
全香型杂交稻产量性状配合力、遗传力及相关性分析
2023-06-04 20:33:27曹应江游书梅张涛杨莉杨乾华秦俭李赓觅李昭祥罗婧高磊郑家奎蒋开锋
江苏农业科学 2023年9期
曹应江 游书梅 张涛 杨莉 杨乾华 秦俭 李赓觅 李昭祥 罗婧 高磊 郑家奎 蒋开锋
摘要:为研究全香型杂交稻亲本产量性状的配合力、遗传力及性状间相关性,利用6个香型三系不育系(品香A、J91A、J92A、3758A、德香074A、宜香1A),与7个香型恢复系(德恢9466、旌玉香针、旌香占、R9315、R9451、农香39、19香),按照NCⅡ设计的原理,共组配了42个全香型杂交稻组合,对产量相关的11个性状进行配合力、遗传力、相关性分析,并比较所配组合与生产大面积应用的宜香优2115的竞争优势。结果表明:(1)各亲本间不同的性状一般配合力差异较大,本试验就不育系而言,品香A综合表现较为优异,其产量、穗长、每穗实粒数、结实率、粒长、长宽比性状的一般配合力均为正效应。恢复系是旌香占综合表现优异,其产量、有效穗数、穗长、每穗实粒数、结实率、千粒质量、粒长、粒宽、长宽比性状一般配合力为正效应。(2)11个性状的一般配合力方差占比均大于50.00%,粒长和长宽比这2个性状占比高达100%,表明亲本间的加性效应在这些性状中占主要作用,穗长、千粒质量、粒宽和长宽比4个性状母本(不育系)一般配合力方差的占比均大于60.00%,母本效应占主要作用。产量、每穗粒数、结实率和粒长这4个性状父本(恢复系)一般配合力方差占比大于50.00%,父本效应占主要作用。(3)穗长、着粒密度、千粒质量、粒长、粒宽和长宽比6个性状的狭义和广义遗传力都大于60.00%,说明这6个性状受到环境的影响相对较小,在杂交选育时,在早期世代选择就有较好的效果。(4)产量与每穗实粒数和结实率呈极显著正相关,这说明增加每穗实粒数是提高产量的有效途径。(5)全香型杂交稻在有效穗数、每穗粒数、每穗实粒数、着粒密度、长宽比5个性状上比宜香优2115更有竞争优势。品香A和旌香占产量性状一般配合力高,容易组配出产量高的杂交稻组合,具有较大的育种利用潜力。
關键词:全香型;杂交稻;产量性状;分析;配合力;遗传力
中图分类号:S511.032 文献标志码:A 文章编号:1002-1302(2023)09-0097-06
基金项目:国家现代农业产业技术体系四川水稻创新团队建设项目(编号:2019NZN017);国家重点研发计划(编号:2017YDF0100205、2016YFD0101101);四川省重点研发计划(编号:2016NY0028)。
作者简介:曹应江(1981—),男,四川古蔺人,硕士,副研究员,主要从事水稻遗传育种研究,E-mail:caoyj987@126.com;共同第一作者:游书梅(1982—),女,四川泸县人,副研究员,主要从事水稻遗传育种研究,E-mail:yousm7322007@126.com。
通信作者:蒋开锋,博士,研究员,主要从事水稻遗传育种研究,E-mail:jiangkf67@126.com;郑家奎,博士,研究员,主要从事水稻生物学与遗传育种研究,E-mail:zhen6102@126.com。
水稻是我国的主要粮食作物。随着社会的发展和人们生活水平的提高,人们对稻米的需求已经从简单的“吃得饱”向“吃得好”改变。香稻是一种具有令人愉悦的芳香气味的特种稻资源,不仅富含多种人体必需的营养成分,而且还有滋补和药用作用,具有较高的营养价值和经济价值,受到越来越多人的喜欢。水稻的香味物质为2-乙酰基-1-吡咯啉[1]。研究表明,水稻香味受1对隐性基因控制,有香为隐性,无香为显性;在香味基因隐性纯合基因型中水稻叶片的香味与米粒的香味呈高度一致性;但在杂合的基因型中,叶片无香的单株,其米粒有不香与有香的分离[1-6]。因此,在实际的生产中,若父、母本之一是有香味杂交稻组合,其杂交F1代米饭由于部分籽粒有香味而表现出整体有香味,但由于香味籽粒的占比较少,往往导致米饭香味不够浓郁。就目前而言,生产上大多数的香稻品种,仅是母本有香味的不育系与父本为非香味恢复系组配而成的非完全香型品种,这种品种生产出的稻米只有部分籽粒有香味,如德香优系列品种[7-8]、宜香优系列品种等[9-10]。全香型杂交稻是指母本(不育系)和父本(恢复系)都是有香味的亲本配组而成的杂交稻品种,这种品种生产出的每一粒米都有香味,米饭香味更加浓郁,口感更好。本研究以品香A、德香074A、宜香1A等6个香型不育系,与旌香占等7个香型恢复系,按照NCⅡ(双列杂交设计)的原理组配了6×7=42个全香型杂交稻组合为材料,研究产量及性状的配合力、遗传力规律,这将为全香型杂交稻组合的选育提供理论参考。
1 材料与方法
1.1 供试材料
试验材料为6个香型不育系与7个香型恢复系按照NCⅡ设计,不完全杂交形成的42个全香型杂交稻组合。用宜香优2115作为竞争优势对照。6个香型不育系分别是四川省农业科学院水稻高粱研究所选育的品香A、德香074A、J91A、J95A、3758A,宜宾市农业科学院选育的宜香1A;7个恢复系分别是四川省农业科学院水稻高粱研究所选育的德恢9466、旌玉香针、旌香占、R9315、R9451,湖南省水稻研究所选育的农香39,广东省农业科学院水稻研究所选育的19香。
1.2 田间试验
田间试验于2020年在四川省农业科学院水稻高粱研究所德阳基地进行,地理位置是四川省德阳市旌阳区东湖乡大地村(104.25°E,31.10°N)。试验播种时间为2020年4月5日,移栽时间为5月8日,栽插规格为16.5 cm×33.3 cm,每份材料栽5行,每行10窝,每窝栽1粒谷苗。试验设3次重复,随机区组设计。田间管理与当地大田生产一致,及时防治病虫草害。整个试验期内气候正常,未见影响水稻生长的极端异常气候。
1.3 性状调查
成熟时去除边际效应,选取中间生长整齐的30株进行小区产量测产,另外选取中间生长较为整齐的3株取样考种,考察的性状包括有效穗数、穗长、每穗粒数、每穗实粒数、着粒密度、结实率、千粒质量、粒长、粒宽和长宽比。
1.4 数据分析
数据用Excel 2010整理后,用DPS 7.05软件计算水稻的一般配合力和其他参数,计算方法参考文献[11-12],根据固定模型估算材料的配合力效应,根据随机模型估算群体配合力遗传方差和遗传参数,结实率性状数据在方差和配合力分析时,进行反正弦转换。竞争优势计算公式:竞争优势=F1/CK×100%。式中:F1为各个全香型杂交稻组合性状值;CK为宜香优2115的性状值。
2 结果与分析
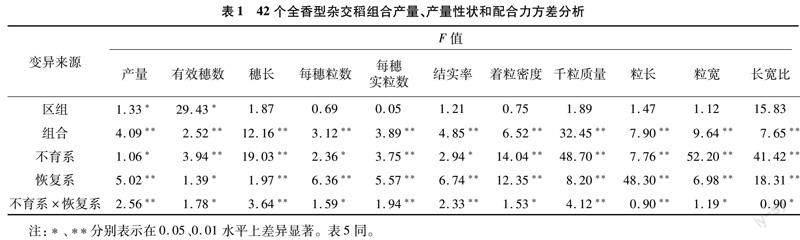
2.1 配合力方差分析
从表1可以看出,大部分性状在区组间均未达到显著差异水平,说明试验田间肥力和管理控制较为均匀。11个性状各组合间均呈极显著差异水平,说明这些组合间存在显著的遗传差异,有进行下一步分析的必要。进一步对组合的遗传差异进行配合力方差分析,发现不育系和恢复系一般配合力方差及不育系×恢复系组合间的方差均达到显著或极显著差异水平,说明这11个性状的遗传受到基因加性和非加性效应的影响。
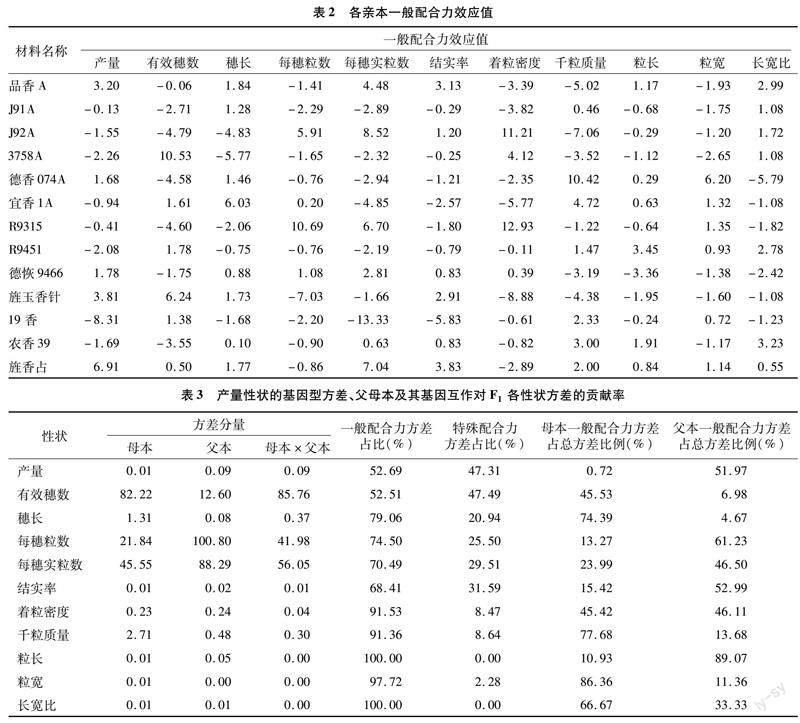
2.2 各亲本一般配合力分析
对6个香型不育系和7个香型恢复系进行配合力分析,一般配合力效应值见表2。在不育系方面,品香A综合表现较为优异,其产量、穗长、每穗实粒数、结实率、粒长、长宽比性状均为正效应。其次是德香074A,其千粒质量和粒宽表现突出,不仅是正效应,且效应值明显高于其他供试亲本。另外3758A的有效穗数一般配合力效应值较高,J92A的每穗粒数、每穗实粒数、着粒密度3个性状一般配合力效应值较高。
恢复系方面,旌香占综合表现较为优异,其产量、有效穗数、穗长、每穗实粒数、结实率、千粒质量、粒长、粒宽、长宽比性状一般配合力为正效应。其次是德恢9466 其产量、 穗长、每穗粒数、每穗实粒数、结实率、著粒密度性状一般配合力为正效应。旌玉香针的有效穗数配合力效应值较高,R9451和农香39的粒长及长宽比一般配合力效应值较高。
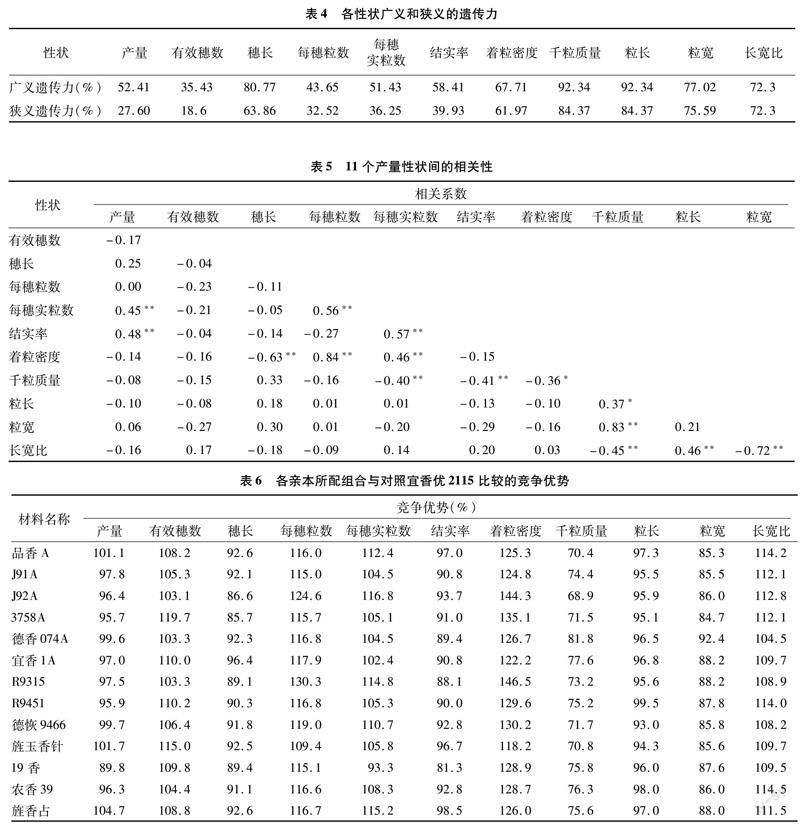
2.3 各性状基因型方差、贡献率及遗传力
利用固定模型算法对产量等11个性状的基因型方差遗传分量进行估算,结果见表3。11个性状的一般配合力方差占比均大于50.00%,粒长和长宽比这2个性状占比高达100%,表明亲本间的加性效应在这些性状占主要作用,进一步分析各亲本一般配合力方差占比发现,穗长、千粒质量、粒宽和长宽比4个性状母本(不育系)一般配合力方差的占比均大于60.00%,表明母本效应在这4个性状中对杂交F1有较大的作用。产量、每穗粒数、结实率和粒长这4个性状父本(恢复系)一般配合力方差占比大于50.00%,表明父本效应在这4个性状中对杂交一代占主要作用。
遗传力是反映性状的遗传变异占总变异的百分比。从表4中可以看出,穗长、着粒密度、千粒质量、粒长、粒宽和长宽比6个性状的狭义和广义遗传力都大于60.00%,表明这6个性状受到环境的影响较小,在杂交亲本选育时,早期世代选择就有较好的效果。其他的性状遗传力均较小,表明这些性状容易受到环境或其他因素的影响。
2.4 各性状相关性
从表5可以看出,产量与每穗实粒数和结实率达到极显著正相关 这说明增加每穗实粒数是提高产量水平的有效途径。千粒质量是产量性状中的重要性状,本试验中,千粒质量与粒长、粒宽呈显著或极显著正相关,尤其是与粒宽相关系数达到0.83,由此可以看出,增加粒长尤其是粒宽,有利于提高水稻的千粒质量。其他有显著或极显著正相关性的性状还有:每穗粒数与每穗实粒数、着粒密度,每穗实粒数与结实率、着粒密度,粒长与长宽比。另外还有穗长与着粒密度,千粒质量与每穗实粒数、结实率、着粒密度、长宽比,粒宽与长宽比6对性状间呈显著或极显著负相关。
2.5 各亲本所配组合与宜香优2115比较的竞争优势
从表6、表7可以看出,品香A、旌玉香针和旌香占3个亲本所配的组合其产量竞争优势大于100%,表明这3个亲本所配的组合产量优于对照宜香优2115。供试组合的有效穗数、每穗粒数、每穗实粒数(19香除外)、着粒密度、长宽比5个性状的竞争优势都大于100%,表明全香型杂交稻F1代组合在这5个性状上表现突出,显著优于当前生产上大面积推广应用的宜香优2115。
3 讨论与结论
3.1 全香型杂交稻主要农艺性状的遗传特点
本研究中着粒密度、千粒质量、粒长、粒宽、长宽比5个性状的一般配合力方差占比大于90.00%,说明亲本基因的加性效应在F1代的形成占绝对的主导地位。产量和有效穗数这2个性状,一般配合力和特殊配合力方差占比均约为50.00%,表明亲本基因的加性和非加性效应都对这2个性状起作用。供试材料11个性状的一般配合力和特殊配合力方差都达到极显著差异水平,说明这些产量性状在全香型杂交稻中受到基因的加性和非加性效应共同作用。这与陈俊孝等非全香型杂交稻的研究结果[13-16]相同或相似。
3.2 全香型杂交稻组合亲本选配中不育系和恢复系的相对重要性
杂交稻是母本和父本杂交后形成的F1代,杂交F1代包含了父本和母本的基因。父母本的基因对后代性状有着不同的作用。黎毛毛等研究认为,母本对杂交F1代粒型的贡献率较大[17]。刘金波等研究认为,播始历期、有效穗数、穗长等性状母本效应占主导地位,而株高、每穗粒数、结实率、千粒质量等性状父本占主导作用[15]。张征等研究认为,父本在千粒质量、单株有效穗数、株高一般配合力方面较为重要,母本在主穗长一般配合力上较为重要[18]。本研究表明,穗长、千粒质量、粒宽、长宽比这4个性状,母本一般配合力方差占比均大于50.00%,说明母本效应对杂交一代的形成占主导地位。产量、每穗粒数、结实率、粒长4个性状父本一般配合力方差占比大于50.00%,说明父本效应对杂交一代的形成占主导作用。杂交稻与常规稻比较的优势之一是父本和母本在重要性状上能实现优势互补。本试验表明,就产量性状而言,母本要注意千粒质量、粒宽和长宽比3个性状的改良和提高,而父本主要强化增加每穗粒数、结实率、粒长3个性状。
3.3 全香型杂交稻各主要农艺性状的相对重要性
水稻产量由各产量要素乘积所得,各产量要素对杂交一代的产量形成有着各自的作用。刘建丰等研究认为,有效穗数与杂交一代产量呈显著正相关[19];曾宪平等研究认为,结实率与杂交一代产量呈显著正相关[20];张进帅等研究认为,有效穗数、每穗总粒数、结实率、千粒质量四大要素均与产量呈显著或极显著正相关[21]。本试验表明,每穗实粒数和结实率这2个性状与产量呈极显著正相关,粒宽与千粒质量呈极显著正相关,因此要提高全香型杂交稻的产量水平,主要注意提高每穗实粒数、增加籽粒的千粒质量和粒宽。
3.4 各供试亲本的表现及评价
本试验中,在不育系(母本)方面,品香A综合表现较为优异,其产量、穗长、每穗实粒数、结实率、粒长、长宽比等性状均为正效应,值得大量应用和测配其他恢复系。其次是德香074A,其千粒质量和粒宽一般配合力均为正效应,且效应值明显高于其他供试亲本。
在恢复系方面,旌香占综合表现较为优异,其产量、有效穗数、穗长、每穗实粒数、结实率、千粒质量、粒长、粒宽、长宽比等性状一般配合力为正效应。其次是德恢9466,其产量、穗长、每穗粒数、每穗实粒数、结实率、着粒密度等性状一般配合力为正效应。
参考文献:
[1]Bradbury L M T,Fitzgerald T L,Henry R J,et al. The gene for fragrance in rice[J]. Plant Biotechnology Journal,2005,3(3):363-370.
[2]彭 波,孙艳芳,陈报阳,等. 水稻香味基因及其在育种中的应用研究进展[J]. 植物学报,2017,52(6):797-807.
[3]陆艳婷,刘庆龙,王俊敏,等. 利用等位基因特异扩增快速检测水稻香味基因[J]. 作物学报,2008,34(2):243-246.
[4]王 丰,李金华,柳武革,等. 一种水稻香味基因功能标记的开发[J]. 中国水稻科学,2008,22(4):347-352.
[5]李金华,王 丰,柳武革,等. 水稻粤丰B的香味遗传分析与SSR标记定位[J]. 分子植物育种,2006,4(1):54-58.
[6]张 涛,张红宇,蒋开锋,等. 水稻香味基因的精细定位[J]. 分子植物育种,2008,6(6):1038-1044.
[7]郑家奎,蒋开锋,杨乾华,等. 杂交水稻新组合德香4103的选育及高产原因初析[J]. 杂交水稻,2008,23(6):13-15.
[8]曹应江,游书梅,蒋开锋,等. 香型优质水稻三系不育系德香074A产量性状配合力分析[J]. 中国稻米,2020,26(3):84-87.
[9]包灵丰,林 纲,赵德明,等. 水稻籼型优质不育系宜香1A特性研究[J]. 植物遗传资源学报,2007,8(1):86-90.
[10]包灵丰,林 纲,赵德明,等. 水稻三系不育系宜香1A香味遗传研究及产量配和力分析[J]. 西南农业学报,2006,19(5):974-976.
[11]莫惠栋. p×q交配模式的配合力分析(续)[J]. 江苏农学院学报,1982,3(4):53-57.
[12]莫惠栋. P×q交配模式的配合力分析[J]. 江苏农学院学报,1982,3(3):51-57.
[13]陈俊孝,游艾青,刘 凯. 杂交水稻表型、杂种优势和配合力的关系[J]. 湖北农业科学,2020,59(24):24-29.
[14]曹应江,游书梅,蒋开锋,等. 水稻产量性状一般配合力QTL定位[J]. 核农学报,2020,34(9):1921-1932.
[15]刘金波,宋兆强,王宝祥,等. 几个新选杂交水稻不育系和恢复系的主要农艺性状的配合力分析[J]. 西南农业学报,2016,29(2):209-213.
[16]曾庆宁,吕建群,陈 林,等. 四川及南亚水稻恢复系农艺性状的配合力及遗传力分析[J]. 西南农业学报,2014,27(2):463-468.
[17]黎毛毛,徐 磊,刘昌文,等. 水稻粒形遗传及QTLs定位研究进展[J]. 中国农业科技导报,2008,10(1):34-42.
[18]张 征,张雪丽,莫博程,等. 籼型杂交水稻农艺性状的配合力研究[J]. 作物学报,2017,43(10):1448-1457.
[19]刘建丰,陈光辉,何 强,等. 不同产量水平杂交稻产量构成因素的分析[J]. 云南农业大学学报,2006,21(6):707-710,720.
[20]曾宪平,何 芳,吕建群. 四川2001—2010年审定水稻品种农藝与米质性状的典型相关分析[J]. 西南农业学报,2011,24(5):1631-1635.
[21]张进帅,黄翠红,王 慧,等. 水稻单株产量相关性状的相关性及其通径分析[J]. 江苏农业科学,2019,47(18):108-113.
猜你喜欢
西北农业学报(2024年7期)2024-07-12 20:26:59
农业科技通讯(2023年1期)2023-02-12 07:09:28
养猪(2022年4期)2022-08-17 07:07:02
猪业科学(2018年5期)2018-07-17 05:56:14
农家科技下旬刊(2016年9期)2016-12-15 08:54:34
河北农业科学(2016年2期)2016-09-09 01:41:31
江苏农业科学(2015年11期)2016-01-27 14:50:33
湖北畜牧兽医(2015年11期)2016-01-11 10:08:24
天津农业科学(2015年10期)2015-10-20 00:15:45
湖北农业科学(2014年7期)2014-06-28 10:29:07
