
远志叶绿体基因组序列特征与系统发育分析
2023-06-08 12:21王星璐李慧娟李彦青秦雪梅张福生
中草药 2023年11期
王星璐,李慧娟,赵 伟,李彦青,秦雪梅,张福生*
1. 山西大学 中医药现代研究中心,山西 太原 030006
2. 山西中医药大学基础医学院,山西 晋中 030619
在植物中核基因组的复杂性使得低拷贝基因的筛选比较困难,目前仅局限在基因组相对较小的模式物种或具有重要经济生态价值的物种中,整体应用范围较为有限。植物线粒体基因组具有在植物类群中变异很大、存在基因组之间水平转移的外源基因和进化速率较慢等特性,使其在系统发育研究中的应用受到了限制[1]。叶绿体基因组作为第2 大基因组,属于相对独立于核基因组之外的基因组[2],其基因组完整且相对独立。大多数被子植物的叶绿体含有大量遗传信息,同时具有相对分子质量小、结构简单,进化速率中等、突变率较低、遗传稳定、成本低及开发难度低、广泛分布着微卫星序列和线粒体 DNA 等特点[3-4],在一定程度上弥补了线粒体和核基因组的部分缺点与不足。
叶绿体起源于古细菌入侵植物细胞[5],是一种拥有自身遗传物质的多功能细胞器[6],为质体中的一种类型。叶绿体基因组的结构大部分为共价闭合的双链状环形分子,也有少部分是线性分子或者多聚体。对于高等植物而言,叶绿体存在于细胞质的基质中,其形状呈现为绿色椭圆球型或者是圆球形[7]。典型的环式双链叶绿体基因组结构,包括4 个部分,即大单拷贝区(large single copy,LSC),小单拷贝区(small single copy,SSC)和2 个反向重复区(inverted repeats,IRs);其中,2 个反向重复区把基因组分隔为大单拷贝区和小单拷贝区[8]。LSC 区长81~90 kb,SSC 区介于18~20 kb;2 个IR 区序列基本一致,大小介于20~30 kb,是叶绿体基因组进化过程中延展或缩小的区域[9]。大多数陆生植物的叶绿体基因组中有110~130 个基因[10]。近年来,由于叶绿体基因组大小、结构和基因种类一般较为保守,叶绿体全基因组的物种鉴定及系统进化研究成为植物系统分类学的一个新趋势,为研究药用植物系统进化及进行物种鉴定提供可靠工具[11]。目前,叶绿体基因组技术已经广泛应用于药用植物的研究中[12-14]。
远志植物为双子叶植物芸香目远志科一种一年生或多年生草本、灌木或小乔木,约500 种,广布于全世界,我国有42 种8 变种,广布于全国各地,而以西南和华南地区最盛[15]。远志属远志PolygalatenuifoliaWilld.最早记载于《华氏中藏经》[16],药用价值广泛,历史悠久,具有较大的药用研究和开发利用价值。近年来由于过度采挖远志药材,导致野生药材资源严重不足,远志已被收入《国家重点保护野生药材物种名录》,保护级别为Ⅲ级。目前关于远志的研究主要集中于其化学成分与药理作用等方面。研究发现,远志具有改善学习记忆、抗氧化、抗抑郁、抗衰老、镇静催眠、益智和抗肿瘤以及祛痰镇咳、影响药物代谢以及抗炎等[17-24]作用。分子生物方面研究主要集中在远志随机扩增多态性DNA 分析[25]、遗传多样性[26]、远志谱系地理学[27-28]、序列鉴定[29-30]等方面;对叶绿体基因组的研究报道较少,虽有对西南远志P.crotalarioidesBuch.-Ham. ex DC.[31]和远志P.tenuifoliaWilld.[32]叶绿体基因组进行测序、拼装和注释的报道,但是报道的远志[32]仅限于描述了序列全长和系统发育位置等一些基本信息,未对其展开详细的分析描述。
目前,远志属物种的叶绿体基因组研究十分缺乏,其相关的亲缘关系值得深入研究,NCBI 数据库中有远志P.tenuifoliaWilld.(NC_050829.1)[32]完整叶绿体基因组数据,但是仅记载了叶绿体基因组总长(165 423 bp)等基础信息,也未进行深入挖掘研究,相关信息依旧不清楚。本研究利用生物信息学相关软件,分析其叶绿体基因组的构成,并对其进行基因组装和注释,分析序列特征,密码子偏好性以及系统发育,阐明远志叶绿体基因组结构特征以及其物种之间的亲缘关系。远志叶绿体基因组的研究是对远志药材种质资源遗传多样性的进一步了解,有利于远志药材优良品种的选育和种质资源的评价及其有效保护与合理利用。
1 材料与方法
1.1 样品收集
从美国国立生物技术信息中心(National Center of Biotechnology Information,NCBI)数据库(https://www.ncbi.nlm.nih.gov/)检索远志科远志属物种的叶绿体全基因组序列信息。检索到远志科远志属8 个物种以及齿果草属1 个叶绿体全基因组序列信息,远志科亲缘关系较近的芸香目苦木科鸦胆子Bruceajavanica(L.) Merr.和芸香科九里香MurrayaexoticaL. Mant.完整叶绿体全基因组序列。下载检索到的物种的叶绿体基因组名称、基因组登录号(表1)。
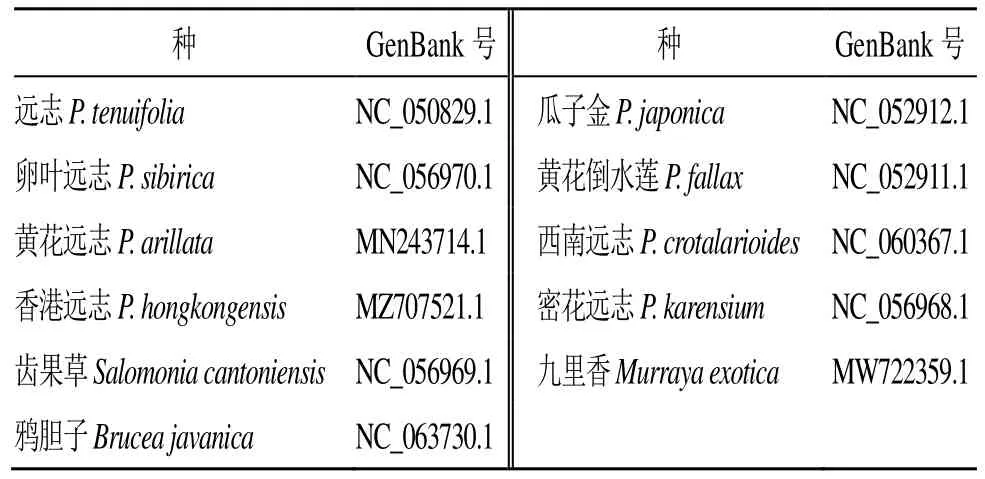
表1 远志科、苦木科和芸香科叶绿体GenBank 登录号Table 1 GenBank accession number of chloroplast in Polygalaceae, Simaroubaceae and Rutaceae
1.2 叶绿体基因组注释、图谱绘制以及基本特征分析
远志属远志的叶绿体全基因组序列(登录号为NC_050829.1 ) 通 过 GeSeq :( https://chlorobox.mpimp-golm.mpg.de/geseq.htm)[33]和Plastid Genome Annotator(PGA)[34]软件进行基因注释,将结果对比矫正,去除错误及冗余注释。通过 Chloroplot(https://irscope.shinyapps.io/Chloroplot/)[35]在线绘制工具绘制。通过GeSeq 初步注释和Chloroplot 画图相结合对叶绿体基因组的总长度及各个区域(LSC、SSC、IR)的长度、基因组成(蛋白编码基因、tRNA基因、rRNA 基因)、碱基组成、GC(AT)含量进行统计和比较分析,解析远志叶绿体基因组序列的基本特征。
1.3 叶绿体全基因组重复序列检测
利用Perl 语言通过MISA 软件(http://pgrc.ipk-gatersleben.de/misa/misa.html)[36]完成简单重复序列(simple sequence repeats,SSRs)检测,参数设置为单核苷酸重复单元不少于10 个,二核苷酸重复单元不少于5 个,三核苷酸和四核苷酸重复单元不少于4 个,五核苷酸和六核苷酸重复单元不少于3 个,且2 个SSRs 之间的最小距离为100 bp[37]。
叶绿体全基因组中的散在重复序列(dispersed repeats)利用REPute 软件(https://bibiserv.cebitec.uni-bielefeld.de/reputer)[38]进行检测,正向重复(forward repeats,F)、反向重复(reverse repeats,R)、互补重复(complement repeats,C)、回文重复(palindromic repeats,P)。参数设置最小重复长度(minimal repeat size)设置为30,hamming 距离(hamming distance)为 3,最大计算重复次数(maximum computed P repeats)5000[39]。串联重复序列(tandem repeats)利用Tandem repeats finder软件(https://tandem.bu.edu/trf/trf.html)进行检测。参数设置选择默认值[40]。
1.4 密码子使用分析
采用CodonW(http://codonw.sourceforge.net)软件[41]分析密码子使用情况。对远志的叶绿体基因组同义密码子相对使用频次( relative synonymous codon usage,RSCU)进行分析和统计。当RSCU>1 时,表明该密码子使用频率较高;RSCU=1 时,说明该密码子无偏好性;RSCU<1时,表明该密码子使用频率较低[42]。
1.5 基因组比较分析
采用Geneious[43]软件统计远志属8 个物种的叶绿体基因组序列的4 个边界(SSC、LSC、IRa和IRb 区域)长度和基因数目类型、GC 含量等信息,并用EXCEL 计算各自的GC 值。使用IRscope(https://irscope.shinyapps.io/irapp/)[44]可视化工具,比较远志属叶绿体基因组4 个区域边界的差异。并采用mVISTA 对其进行全基因组比对分析。
1.6 基于叶绿体基因组序列的系统进化分析
为了确定远志的系统发育位置,下载NCBI 数据库中收录的远志科上述所有9 种植物的叶绿体全基因组序列。与本研究远志叶绿体全基因组序列共同构建序列矩阵,同时选择远志属的近缘类鸦胆子和九里香作为外类群。利用MAFFT 7 软件(https:// mafft.cbrc.jp/alignment/software/)[45]完成序列比对。利用IQtree2.0.5(http://www.iqtree.org/)[46]以最大似然法(maximum likelihood,ML)进行系统发育关系重建分析,核苷酸替换模型选择GTR+G,其他参数选择默认设置,进行1000 次自展重复分析,计算系统发育树各节点的靴带支持率(bootstrap values,BS)。并利用MEGA X 软件,使用邻接法(neighbor-joining,NJ)构建系统进化树,并进行系统发育树的比较。
2 结果与分析
2.1 远志叶绿体基因组的基本特征
远志的叶绿体基因组与典型的陆地植物叶绿体基因组基本特征一样,均为典型的四分体结构,远志属远志叶绿体基因组呈双链环状四分体结构(图1),大小为165 423 bp,包括大单拷贝区、小单拷贝区以及2 个反向重复区,各区长度分别为83 699、8044、36 840 bp。远志叶绿体基因组的总GC 量为36.7%,IR 区的GC 量为39.7%,高于LSC 区(34.7%)及SSC 区(29.4%)。整体而言,IR 区的保守性高于LSC 区和SSC 区,这可能是IR 区的GC 量高于LSC 区和SSC 区而造成的。计算各分区的碱基组成(表2)。
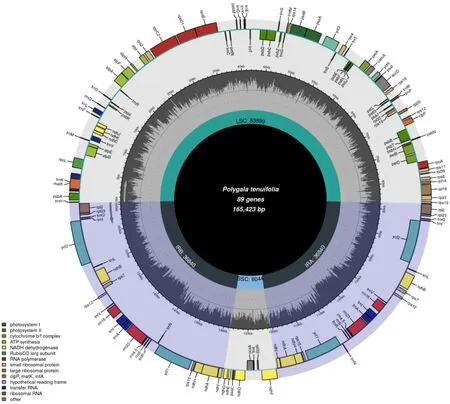
图1 远志叶绿体基因组图谱Fig. 1 Chloroplast genome map of P. tenuifolia
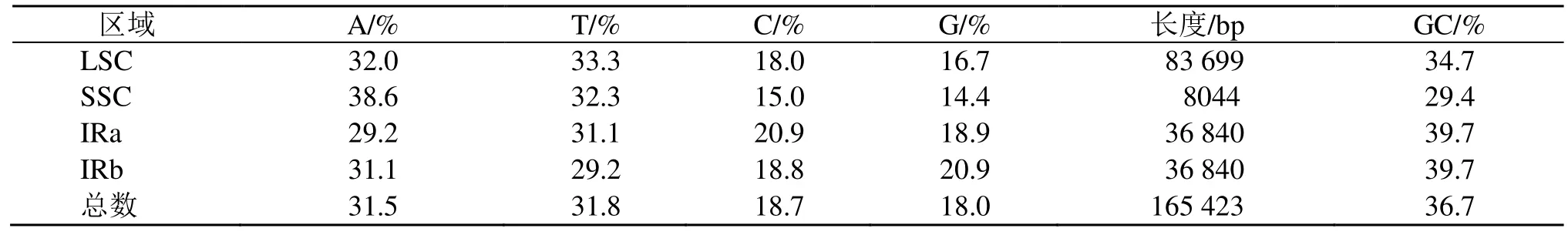
表2 远志叶绿体基因组碱基组成Table 2 Base composition of chloroplast genome of P. tenuifolia
2.2 远志叶绿体基因组的基因注释及归类分析
远志叶绿体基因组共注释得到135 个基因,包括8 个rRNA 基因、38 个tRNA 基因和89 个蛋白编码基因。根据其功能可分为以下4 个大类(表3)。第1 类:48 个与光合作用相关的基因,包括5 个光合系统I基因、15 个光合系统II 基因、6 个细胞色素b/f复合体基因、6 个ATP 合酶基因、1 个二磷酸核糖体羧化酶、15 个NADH 氧化还原酶基因。第2 类:除rRNA 基因和tRNA 基因外,有30 个与自我复制相关的基因,包括10 个核糖体蛋白大亚基基因、16 个核糖体蛋白小亚基基因和4 个RNA 聚合酶亚基基因。第3 类:6 个其他编码蛋白质的基因。第4 类:6 个功能未知的基因。在这些基因中,有12 个蛋白编码基因(rpl2、rpl23、ycf2、rps12、rps7、rps15、ycf1、ndhA、ndhB、ndhH、ndhI、ndhN)、7 个tRNA 编码基因(trnQ、trnI、trnL、trnV、TrnA、trnN、trnR)和4 个rRNA 编码基因(rrn4.5、rrn5、rrn16、rrn23)位于IR 区。

表3 远志叶绿体基因组基因功能注释与分类Table 3 Gene functional annotation and classification of P. tenuifolia chloroplast genome
2.3 重复序列统计分析
叶绿体基因组SSR 分析在远志叶绿体基因组中搜索到161 个SSR 位点。其中,29 个为复合型SSRs;单核苷酸重复Motif 位点最多为113 个,二核苷酸、三核苷酸、四核苷酸、五核苷酸和六核苷酸重复基序分别有30、3、9、4、2 个(表4)。SSR的类型以A/T 为主,无含有G/C 组成的位点,占总重复类型的70.18%;其次为AT/AT,共有25 个(15.52%),AG/CT(3.11%),其余总数占总重复类型的11.18%。A/T 和AT/AT 占简单重复序列位点重复单元总数量的85.71%,因此远志叶绿体基因组富含AT。从分布区段上看,34 个位点位于IR区段,115 个位于LSC 区段,12 个位于SSC 区段,说明远志叶绿体基因组SSRs 分布的不均匀性。

表4 远志叶绿体基因组SSRs 位点类型及数量Table 4 Type and number of SSRs of P. tenuifolia chloroplast genome
使用REPuter 软件在远志叶绿体基因组中共检测到223 个散在重复序列,包括正向重复序列(F型)、反向重复(R 型)、回文重复序列(P 型)和互补重复(C 型)4 种类型。其中,正向重复序列(F 型)89 个,反向重复(R 型)13 个,回文重复序列(P 型)109 个和互补重复(C 型)12 个。长度为30~39 bp 的重复序列中包括61 个正向重复,12 个反向重复,72 个回文重复和12 个互补重复。其分布数量相对较多(图2)。发现90 条长度范围在2~54 bp 的串联重复序列。串联重复序列,4个区域均有分布,其中,LSC 区35 条,重复次数1.9~26.8 次,TRB 区24 条,重复次数由1.9~12.2次,SSC 区1 条,重复次数由2 次,TRA 区30 条,重复次数由1.9~8.1 次。
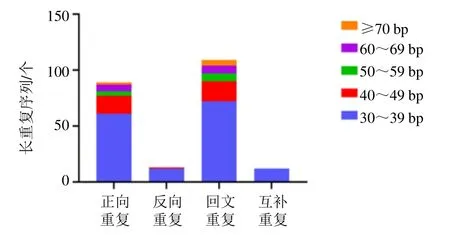
图2 远志叶绿体基因组散在重复序列类型与数量Fig. 2 Type and number of long repeats in P. tenuifolia chloroplast genome
2.4 叶绿体基因组密码子偏好性分析
用CodonW 软件分析远志叶绿体基因组蛋白编码序列密码子的使用偏好性(图3),共预测了55 141 个密码子,其中编码亮氨酸(Leu)的密码子数量最多(5629 个,10.21%),编码色氨酸(Trp)的密码子数量最少(705 个,1.28%)。在远志叶绿体基因组中有20 种氨基酸,除甲硫氨酸(Met)和色氨酸(Trp)使用1 个密码子AUG 和UGG 外,其余氨基酸均有 2~6 个同义密码子。亮氨酸(Leu)、精氨酸(Arg)、丝氨酸(Ser)使用6 个同义密码子,使用频率最高的密码子分别为UUA、UCU、AGA;丙氨酸(Ala)、甘氨酸(Gly)、脯氨酸(Pro)、苏氨酸(Thr)、缬氨酸(Val)分别使用4 个同义密码子,使用频率最高的密码子分别为GGA、CCA、GUU、CCU、ACU;异亮氨酸(lle)使用3 个同义密码子,使用频率最高的密码子是AUU;半胱氨酸(Cys)、天冬氨酸(Asp)、谷氨酸(Glu)、苯丙氨酸(Phe)、组氨酸(His)、赖氨酸(Lys)、天冬酰胺(Asn)、谷氨酰胺(Gln)、酪氨酸(Tyr)均使用2 个同义密码子,但使用频率并不相同,这9 个氨基酸使用频率较高的密码子分别为UUU、AAA、AAU、UAU、UUC、GAA、GAU、AAG、CAA。结果表明,远志叶绿体基因偏好使用含有A、U 碱基的密码子,密码子第3 位也偏好以A 和U 结尾。
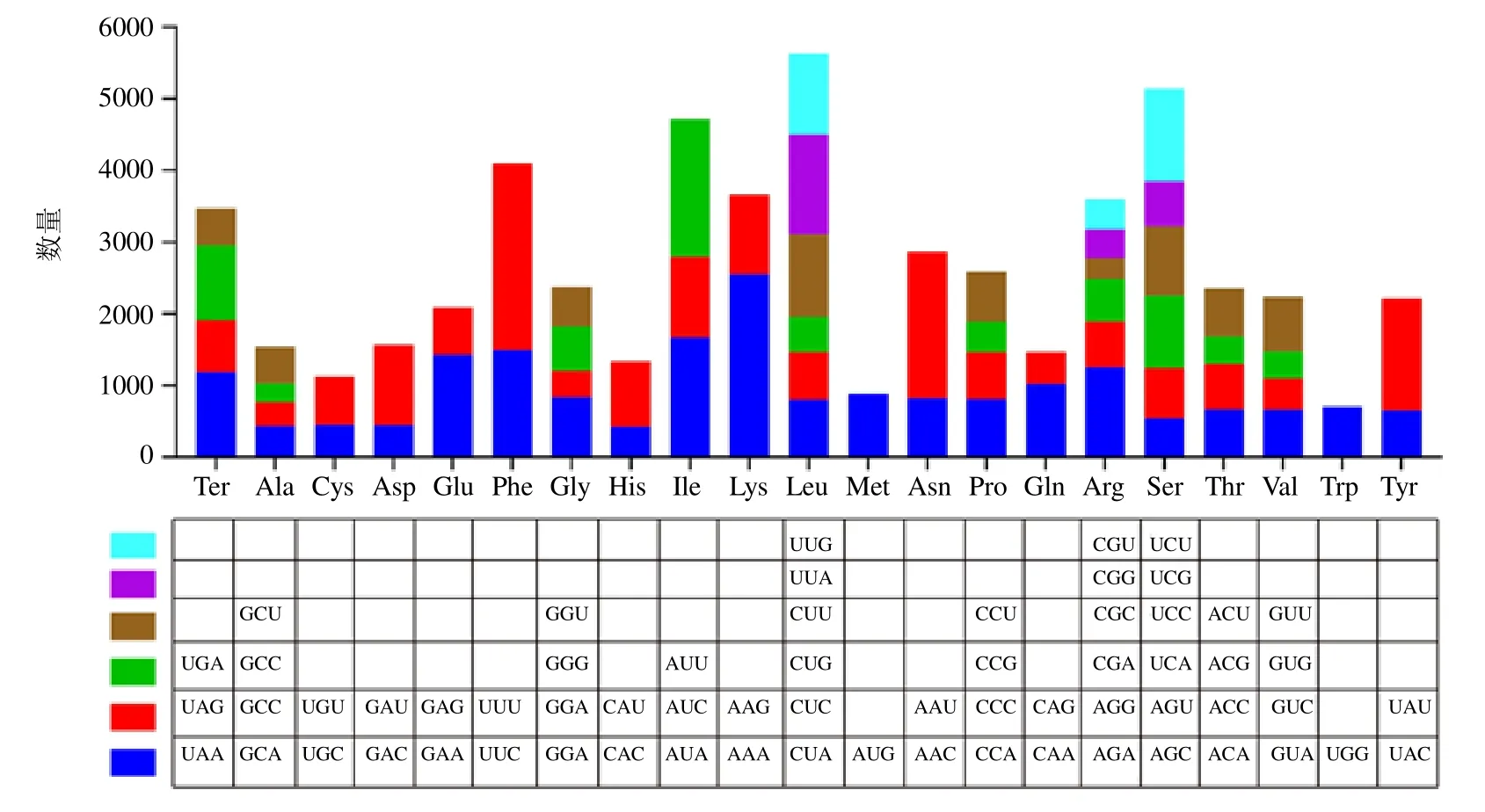
图3 远志叶绿体基因组的密码子使用频率Fig. 3 Codon usage frequency in chloroplast genome of P. tenuifolia
相对同义密码子使用度(RSCU)分析表明,在所有被编码的密码子中,RSCU>1 的密码子共有35 个(AGA、UCU、UUA 等),其中27 个密码子以A/U 碱基结尾,以A 结尾的占51.85%,以U 结尾的占48.15%,故这些密码子的结尾具有A/U 偏好性。RSCU<1 的密码子共有29 个(CGC、AAC、GAC 等),以G/C 结尾的共有23 个,且以C 结尾占52.13%和以G 结尾的占47.83%(表5)。
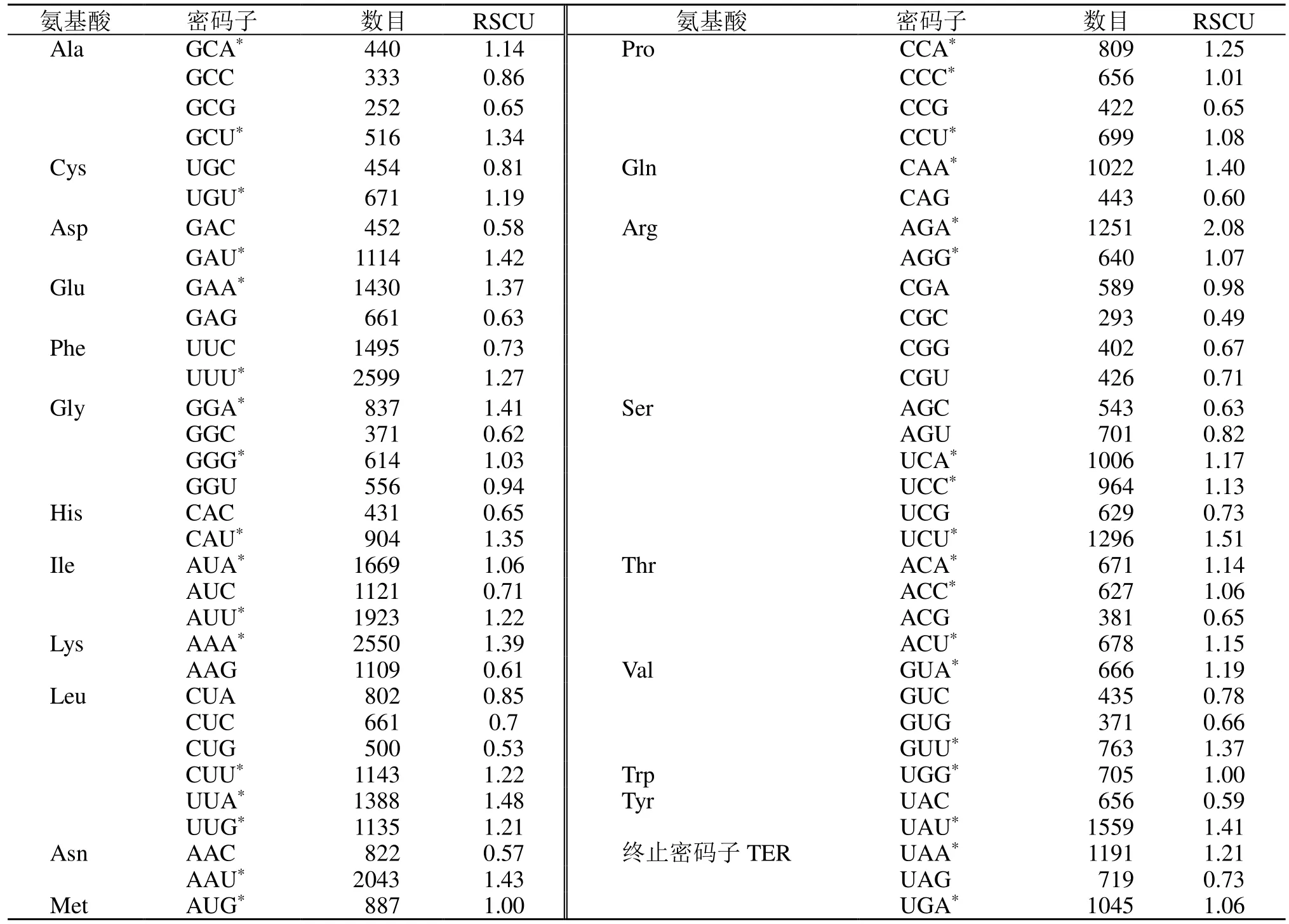
表5 远志绿体基因组蛋白编码序列RSCU 分析Table 5 RSCU of protein coding region in chloroplast genome of P. tenuifolia
2.5 基因组特征及IR 边界比较分析
从NCBI 下载了远志、瓜子金、卵叶远志、黄花倒水莲、黄花远志、西南远志、香港远志和密花远志8种远志属植物的叶绿体基因组,它们的全长分别为165 423、165 439、165 397、164 687、164 947、164 268、165 296、171 893 bp。黄花倒水莲叶绿体基因组的GC 值最大,为36.9%,其余GC 含量均在36.7%~36.9%(表6)。瓜子金的LSC 最长,为83 722 bp、密花远志的SSC 最长为8409 bp。
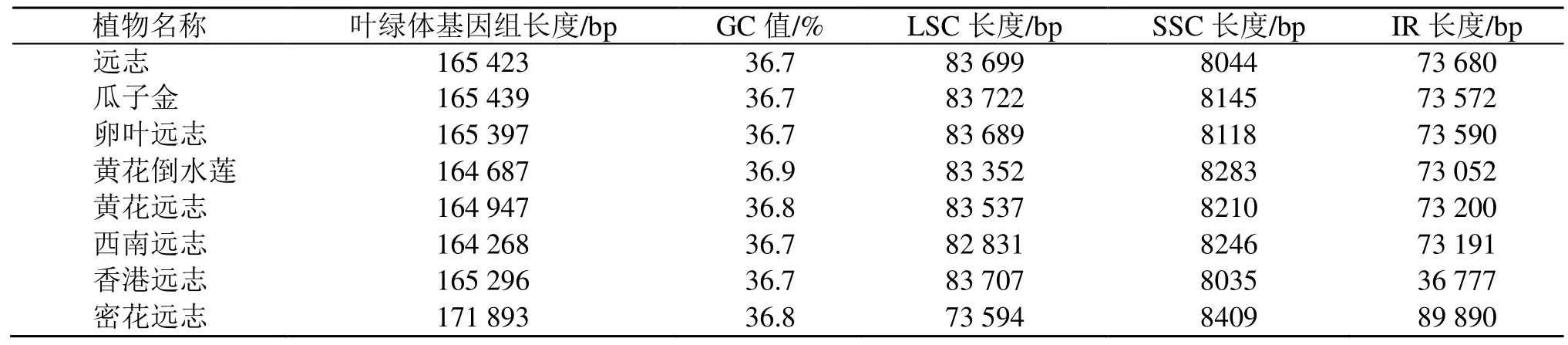
表6 8 种远志属植物叶绿体基因组的特征Table 6 Chloroplast genome characteristics of eight Polygala plants
在远志属8 种植物的叶绿体基因组边界区域中,远志叶绿体基因组共存在4 个边界,远志与其他7种植物的LSC/IRb、IRb/SSC、SSC/IRa 和IRa/LSC边界及基因分布(图4)。除密花远志和西南远志外,JLB(LSC/IRb)边界两侧分布均有rpsl2和rps19基因,密花远志边界位于petB基因中,是其独有的基因,该基因的保守长度为1390 bp,西南远志JLB 边界有rpl23基因存在,距离边界184 bp。其中远志中的rps19基因距离JLB(LSC/IRb)边界长度为1 bp,其位于LSC 区域内,rps12位于IRb 区域内。
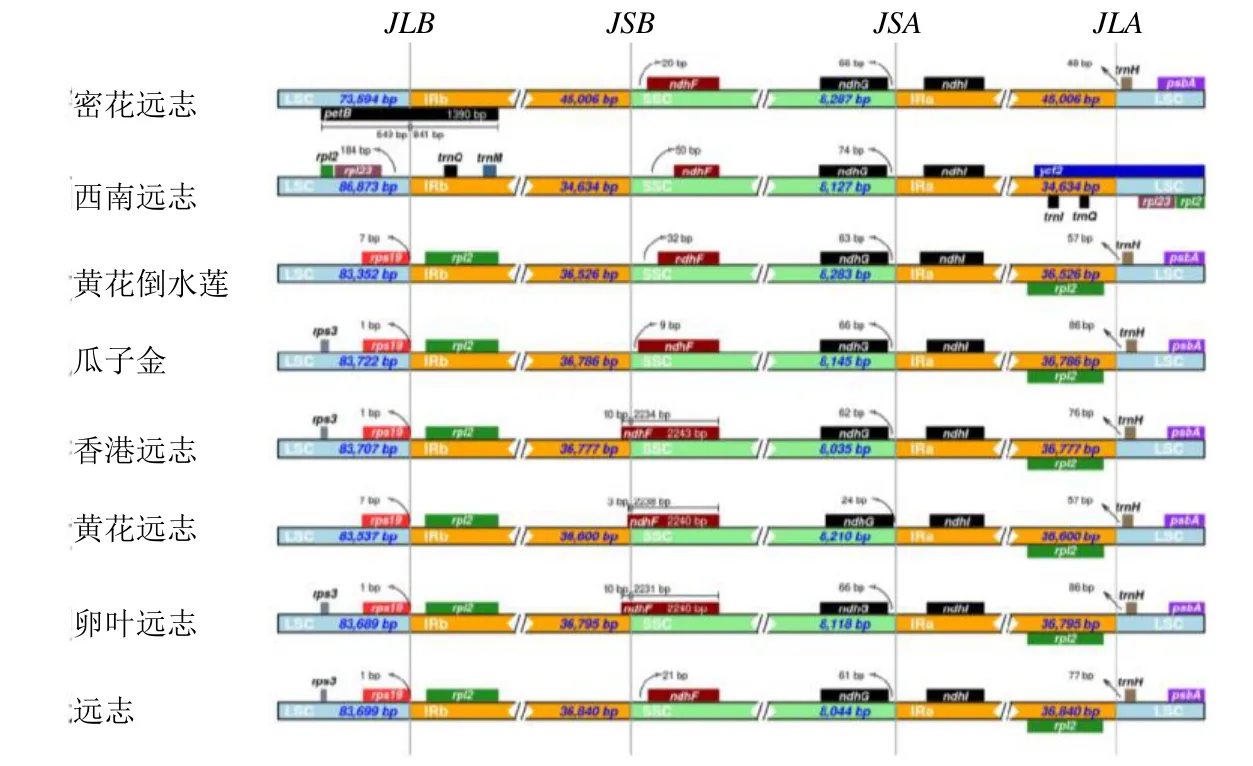
图4 远志属8 个物种叶绿体基因组的IR 与SC 边界比较Fig. 4 Boubdary comparison of IR and SC region of chloroplast genomes in nine species of Polygala
ndhF基因位于JSB(IRb/SSC)的边界,8 种植物在该位置与基因存在间隙或重叠存在。其中该基因在远志中距SSC 区仅21 bp,其中香港远志和卵叶远志ndhF扩张最大。香港远志、黄花远志和卵叶远志的JSB(IRb/SSC)边界均位于ndhF,其余远志属物种均在SSC 区域,该基因相对保守长度均为2240 bp。另一个ndhG基因位于JSA(SSC/IR)交界处,均位于SSC 区域中。除了黄花远志距离边界24 bp,其余距离边界60~74 bp。
ndhI基因位于JSB(IRb/SSC)的边界,均位于IRa 区域中。西南远志JLA(IRa/LSC)边界分布有ycf2基因,密花远志分布有trnH基因。而其他6 种植物在该处均为trnH和rpl2基因。其中,trnH基因均分布在LSC 区域,rpl2基因均分布在IRa 区域。IRA-LSC 边界均与trnH基因存在着48~86 bp 的间隙。总体来说,远志属的西南远志和密花远志叶绿体基因组和边界基因与其余6 个叶绿体基因组相互之间具有明显的差异。
2.6 基于叶绿体基因组的远志系统发育分析
利用最大似然法构建系统发育分析结果显示,11 种植物被分为2 大类,即远志科和外类群(苦木科和芸香科),见图5。包括远志在内的远志属8 个物种和一个齿果草属以100%的支持率构成一个单系分支与芸香科和苦木科区分开来。同时,这9 个物种可以进一步划分为2 个次级单系分支,其中,同来自远志属的黄花远志、黄花倒水莲、密花远志和齿果草属齿果草4 个物种构成一个单系的支持率为100%,齿果草属和部分远志属聚类到一块值得进一步研究,侧面说明叶绿体基因组序列也并不能完全解决远志属内部网状进化等问题,仍需和其他基因组学联合分析。结合IR 边界分析可知远志、瓜子金、卵叶远志和香港远志的亲缘关系最近。本研究的远志叶绿体基因组序列可为远志科后续开展遗传多样性研究提供重要信息。
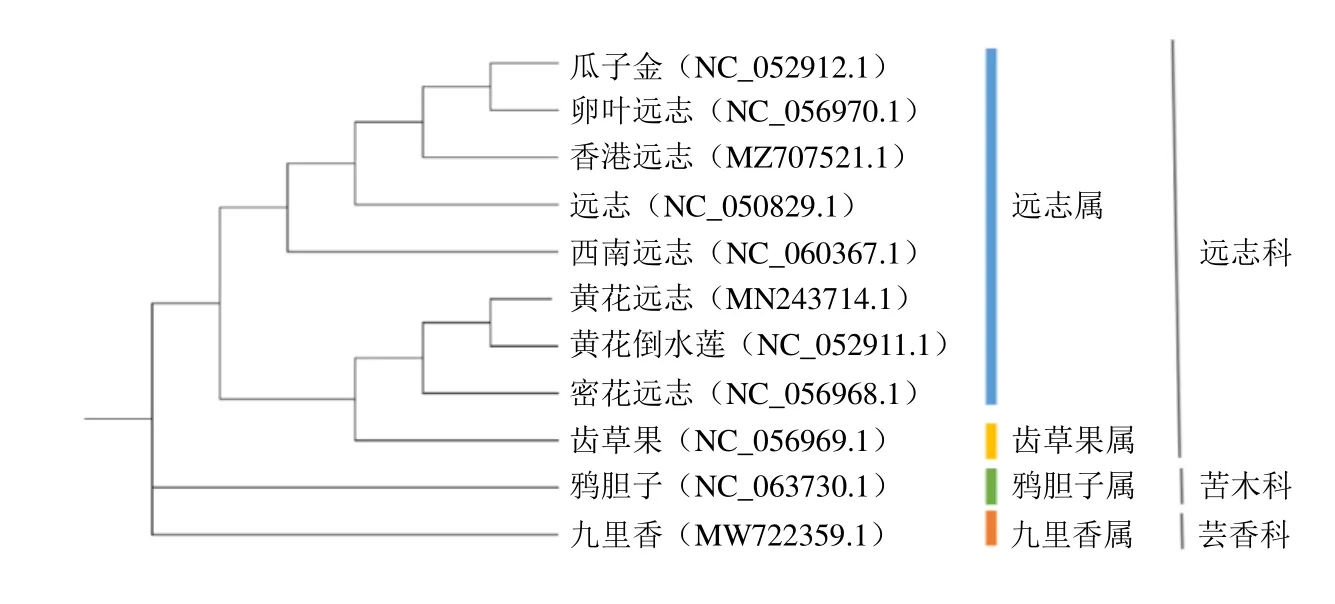
图5 基于叶绿体全基因组序列构建ML 系统进化树Fig. 5 Phylogenetic analysis based on chloroplast genome sequences by maximum likelihood (ML) tree
邻接法的结果与最大似然法构树除了齿果草和密花远志位置互换外,其余结果一致,见图6。远志科仍分为2 大类同来自远志属的远志和卵叶远志、瓜子金和香港远志4 个物种分为一类,黄花倒水莲、黄花远志、密花远志和齿果草属齿果草聚到一类。仍然可以说明远志和卵叶远志、瓜子金和香港远志关系密切。
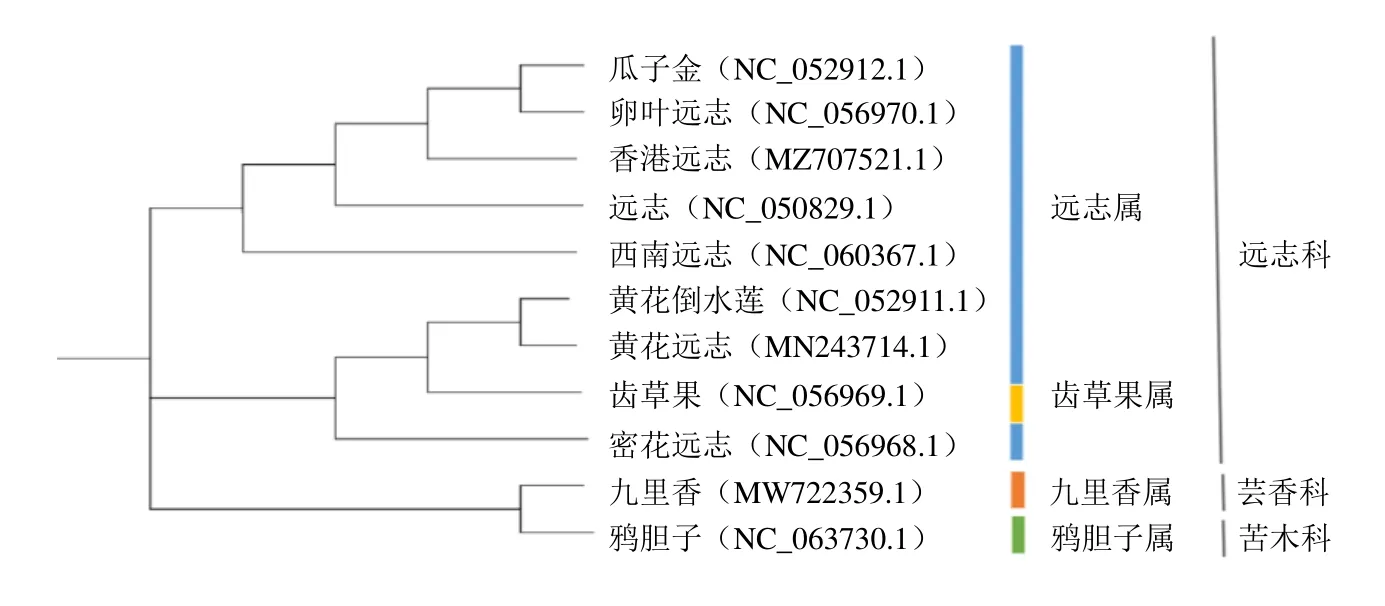
图6 基于叶绿体全基因组序列构建NJ 系统进化树Fig. 6 Phylogenetic analysis based on chloroplast genome sequences by neighbor-joining (NJ) tree
3 讨论
《中国药典》2020 年版中将远志科植物远志P.tenuifoliaL.的干燥根作为远志的主流基原药材[47]进行商品流通,其药用价值显著。近年来,面临远志的野生资源状况不容乐观,植物资源日益贫乏的危险。本实验基于叶绿体基因组技术进行序列分析、密码子偏好分析、远志属基因组比较分析以及远志属植物系统发育关系研究为远志在野生资源保护、分子育种及远志属药用植物的物种鉴定等领域提供了宝贵的基因资源。
本实验结果发现,远志叶绿体基因组长为165 423 bp,GC 含量为36.7%,共编码135 个基因,这与先前报道的远志属其他植物西南远志[31]的叶绿体基因组结构特征类似,表明远志的叶绿体基因组结构相对保守。与韩国学者Lee 等[32]发表的文章相比较,本研究将系统发育树聚焦到远志科植物上,且增加对其重复序列,密码子偏好性,远志属基因组比较,IR 边界内容的分析,补充和完善前者文章分析量少的缺点,从而对其叶绿体基因组有了更加全面的了解。
SSR 在植物叶绿体基因组中十分常见,其类型、数目及分布都因植物不同而异,被广泛应用于植物群体遗传多样性和系统发育研究以及分子标记研究等内容[48-49]。本研究通过分析远志叶绿体基因组,共检测到161 个SSR 位点,其主要位于LSC 区域;并且单核苷酸A/T 碱基在SSR 位点中出现频率最高,调研文献推测原因是在大部分植物中A/T 含量均高于G/C 含量,即A/T 类型的SSR 在植物中最多[37,39,42]。本研究通过对远志叶绿体基因组中SSR 的数量、组成进行分析,为后续进一步研究分子标记、群体遗传分析以及作物育种提供参考。
CUB 是基因组中重要的进化特征,RSCU 是作为密码子偏好性的指标之一[50]。研究表明,自然界存在的20 种氨基酸中,除Met 和Trp 由唯一密码子编码外,其他氨基酸均对应2~6 个同义密码子,由于同义密码子在机体内的使用频率存在差异,导致植物密码子的出现频率不同,在不同物种翻译的过程中。存在突变和自然选择等多种因素,表现出一定的偏好性[51]。本研究中密码子偏好性分析表明,亮氨酸(Leu)是远志叶绿体基因组中占比最高的氨基酸,并且RSCU>1 的27 种密码子均以A/U 结尾,这与其他高等植物相似。
通过对远志属基因组特征分析,可知黄花倒水莲叶绿体基因组的GC 值最大,为36.9%,其余GC 含量均在36.7%~36.9%。瓜子金的LSC 最长为83 722 bp、密花远志的SSC 最长为8409 bp,通过IR 边界分析远志、卵叶远志、瓜子金、香港远志、黄花远志、黄花倒水莲亲缘关系较近,其物种结构更加相似。相对保守的IR 区域的收缩与扩张现象代表着植物的进化,不同植物的叶绿体基因组大小与其密切相关。分析IR-LSC/SSC 区域的边界信息,对叶绿体基因组结构的差异、物种进化等有进一步了解[52]。通过对8 个物种的IR 边界研究发现,在密花远志叶绿体基因组的IR 长度最大,其叶绿体基因组长度8 个物种中也最大,远志属植物叶绿体基因组除了西南远志和密花远志外,基本边界变化呈现规律性。
基于远志科9 个物种的叶绿体基因组数据构建最大似然树和邻接法两种构建系统发育树,支持率均为100,其进化树的拓扑结构略有不同,与Ma 等[31]构建的进化树存在一些差异,分析其系统发育关系可知,远志、卵叶远志、瓜子金和香港远志为姊妹类群,表明四者亲缘关系最近,另外远志科与外类群其他属能够很好的区分。原因首先基于叶绿体不同数据集构建的进化树,本研究相比其数据量更为丰富些。其次叶绿体基因片段可能会丢失某些重要的信息,难以解决物种多、分类较难的大科的系统进化问题。并且基于叶绿体的不同数据集构建的进化树相比于其他方法构建的进化树支持率更高,可靠性更强,为远志的系统分类地位和种间进化关系研究奠定一定的理论基础。
本研究对远志叶绿体基因组进行研究,并分析了其叶绿体基因组结构特征,挖掘其叶绿体基因组的重复序列位点、分析其密码子偏好性和远志属叶绿体基因组IR 与SC 边界分析并利用叶绿体基因组数据构建系统发育树,有助于后续分子标记、DNA条形码技术等研究的深入进展,提供了远志叶绿体基因组信息支持。揭示远志属物种之间的系统关系,为药用远志的资源筛选、鉴定、保存及遗传多样性分析等后续研究提供了分子依据,为产业化应用奠定基础,进而为保护远志物种资源提供有力保障。
利益冲突所有作者均声明不存在利益冲突
猜你喜欢
生物学通报(2020年11期)2020-10-22
延河(下半月)(2019年7期)2019-07-30
中成药(2018年7期)2018-08-04
延河(下半月)(2017年8期)2017-09-12
广西林业科学(2016年1期)2016-03-20
茶叶通讯(2014年4期)2014-02-27
食品科学(2013年6期)2013-03-11
中国烟草学报(2012年2期)2012-04-09
河北医科大学学报(2011年1期)2011-03-25
植物营养与肥料学报(2010年2期)2010-10-24
