
干旱胁迫下野生大豆水通道蛋白GsPIP2;7功能研究
2023-06-05 06:23王艳丽俎天娇司增志孙伟明乔亚科
核农学报 2023年7期
王 宇 赵 圆 王艳丽 俎天娇 司增志 孙伟明 乔亚科 张 锴
(河北科技师范学院,河北省作物逆境生物学重点实验室(筹),河北 秦皇岛 066004)
水通道蛋白(a quaporins,AQPs)是植物中运输水分及其他小分子物质的重要通道,存在于所有真核生物和大多数原核生物中[1]。在高等植物中,AQPs 由6个高度保守的α-螺旋构成跨膜结构,通过A、B、C、D、E 5 个环相连接。水通道蛋白的B 环和E 环具有高度保守的天冬酰胺-脯氨酸-丙氨酸(Asn-Pro-Ala,NPA)独特结构。B 环和E 环各自形成半个跨膜螺旋,围绕形成狭窄的水孔,形成水通道蛋白的“水漏”结构模型[2]。
根据AQPs 在细胞内的位置可分为7 个亚族:质膜内在蛋白(plasma membrane intrinsicproteins,PIPs)、液泡膜内在蛋白(tonoplast intrinsic proteins,TIPs)、类根瘤素26(NOD26)膜蛋白(nodulin-26 like intrinsic proteins,NIPs)、膜内在小分子碱性蛋白(small basic intrinsic proteins,SIPs)、类GlpF 膜内在蛋白(GlpF-like intrnsic proteins,GIPs)、混合内在蛋白(hybrid intrinsic proteins,HIPs)和未鉴定的膜内在蛋白(X intrinsic proteins,XIPs)[3]。PIPs 存在于细胞质膜上,分为PIP1和PIP2 两个亚类;TIPs 有α、β、γ、δ、和ε 五个亚类,分别存在于不同液泡膜上[4];NIPs 位于质膜和细胞内膜上,分为NOD26和LIP2两个亚类;SIPs位于内质网上,分为SIP1 和SIP2 两个亚类,可促进水分运输[5];GIPs和HIPs 仅存在于小立碗藓(Physcomitrella patens)等几种苔藓植物中[6]。
目前,在拟南芥(Arabidopsis thaliana)中已发现35 个AQPs[7]、玉米(Zea mays)中36 个AQPs[8]、水稻(Oryza sativa)中33 个AQPs[9]、栽培大豆(Glycine max)中72个AQPs[10]、野生大豆(Glycine soja)中鉴定了62个AQPs[11]。前人研究结果说明水通道蛋白PIPs基因亚家族在植物对抗干旱、盐胁迫等非生物胁迫时具有正向调控作用[11-16]。但是也有一些不同研究结果的报道,如Wang等[17-18]研究发现过表达野生大豆GsPIP2;1和GsTIP2;1的转基因拟南芥对盐和干旱胁迫的耐受性降低。此外,干旱胁迫下,不同植物中的PIPs基因表达量也不同[19-20]。以上研究结果表明,植物AQPs的基因转录及蛋白表达调控比较复杂,AQPs在调控植株非生物胁迫中的功能尚需进一步研究。
本课题组(河北科技师范学院野生大豆资源课题组)前期筛选到耐旱性强的野生大豆材料永46,对其干旱胁迫后的基因转录组进行了研究,发现与敏感材料相比,高耐旱野生大豆材料在干旱胁迫后AQPs基因存在显著上调或下调表达。为了进一步明确AQPs 在野生大豆抗旱反应中的分子功能,本研究对野生大豆中的AQPs 蛋白结构进行分析,研究耐旱野生大豆材料的AQPs基因在干旱胁迫后的基因转录谱;通过转基因技术在拟南芥中过表达GsPIP2;7基因,分析基因过表达植株与野生型在干旱胁迫下的耐旱能力差异,以期明确野生大豆中AQPs基因在干旱胁迫中的表达情况,揭示GsPIP2;7在野生大豆耐干旱胁迫中的调控功能,为培育耐旱栽培大豆奠定理论基础。
1 材料与方法
1.1 试验材料种植及取样方法
高耐旱野生大豆材料永46 由河北科技师范学院野生大豆资源课题组提供。由于野生大豆籽粒外覆泥浆膜,播种之前需用小刀切种,切种在野生大豆籽粒种脐背面进行,避免破坏种脐。切种后的野生大豆籽粒播种于铺满营养土的培养钵中,上覆薄层蛭石。培养钵置于温度25 ℃(白天)/15 ℃(晚上)、日照长度14 h的温室中。野生大豆植株生长21 d 后,在营养钵中倒入300 mL 浓度为20%的PEG 6000 模拟干旱胁迫处理。干旱胁迫后分别在0(未干旱胁迫,对照)、6、12、24和48 h后随机取野生大豆材料叶片1 g,迅速保存于液氮中,作为转录组分析的样本。所有样品设3 次生物学重复。
1.2 测序数据来源
永46 在干旱胁迫下的转录谱由华大基因(北京)进行测序。提取野生大豆叶片RNA,构建cDNA 文库,利用BGISEQ-500 平台进行测序。利用Trimmomatic 0.36 软件对原始数据进行过滤,HISAT2 2.1.0 软件比对野生大豆(Glycine soja)参考基因序列。通过华大基因交互系统(https://report.bgi.com/ps/login/login.html)筛选在干旱胁迫后显著变化(|log2FoldChange|>1.5)的AQPs基因,基因表达量采用FPKM(Fregments Per Kilobase per Million)法进行计算,利用Heml 1.0.3.7 软件对取对数值(log2)的基因表达量进行聚类分析,利用MEME(http://alternate.meme-suite.org/tools/meme)在线分析工具进行蛋白结构域预测,利用TMHMM(https://services.healthtech.dtu.dk/service.php?TMHMM-2.0)在线分析工具进行蛋白跨膜结构域预测。
1.3 GsPIP2;7基因克隆及转基因拟南芥后代鉴定
提取液氮保存的野生大豆材料永46的叶片RNA,反转录为cDNA 后,利用引物F(5′→3′):GCTCTAGAA TGGCTAAAGATGTTGAGG/R(3′→5′):AGGCCCGGGT TAAGCGTTGCTTCTGAAG 扩增GsPIP2;7基因(glysoja_008314)。扩增的基因连接到pMD18 载体(D101A,宝生物工程有限公司,大连)上,后转到表达载体pCHAC(河北农业大学提供)上,以pCHAC::GsPIP2;7阳性载体转化农杆菌。拟南芥Columbia-0 生态型为受体,待植株生长至抽苔状态,将拟南芥的花蕾部位置于含有农杆菌渗透液中浸花5 min,将拟南芥植株在黑暗条件下培养24 h 后正常培养,获得T0代转基因材料。收获T0代种子种植,对生长的T1代转基因材料进行潮霉素抗性筛选,即在含有20 mg·L-1潮霉素的MS 培养基中对转基因拟南芥进行抗性筛选。培养7 d后,待拟南芥2 片子叶展开,将培养物转移至无抗生素的MS 培养基中培养,收获阳性后代转基因材料。T2代转基因材料筛选方法同T1代材料,最终获得T3代转基因材料。
1.4 转GsPIP2;7基因拟南芥基因表达分析
转基因植株GsPIP2;7基因表达量的测定参照王宇等[21]的方法。T3代纯合的转基因拟南芥种子的种植及干旱处理方法同1.1。取转基因及野生型拟南芥材料植株叶片提取总RNA,反转录得到cDNA。以cDNA为模板,以拟南芥的Actin基因作为内参基因,所用引物如下:GsPIP2;7基因引物F(5′→3′):GTCGTTGA TAAGGGCGTTGT/R(3′→5′):GCACCCAAAGCAGTACCA TT;Actin基因引物F(5′→3′):TGGAATCCACGAGACA ACCTA/R(3′→5′):TTCTGTGAACGATTCCTGGA。采用One Step SYBR®PrimeScript™ RT-PCR Kit 试剂盒(RR066A,宝生物工程有限公司,大连)进行实时荧光定量PCR(quantitative real-time PCR,qRT-PCR)反应,利用Bio-Rad CFX96 荧光定量PCR 仪(伯乐,美国)测定目的基因及内参基因的Ct 值。数据采用2-ΔΔCt法进行分析。
1.5 转GsPIP2;7基因拟南芥生长及生理指标测定
干旱胁迫后7 d进行干物质重和叶面积测定;利用根系分析系统进行植株根系测定;干旱胁迫后0 h(未干旱胁迫,对照)、12、24 和48 h 后取转基因拟南芥和野生型拟南芥同一叶位的叶片测定如下生理指标:利用氮蓝四唑(nitro-blue tetrazolium,NBT)光还原比色法测定超氧化物歧化酶(superoxide dismutase,SOD)活性;利用愈创木酚法测定过氧化物酶(peroxidase,POD)活性;利用硫代巴比妥酸(thiobarbituric acid,TBA)比色法测定丙二醛(malondialdehyde,MDA)含量;利用80%丙酮比色法测定叶绿素含量;利用过氧化氢酶测定试剂盒(A007-1-1,南京建成生物工程研究所)测定过氧化氢酶(catalase,CAT)活性。
1.6 数据分析
应用SAS 9.2 统计软件进行各处理的差异显著性分析,以P<0.05为显著水平。
2 结果与分析
2.1 野生大豆AQPs基因表达及蛋白结构分析
通过野生大豆转录组分析的基因注释结果,鉴定到野生大豆中的69个GsAQPs基因。对69个GsAQPs基因所属亚族进行分析,发现15个属于NIPs亚族,25个属于PIPs 亚族,23 个属于TIPs 亚族,6 个属于XIPs 亚族。对69 个GsAQPs 蛋白结构进行分析,发现全部GsAQPs蛋白均含有2 个NPA 基序结构,同一亚族中的蛋白具有相似的结构域,如所有的GsPIPs 均具有质膜水孔蛋白的保守结构域GGGANXXXXGY,但在其他亚族蛋白中均不含有此结构域,说明GsAQP 蛋白具有一定的保守性,并且各个亚族保守性存在差异(电子附图1)。
在干旱胁迫下,抗旱野生大豆材料中有10 个GsPIPs基因表达差异达到显著水平(图1)。10 个差异表达GsPIPs基因有两种表达模式:处理后6 和24 h 显著上调表达,GsPIP1;9、GsPIP1;12、GsPIP2;8、GsPIP2;7属于此种表达模式;处理后先下调表达,至24 h后上调表达,GsPIP2;2、GsPIP2;3、GsPIP1;4基因属于此种表达模式。对10 个差异表达GsPIPs基因的蛋白结构进行分析(图2),发现除GsPIP2;7 外,其余9 个GsPIPs 蛋白均有6个跨膜结构,由A、B、C、D、E 5个环相连接,在B 环和E 环均有1 个NPA 基序结构。鉴于GsPIP2;7 蛋白具有与其他GsPIPs 不同的独特蛋白结构,并且GsPIP2;7在干旱胁迫下显著上调表达,是上调表达最高的基因,故将其作为候选基因,研究该基因在干旱胁迫下的功能。

图1 干旱胁迫处理后野生大豆GsPIPs基因表达聚类分析Fig.1 The cluster analysis of GsPIPs in wild soybean under drought stress

图2 野生大豆GsPIPs蛋白结构域及重要结构域基序预测结果Fig.2 Prediction results of GsPIPs proteins domain and domain motif of wild soybean
2.2 GsPIP2;7基因过表达拟南芥后代筛选
转基因拟南芥阳性植株真叶长出,而阴性苗的子叶黄化,真叶不能展开,生长停滞(电子附图2)。对T1和T2代转基因植株阳、阴性分离比进行卡方测验,结果如表1 所示。转基因拟南芥的阳、阴性分离比均符合孟德尔遗传规律3∶1 的比例,说明外源基因为单显性基因,遗传规律遵循孟德尔分离定律。

表1 GsPIP2;7基因过表达拟南芥抗性筛选分离比Table 1 Isolation ratio of resistance identification in GsPIP2;7 overexpressed Arabidopsis thaliana
2.3 GsPIP2;7基因过表达拟南芥的生长指标测定
干旱胁迫4 d后,野生型拟南芥的叶片黄化失绿并且萎蔫,转基因拟南芥的叶片边缘开始卷曲,仍然保持绿色(图3)。

图3 干旱胁迫下野生型(A)与GsPIP2;7基因过表达(B)拟南芥的表型对比Fig.3 Phenotypic comparison between wild-type(A) and GsPIP2;7 overexpressed(B) Arabidopsis thaliana with drought stress
干旱胁迫7 d后,转基因拟南芥的根系总长度是野生型的1.42 倍,达到显著水平,根系表面积和总根体积与野生型之间无显著差异(表2、电子附图3)。转基因拟南芥的叶干重、根干重、单株叶面积及根冠比均显著高于野生型拟南芥。
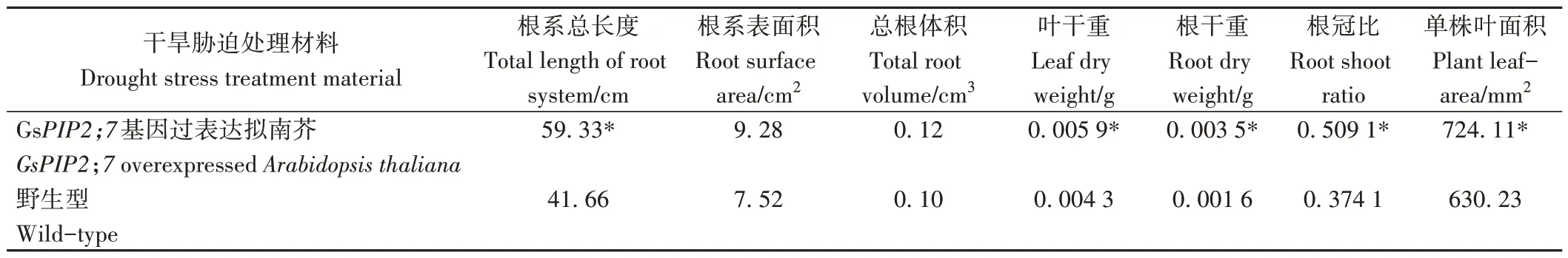
表2 干旱胁迫后GsPIP2;7基因过表达拟南芥与野生型拟南芥生长指标差异Table 2 Differences of growth indexes between GsPIP2;7 overexpressed and wild-type Arabidopsis thaliana with drought stress
2.4 GsPIP2;7 基因过表达拟南芥的基因表达及生理指标测定
对干旱胁迫下过表达拟南芥的GsPIP2;7基因表达量进行测定,结果发现,与野生型相比,过表达植株的GsPIP2;7基因表达量在干旱胁迫下的各个时间点均显著上调(图4-A)。为进一步明确转基因与野生型拟南芥在干旱胁迫下的生理差异,对氧化酶清除酶系(POD、CAT、SOD)酶活性进行了测定,发现与野生型相比,转基因植株的POD 活性在干旱胁迫后12、24和48 h均显著升高,CAT 活性在干旱胁迫后12 和24 h 显著升高,SOD 活性在干旱胁迫后48 h 显著升高(图4-B、C、F)。MDA 含量反映细胞内的脂质过氧化水平,对其进行测定发现,转基因植株的MDA 含量在干旱胁迫后48 h才有所增加,而野生型拟南芥的MDA 含量在12 h即快速升高,并且在12、24 和48 h 均较转基因植株显著增加(图4-D)。
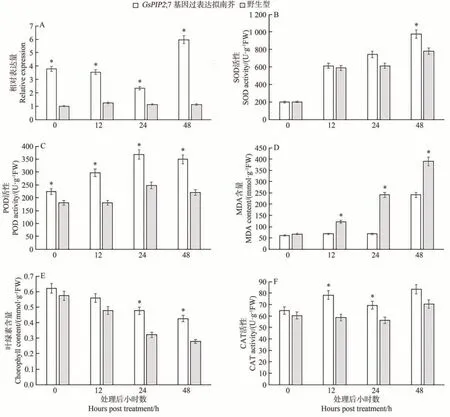
图4 干旱胁迫下GsPIP2;7基因过表达拟南芥与野生型的GsPIP2;7基因相对表达量,SOD、CAT及POD活性,MDA及叶绿素含量比较Fig.4 The GsPIP2;7 gene relative expression,SOD,POD and CAT activity,MDA and chlorophyll content of GsPIP2;7 overexpressed and wild-type Arabidopsis thaliana with drought stress
鉴于转基因拟南芥在干旱胁迫后4 d 叶片仍保持绿色(图3),对其在干旱胁迫后的叶绿素含量进行了测定。结果发现,与野生型相比,转基因植株的叶绿素含量降低速度更加缓慢,在干旱胁迫后24 和48 h 叶绿素含量差异达到显著水平(图4-E)。
3 讨论
水通道蛋白AQPs 属于主嵌入蛋白(major intrinsic protein,MIP)家族,广泛分布于植物、动物和微生物中,是调节水及小分子物质进出细胞的重要通道,对生物的正常生长具有重要作用[3]。Zhang 等[11]研究报道了野生大豆中存在的62 个AQPs,包括15 个NIPs、19 个PIPs、21个TIPs和7个SIPs。本研究在野生大豆中鉴定到69个AQPs。与Zhang等[11]的研究结果相比,新鉴定的AQPs包括2个NIPs、6个PIPs、2个TIPs和4个XIPs。本研究鉴定到7 个类SIPs(SIPs-like)蛋白结构的野生大豆蛋白,由于这些SIPs-like 蛋白不含有典型的6 个跨膜结构域,故本研究未将其归为野生大豆的AQPs。
目前,关于干旱胁迫下植物AQPs基因表达的研究报道较多[22-25]。如葡萄(Vitis vinifera)在干旱胁迫下VvPIP1;1表达量显著上调,而VvPIP2;2表达量保持不变[20]。拟南芥在干旱胁迫下的AQPs基因表达模式有很大差异,如AtPIP2;5始终上调表达(胁迫后4、12、24、48 h),AtPIP1;5、AtPIP2;3、AtPIP2;4和AtPIP2;6始终下调表达,而AtPIP1;1和AtPIP1;2则是先上调表达(胁迫后4 和12 h)后下调表达(24 和48 h)[26]。水稻在干旱胁迫下OsPIP1;1和OsPIP1;2基因表达量上调,而全部的OsPIP2s基因均表现为下调[27]。以上研究结果均说明,干旱胁迫下植物AQPs基因表达模式比较复杂。本研究发现与上述结果类似,干旱胁迫下野生大豆GsPIPs基因有两种主要表达模式:一种是“上调-下调-上调”表达模式,另一种是“下调-上调”表达模式。这进一步说明了干旱胁迫下植物AQPs基因表达受到复杂的信号网络调控,但AQPs基因在植株抗旱中的调控机制还需进一步研究。
鉴于植物中的AQPs基因数量较多,研究单个AQP基因功能成为揭示AQPs基因功能的突破口。前人在拟南芥中和小麦(Triticum aestivum)中分别过表达珠美海棠(Malus zumi)、苹果(Malus domestica)和小麦的PIP2;1基因,发现转基因植株的干旱胁迫和盐胁迫抗性增强,转基因小麦在干旱和盐胁迫下的CAT、SOD活性增加,MDA 含量较低,推测这是其比野生型植株生存能力更强的原因[28-30]。Zhang 等[31]关于柳枝稷(Panicum virgatum)PvPIP2;9基因的研究结果也发现,基因过表达植株在干旱胁迫下表现出更高的株高、叶长、地上生物量、纤维素含量和蛋白质含量,说明PIPs基因对植物耐干旱胁迫具有正向调控作用。此外,关于植物PIP2;7基因功能也有一些报道。如Khan 等[32]在拟南芥中过表达了麻风树(Jatropha curcas)的JcPIP2;7基因,发现过表达植株在盐和干旱胁迫下的根长增加,并在复水后能够正常生长,表现出比野生型植株更强的抗盐和抗干旱胁迫,Pou 等[33]也得到了相似的研究结果。本研究在拟南芥中过表达了野生大豆GsPIP2;7基因,发现与野生型相比,过表达植株在干旱胁迫下的根长更长,活性氧清除酶系的酶活性保持较高水平,这可能与细胞中较低的MDA含量有关。并且过表达植株的叶片保持绿色,叶绿素含量降低速率较慢,这都显示出过表达植株对干旱胁迫表现更强的抗性。以上结果说明GsPIP2;7基因在野生大豆耐干旱胁迫中发挥正向调控作用,但其他AQPs基因是否发挥了相同的功能尚需进一步研究。
4 结论
本研究鉴定到69个野生大豆AQPs蛋白,其中2个NIPs、6 个PIPs、2 个TIPs 和4 个XIPs 为新鉴定AQPs 蛋白;分析了干旱胁迫下高抗干旱野生大豆材料的AQPs基因表达模式,发现差异表达AQPs基因主要有“上调-下调-上调”和“下调-上调”两种表达模式。在拟南芥中过表达了野生大豆GsPIP2;7基因,发现过表达植株在干旱胁迫下的根系总长度更长,活性氧清除酶系活性较高,MDA 含量较低,叶绿素含量降低速率较慢,表现出较强的干旱胁迫耐性水平。
猜你喜欢
中学生天地(A版)(2023年1期)2023-02-17
江苏农业科学(2022年17期)2022-09-24
智慧农业导刊(2022年17期)2022-09-16
河北科技师范学院学报(2022年2期)2022-08-26
四川大学学报(自然科学版)(2021年4期)2021-07-15
中国农业科学(2020年5期)2020-03-30
信阳农林学院学报(2019年2期)2019-01-12
生命科学研究(2018年1期)2018-05-29
上海农业学报(2017年3期)2017-04-10
辽宁农业科学(2017年1期)2017-03-16
