纪80 年代2 群7~9 只增至2022 年的5 群36 只[16],目前仅分布在海南热带雨林国家公园霸王岭片区斧头岭及南崩岭区域[17],属于高度树栖的昼行性小型猿类[18-19],以成熟果实、嫩叶、嫩芽为食[20-21]。
海南长臂猿作为海南热带雨林国家公园野生动植物保护的标志性物种和生态指示种[22],对于该物种的保护和研究有助于提升热带雨林生态系统原真性和完整性。目前对海南长臂猿的研究集中在种群现状、食性和栖息地等方面[23-24],缺少海南长臂猿同域分布区伴生鸟兽多样性组成及季节性变化相关研究。本研究拟使用红外相机技术分析海南长臂猿林冠层同域分布物种的多样性、年活动格局以及海拔分布格局,探讨海南长臂猿与其同域分布鸟兽物种的共存机制,为未来基于生物多样性的海南长臂猿栖息地修复和生态廊道建设提供参考。
1 研究方法
1.1 研究区域概况海南热带雨林国家公园霸王岭片区(以下简称为霸王岭片区)地处海南昌江县和白沙县境内(19°02′~19°08′N, 109°02′~109°13′E),总面积约850 km2,海拔75~1 655 m,年平均温度21.9 ℃,年平均相对湿度84.2%,年均降水量1 657 mm,属热带季风气候,旱季雨季分明,5—10 月为雨季,11 月至次年4 月为旱季,4—8 月常有台风;其核心保护区内覆盖有热带低地雨林、热带山地雨林、热带山地常绿阔叶林和热带山顶矮林[13]。国家一级重点保护动物有海南长臂猿、海南山鹧鸪(Arborophila ardens)、大灵猫(Viverra zibetha)、海南孔雀雉(Polyplectron katsumatae)等8 种,国家二级重点保护动物有水鹿(Rusa unicolor)、白鹇(Lophura nycthemera)、蟒蛇(Python bivittatus)、猕猴(Macaca mulatta tcheliensis)和巨松鼠 (Ratufa bicolor)等50 种[25]。
1.2 相机安装和数据收集根据海南长臂猿各个家庭群分布现状、海拔分布和猿食植物分布,选择5 个家庭群中的B、C、D 群分布区作为研究区域,面积约15 km2。B、C、D 群分布区主要为热带山地雨林,其他林型分布较少。2019 年1 月开始,布设33 个红外相机监测位点66 台红外相机(图1),红外相机安放主要考虑长臂猿实际利用区域(护林员记录的长臂猿活动强度及猿食植物分布),兼顾较高和较低海拔(600~800 m 热带低地雨林布设16 台,800~1 100 m 热带山地雨林布设40 台,1 100 m 热带苔藓林布设10 台)。将相机安装在离地13~25 m 的树冠层,选取海南长臂猿可能经过或者附近有猿食植物且视野开阔的树干进行安放,避免树枝遮挡镜头;镜头与地面呈5°~15°俯视角,避免阳光直射。每个监测位点安装的2 台相机机身相靠、镜头相反,各自反方向拍照,相邻位点之间的直线距离在100~500 m[26-27]。拍摄模式设置为连拍3 张照片同时录制视频10 s,拍摄间隔为0 s,灵敏度为中。安装前进行性能测试,安装时记录每个位点相机的编号、GPS 位点、海拔和生境因子等信息。
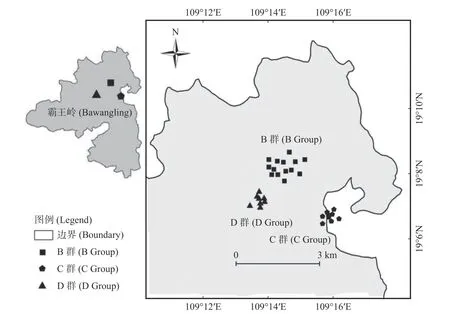
图1 海南长臂猿分布区林冠层红外相机监测点分布图
2019 年8 月,完成第一次红外相机的数据回收(替换SD 卡)和维护(更换电池),同时发现有11 台红外相机长时间受到雨水冲刷从而彻底损坏,最终实际正常工作的红外相机为55 台(海拔600~900 m 14 台,海拔900~1 100 m 31 台,海拔1 100 m 以上10 台)。2019 年11 月和2020 年6 月完成第二次和第三次数据回收,未有相机损坏。熟悉海南本土物种的专家人工对照片和视频进行物种识别,剔除模糊不清或没有明显物种特征的照片,所拍摄到照片和视频中的物种鉴定主要参考《中国鸟类分类与分布名录》和《中国哺乳动物多样性》[28-29]。
1.3 数据分析
1.3.1 海南长臂猿分布区林冠层物种组成利用相对丰富度指数衡量海南长臂猿分布区林冠层兽类和鸟类各物种相对种群数量。独立有效照片是指同一相机位点含同种个体的相邻有效照片( 或视频)间隔时间至少为 30 min[30],1 个相机工作日以单台红外相机在野外持续工作24 h 为标准,每台红外相机有效工作日以该位点拍摄到的第一张野外工作照片(显示有正确设置信息)和最后一张野外工作照片的日期间隔为有效工作日。将拍摄到各种鸟类和兽类物种的独立有效照片与所拍摄物种独立有效照片总数的百分比作为每个物种的相对丰富度(relative abundance index, RAI)[31]。
1.3.2 分布区内鸟兽年活动格局依据统计所得数据,计算月相对丰富度指数(monthly relative abundance index, MRAI),分析独立有效照片较多的动物年活动规律。
1.3.3 分布区内鸟兽海拔分布格局根据海南长臂猿栖息地类型和相机实际安装情况将研究区域分为3 个海拔梯度(600~800、 800~1 100、1 100 m 以上),并统计不同海拔梯度的独立有效照片,采用Shannon-Wiener 指数、Pielou 均匀度指数和Simpson 指数判断不同海拔梯度的α多样性水平[32]。
Shannon-Wiener 指数(H')是将物种丰富度与种的多度结合起来的指数,通常来说H'值越大,物种多样性越丰富。由于每个海拔梯度布设的红外相机台数的不同,在比较不同海拔梯度的物种数和多样性指数时,以每一台相机作为独立样本,计算每一台相机的物种数、多样性指数和均匀度指数,通过比较其平均值表示不同海拔梯度的物种多样性情况[33]。采用Kruskall-Wallis test 来比较不同海拔梯度物种多样性的差异,差异显著水平设定为0.05。所有数据都在SPSS 24.0 和Excel 2021上进行处理。
Pielou 均匀度指数(J)反映物种分布的均匀程度,能够表征分类单元中每个物种个体间的差异。均匀度(J)数值越大,群落中物种个体间的差异越小。
物种优势度(C)采用Simpson 指数计算,C 值越大说明群落内物种数量分布越不均匀,优势种地位越突出[34]。
2 结果与分析
2019-01-20—2020-06-30 期间,55 台红外相机累计监测到16 266 个有效相机工作日,共拍摄到兽类、鸟类照片和视频14 136 份,鉴定出独立有效照片1 747 张, 其中兽类1 569 张(89.8%)、鸟类178 张(10.2%)。记录到3 目4 科10 种兽类,6 目11 科20 种鸟类。拍摄到海南长臂猿(21 个)、海南鼯鼠(Petaurista hainana)(16 个)、红颊长吻松鼠(Dremomys rufigenis)(16 个)和果子狸(Paguma larvata)(15 个)的位点数居于前4 位,其他兽类和鸟类较少(表1)。
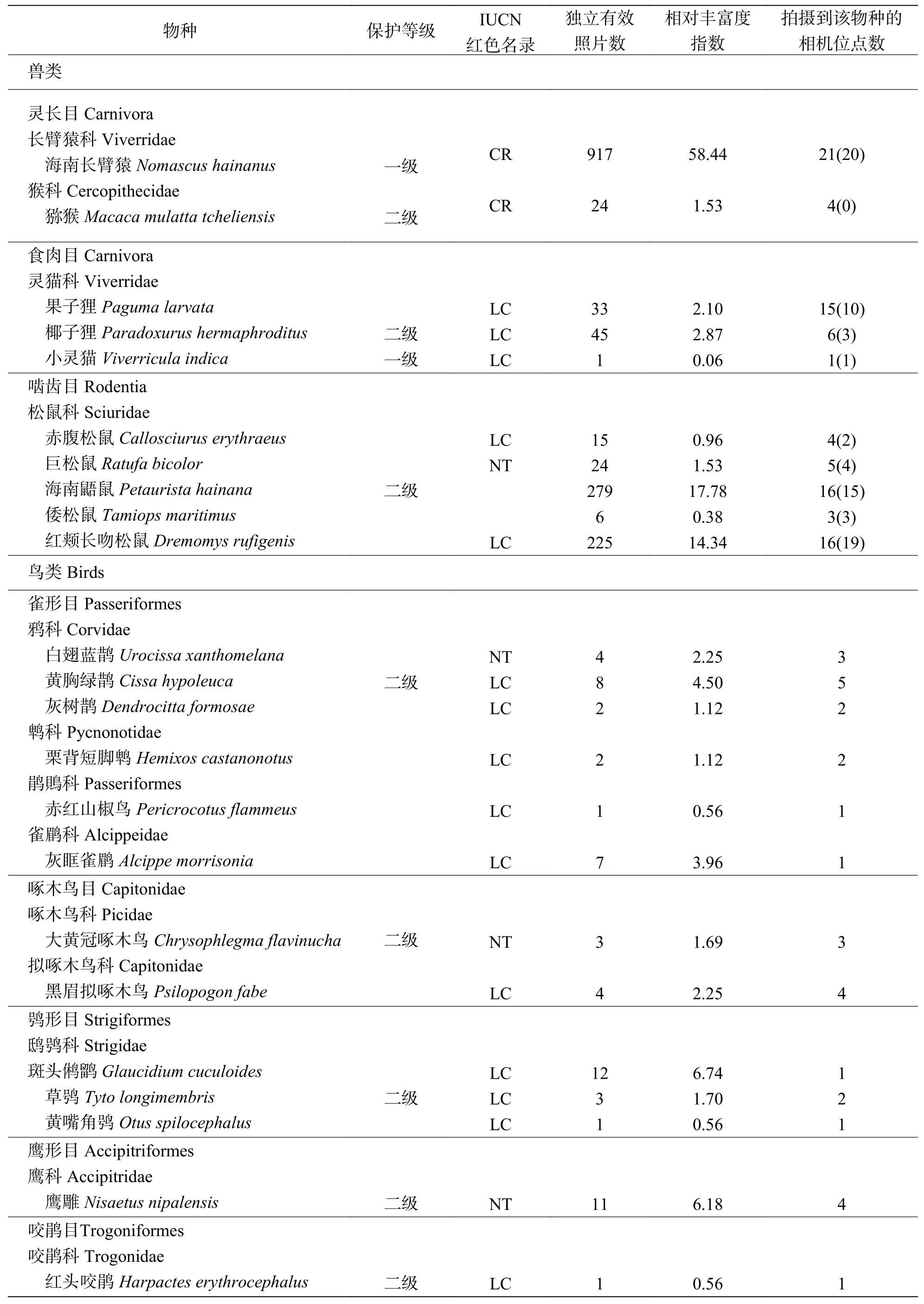
表1 海南长臂猿同域分布区林冠层兽类与鸟类动物名录
2.1 鸟兽相对丰富度记录到的10 种兽类中,国家一级重点保护动物2 种,即海南长臂猿和小灵猫(Viverricula indica);国家二级重点保护野生动物3 种,分别是猕猴、巨松鼠和椰子狸(Paradoxurus hermaphroditus),国家一、二级重点保护动物合计占所有兽类物种数的50%。海南长臂猿被IUCN 红色名录列为极危物种(CR),巨松鼠被列为近危物种(NT)。此外,兽类中相对丰富度指数居于前三的是海南长臂猿(RAI =58.44)、海南鼯鼠(RAI =17.78)和红颊长吻松鼠(RAI =14.34),其他兽类的相对丰富度指数均低于3,小灵猫仅有1 张独立有效照片(表1)。
记录到的17 种鸟类中,国家二级重点保护鸟类8 种,分别是黄胸绿鹊(Cissa hypoleuca)、大黄冠啄木鸟(Chrysophlegma flavinucha)、斑头鸺鹠(Glaucidium cuculoides)、鹰雕(Nisaetus nipalensis)、红头咬鹃(Harpactes erythrocephalus)、厚嘴绿鸠(Treron curvirostra)、山皇鸠(Ducula badia)、和蓝须蜂虎(Nyctyornis athertoni),占所有鸟类物种数的53%。记录到的17 种鸟类中,3 种为近危物种(NT),分别是白翅蓝鹊(Urocissa xanthomelana)、大黄冠啄木鸟和鹰雕。鸟类中相对丰富度指数居于前三的是山皇鸠(RAI=44.92)、厚嘴绿鸠(RAI=20.23)和斑头鸺鹠(RAI=6.74); 赤红山椒鸟(Pericrocotus flammeus)、红头咬鹃、黄嘴角鸮(Otus spilocephalus)和蓝须夜蜂虎的相对丰富度指数较低(表1)。
2.2 林冠层鸟兽年活动格局兽类各月份活动强度较高,鸟类则较低,在11、12 月份未拍摄到鸟类照片和视频。海南长臂猿和海南鼯鼠年活动格局相似,且与季节(旱季和雨季)显著相关(海南长臂猿:χ2= 8.308, df = 1,P= 0.002; 海南鼯鼠:χ2=3.124, df = 1,P= 0.047)。海南长臂猿在9 月份活动强度最低,10 月份快速上升,3 月份达到全年活动峰值,旱季则较低。海南鼯鼠在3 月份出现活动峰值,在5—10 月份趋于平缓。其他兽类如红颊长吻松鼠、椰子狸、倭松鼠(Tamiops maritimus)的年活动格局均与季节无显著相关,在各个月份的活动强度较为平缓。5 种鸟类的年活动格局均与季节无显著相关,其中山皇鸠和厚嘴绿鸠在1 月份出现活动峰值,斑头鸺鹠和白翅蓝鹊在8 月份出现峰值,黄胸绿鹊在4 月份出现峰值,其他月份鸟类月相对丰富度较低,主要集中在1—4 月份和7—8 月份活动(图2)。
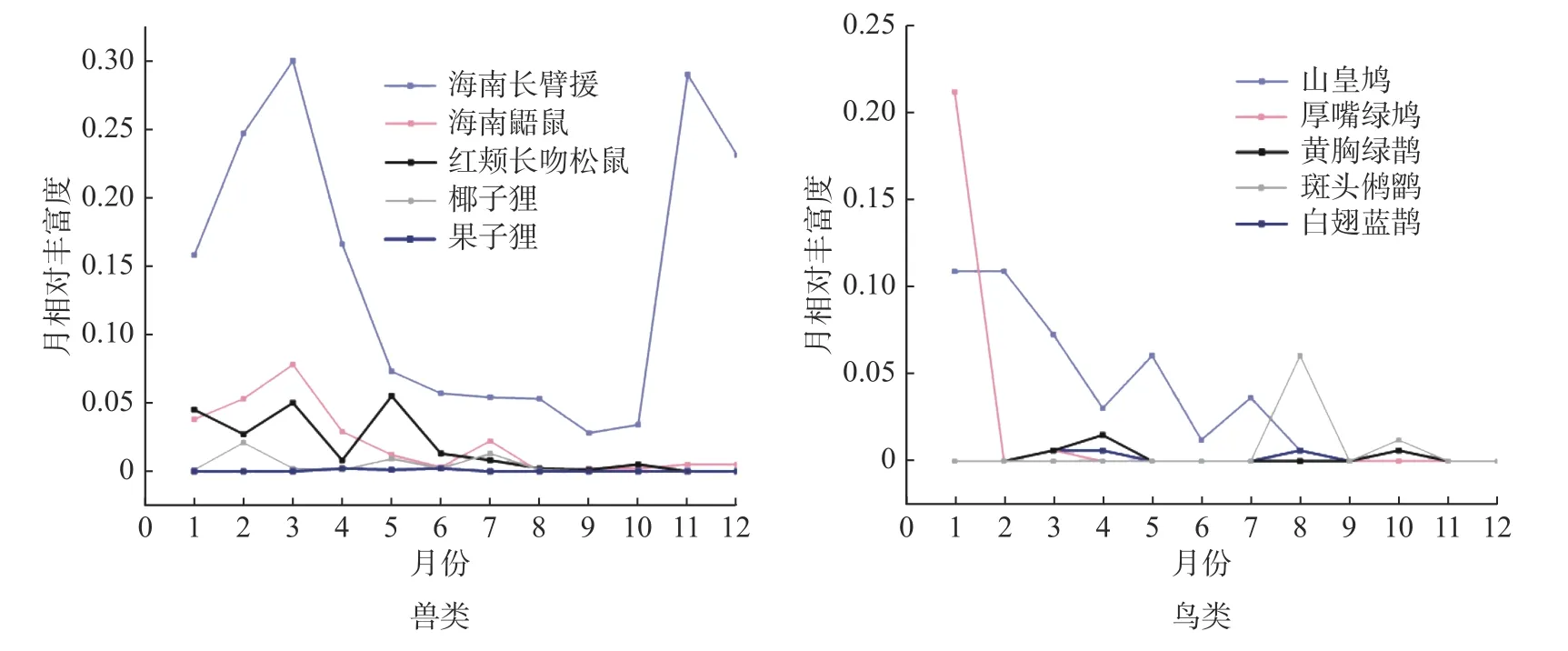
图2 兽类和鸟类的年活动格局
2.3 物种海拔分布格局不同海拔梯度下鸟类和兽类物种数均不存在显著差异(鸟类:χ2= 2.085,df = 2,P=0.339;兽类:χ2= 5.53, df = 2,P= 0.061);鸟兽物种总数则存在显著差异(鸟兽:χ2= 6.399,df = 2,P= 0.038),主要表现为,海拔1 100 m 以上的鸟兽类物种总数低于其他海拔梯度(图3)。比较不同海拔梯度的物种多样性指数、均匀度指数以及优势度指数,只发现鸟类的优势度指数存在显著性差异性(鸟类:χ2= 18.585, df = 2,P= 0.000 1),主要表现为,鸟类优势度指数在海拔1 100 m 以上明显低于其他海拔梯度;兽类和鸟兽的多样性指数和均匀度指数随着海拔的升高呈单调递减;兽类中海南鼯鼠和红颊长吻松鼠是优势种,优势度指数分别是0.13 和0.10;鸟类中山皇鸠和厚嘴绿鸠是优势物种,优势度指数分别是0.40 和0.20(图3)。
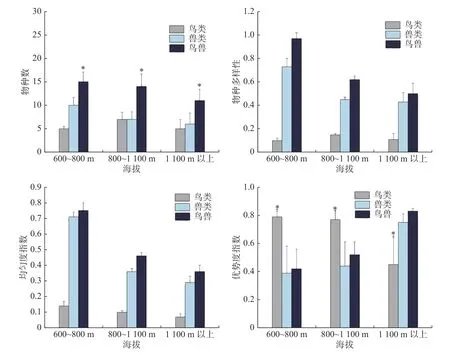
图3 不同海拔梯度下鸟类和兽类的物种数和多样性分布差异
3 讨 论
3.1 海南长臂猿及同域物种的多样性密切相关的同域物种会采用导致生态位分离或资源划分的过程,从而使物种共存成为可能[35-36]。本研究监测到与海南长臂猿同域分布的兽类包括巨松鼠、赤腹松鼠、海南鼯鼠,果子狸和椰子狸等,尽管这些兽类与海南长臂猿的食物生态位存在重叠,但在活动模式和空间利用的差异使它们可以共存[37-38]。在活动模式方面,海南长臂猿是昼型性灵长类,椰子狸、果子狸和海南鼯鼠则是典型的夜行性动物[39],这些物种时间生态位的高度分离有利于物种共存[35]。在空间利用方面,海南长臂猿是高度树栖动物,几乎不会下到地面寻找食物,而其他物种均有地面活动现象,如赤腹松鼠会在果实资源丰富的季节下到地面层搜寻食物[40]。此外,研究区域还发现另外一种灵长类猕猴,虽然该物种与海南长臂猿的食物、时间生态位高度重叠,但海南长臂猿出现位点并未发现猕猴,海南长臂猿在与猕猴的生态位竞争中可能处于优势。海南长臂猿缺少自然天敌,但少数大型猛禽,如蛇雕(Spilornis cheela)等可能会攻击长臂猿幼崽,本研究共记录到2 种猛禽,即鹰雕和草鸮(Tyto longimembris),红外相机监测和人为跟踪均未发现对海南长臂猿幼崽的伤害行为。
3.2 海拔分布格局及年活动格局的差异物种丰富度的海拔分布格局可分为4 种常见的格局:递减型、低高原型、低高原中峰型和中峰型[41],本研究兽类物种数虽然在海拔梯度间不存在显著性差异,但整体上表现为随着海拔的升高呈单调递减趋势,随着海拔的升高,温度、植被类型、食物资源等会呈现出一定的垂直分布和变化,进而对动物分布产生重要影响[42]。该研究区域海拔600~800 m、800~1 100 m、1 100 m 以上分别对应的是热带沟谷雨林、热带山地雨林和山地苔藓林。在山地苔藓林区域树木稀疏且缺少高大乔木,会引发食物资源短缺,物种间竞争变大,亦或不能为某些树栖动物提供隐蔽的栖息场所,如海南长臂猿会选择胸径更大,高度更高、冠幅更大、枝下更高的乔木夜宿[43]。鸟类的物种数指数先升高后减少,是典型的中峰模式,随海拔上升气温下降,导致鸟类在海拔1 100 m 以上的分布种数、分布数量逐渐下降,这可能是高海拔区域鸟类物种多样性指数较低的原因[33];分布区内低海拔区域较强的人为干扰,可能影响了鸟类的分布[34]。
海南长臂猿以及其他兽类年活动格局表明,海南长臂猿和海南鼯鼠的月活动强度有季节性变化,旱季月活动强度较高,雨季较低,这与海南长臂猿分布区的植被类型和气候有关。在雨季,霸王岭片区雨水充沛,大部分植物生长茂盛且处于结果期,故食物资源丰富且分布均匀;旱季降雨量偏低,植物果实减少引起海南长臂猿和海南鼯鼠的食物短缺。海南长臂猿等因移动需要付出一定的能量,可能也需要增加对叶芽类取食的策略或增加食物的摄入来弥补所消耗的能量[24],活动强度往往会因食物资源的时空变化而存在季节性差异[44]。雨季结束,旱季到来食物资源减少时,布设在果树附近的红外相机拍摄到的视频显示海南长臂猿和海南鼯鼠取果、采食画面的滞留时间变长,也表明旱季的月活动强度比雨季高。其他兽类在雨季和旱季活动强度相对稳定,与食性有关,如饮食灵活的猕猴可以通过改变其食料偏好来避免与其他类群的种间竞争[45]。鸟类的生殖期具有明显的季节性,生活在我国的鸟类一般在3~8 月份生殖[46],研究区域的鸟类活动规律与此相符,活动峰值主要集中在1— 4 月份和7—8 月份。在繁殖期间,雌雄个体相遇的概率大幅提升,从而提高了月丰富度指数。然而,在1—2 月份山皇鸠和厚嘴绿鸠的月丰富度指数高,这可能与该地区的温度较高有关。
3.3 树栖红外相机的监测优势传统的调查方法倾向于关注昼行性的野生动物,红外相机技术可以监测到动物夜间活动,从而实现对目标物种的24 h 适时监测[47]。在监测活跃的、体型较大的树栖物种,如海南长臂猿等也特别高效,尽管该物种实际数量很少,但拍摄到最多独立有效照片,而且发现动物不会回避红外相机,在自然条件下各种行为可以详细地、近距离地记录下来[2]。此外,虽然树栖红外相机高度暴露在雨水和阳光下,但此次林冠层红外相机的故障率17%低于其他地方报道的热带雨林红外相机的故障率70%[48]。本研究结果表明,在树冠层中使用红外相机调查树栖哺乳动物效果与陆地相似,是调查林冠内隐蔽、夜间活动和活跃动物群落的有效方法。国内应用树栖红外相机调查林冠层野生动物的研究极少,可以在未来的调查工作中尝试定义和标准化安装规范,如树栖红外相机安装的高度和树种的选择以及考虑到热带雨林较高的生物多样性,在1 个布设位点安装多个红外相机等。
致谢:海南热带雨林国家公园霸王岭片区对研究予以大力支持,护林员张志诚、李文永、谢赠南等对野外工作的顺利开展提供了不可或缺的帮助;陈少莲和杨川在鸟类鉴别方面给予许多帮助,谨此致谢。
猜你喜欢
小哥白尼(野生动物)(2022年7期)2022-09-20
Chinese Physics B(2021年9期)2021-09-28
云南画报(2021年7期)2021-08-06
杂文月刊(选刊版)(2021年5期)2021-06-02
环保科技(2020年2期)2020-05-21
浙江农林大学学报(2019年6期)2019-11-13
安徽农学通报(2018年8期)2018-05-30
四川林业科技(2016年1期)2016-04-11
小小说月刊·下半月(2015年11期)2015-05-14
小学阅读指南·低年级版(2009年9期)2009-10-27
