
极低浮游动物丰度生境食蚊鱼仔稚鱼食物组成与摄食策略
2023-06-01 04:58:14陈国柱
四川动物 2023年3期
陈国柱
(1.西南林业大学云南省高原湿地保护修复与生态服务重点实验室,昆明 650224;2.西南林业大学云南省滇池湿地生态系统国家定位观测研究站,昆明 650224;3.西南林业大学国家高原湿地研究中心/湿地学院,昆明 650224)
云贵高原地处喀斯特地貌区域,独特的岩溶地下河和出露岩溶泉是该区鱼类演化的关键物质基础。岩溶水体常因营养贫乏而使饵料资源受限。长期演化中,土著鱼类以特化形式适应了此类栖息地,而外来入侵种能否或如何适应这种栖息地是有待探明的重要科学问题。浮游动物丰度被认为是制约依赖于浮游动物为食的鱼类仔稚鱼早期存活的关键生态因子(殷名称,1991)。在岩溶水体中,极低的浮游动物丰度将是考验入侵鱼类仔稚鱼早期生存的关键性因素,观察入侵鱼类仔稚鱼在饵料资源不足生境中的适应策略将有助于揭示其关键入侵生态学机制。
星云湖是云南九大高原湖泊之一,面积30 余km2,近代以来陷入了重度富营养化污染及大量入侵种入侵的双重困境。尽管如此,在湖周广阔的面山区域分布着相当丰富的岩溶泉,当地习惯称为“龙潭”,许多土著鱼类孑遗种群得以保存,如长臀云南鳅Yunnanilus analis、长鳔盘Discogobio macrophysallidos(金锦锦等,2021)。这些水体水质清冽但营养贫乏,初步调查显示其浮游动物丰度周年维持在极低水平。然而,在长期的观察中发现,入侵种食蚊鱼Gambusia affinis在许多泉眼中已形成稳定种群。食蚊鱼是世界入侵范围最广泛的小型鱼类(Krumholz,1948;陈国柱等,2008a),1920s 经上海引入中国大陆(潘炯华等,1980),它对云贵高原泉潭生境的入侵可能对该区关键生物多样性造成重要的影响。Fairfax等(2007)指出,食蚊鱼的入侵是美国荒漠地区以泉眼为主要栖息生境的小型特有鱼类濒危或灭绝的关键因素。作为入侵者,食蚊鱼仔稚鱼如何成功克服极低饵料丰度所带来的困境?深入了解该问题将为理解食蚊鱼在极端环境中的生存与扩散策略提供重要帮助。
本文以星云湖“天中”龙潭群食蚊鱼种群为研究对象,针对其生境浮游动物丰度、仔稚鱼食物组成及摄食策略等问题展开研究,以揭示食蚊鱼仔稚鱼对极低饵料丰度生境的适应性及适应策略,为进一步探索其早期生活史及入侵机制奠定基础。
1 材料与方法
1.1 研究区域
“天中”龙潭群位于云南省玉溪市江川区大街镇大营村村委会摆寨村以北、江华公路两侧,星云湖东南角(102°47′54.10″E,24°16′58.63″N,海拔1 776 m),自北向南记为1~5号龙潭,1号、5号龙潭间直线距离约800 m。1号龙潭最大,面积近50 m2,最大水深约2.5 m;其余4 个龙潭蓄水面在6~10 m2之间,最大水深约0.6 m,各以30~40 cm 宽出水水道与下游农田灌溉渠道网连接。各龙潭均为当地农业主要灌溉水源。食蚊鱼在3 号、4 号、5 号龙潭的数量较多,2 号龙潭较少,1 号龙潭则因2020 年建造烟叶烘干房而硬化覆盖了南侧出水道,食蚊鱼种群数量降至偶见。各龙潭中主要的沉水植物为黑藻Hydrilla verticillata,在5—8 月可形成植被覆盖水面,但常被当地人周期性清理。在蔬菜收获期,各龙潭为菜农清洗蔬菜场所。流水周年不断,在3 号龙潭对侧马路旁山岩为一抽水房,定期抽水作为饮用水源。本研究以3 号、4 号龙潭为食蚊鱼样品采集地;1~5 号龙潭均进行浮游动物样品采集与分析。
1.2 样品采集
2021—2022 年每年5 月底“天中”龙潭区域食蚊鱼开始繁殖,6 月初仔稚鱼逐步增多,但较星云湖湖区的种群延后约1 个月。在食蚊鱼繁殖盛期采集仔稚鱼,并对应在仔稚鱼出现盛期(6—7月)、消失期(11 月)采集浮游动物样品,以检视其周年变动情况。
浮 游 动 物 样 品 在2021 年6 月14 日、11 月14 日,2022 年6 月25 日、7 月16 日共采集4 次。采集时,以500 mL塑料量杯在龙潭中采集10 L水样,由13 号浮游动物采集网过滤浓缩至25 mL 水体后转入50 mL离心管,即加入2~3 mL甲醛固定待检。
鱼类样品采集在2021 年6 月14 日、6 月19 日,2022年6月25日、7月16日进行。以网孔2 mm,网口直径40 cm的手抄网采集,10%甲醛固定待检。
样品分析在室内进行。在显微镜下鉴定浮游动物种类,清点数量。在体视显微镜下解剖食蚊鱼仔稚鱼,鉴定及清点其消化道中食物种类、数量等。
食蚊鱼体长9.5 mm以下或腹鳍未完全长成的仔稚鱼对水体浮游动物饵料资源高度依赖。为探究特定贫营养水体对其早期摄食的影响,以体长<9.5 mm的食蚊鱼仔稚鱼为研究对象,因此,文中若其无特别指出体长等的未性成熟个体,则依原文献统一表述为幼鱼。
1.3 数据处理
1.3.1 指标计算 以常规鱼类摄食生态学指标(窦硕增,1996;薛莹,金显仕,2003)等进行分析:
出现频率(F/%)=100Fi/Fs,式中,Fi为某一食物种类在其消化道中出现的食蚊鱼仔稚鱼总数,Fs为总检查食蚊鱼仔稚鱼数。
数量百分比(N/%)=100Ni/Ns,式中,Ni为某一食物种类在食蚊鱼仔稚鱼消化道中的数量,Ns为食蚊鱼仔稚鱼消化道中各种食物种类总数。
摄食强度(ind./ind.)=某一食物种类在平均每尾食蚊鱼仔稚鱼消化道中的数量。
食物充塞度:以肠中食物团占据肠道的长短判定其所占肠道比例,定义为食物充塞度指数。完全空肠定为0,肠道充满食物且前端略膨大定为1,分析时以食物团长度占据肠道比例在0~1 间连续定性取值。
特定饵料丰度(%)=食蚊鱼仔稚鱼消化道某种食物的数量/出现该种食物的食蚊鱼仔稚鱼消化道中食物的数量。
肥满度(K)=100×体重/体长3(mg·mm-3)。
1.3.2 统计分析 单因素相似性分析(ANOSIM)比较年际、不同地点食蚊鱼仔稚鱼食物组成的差异性,显著性水平设为α=0.05。
食蚊鱼仔稚鱼摄食策略分析采用Amundsen等(1996)修改的Costello(1990)图形法,以特定饵料丰度(%)与出现频率(%)作图。利用该图形分析摄食策略时,集中在右上角的饵料为主要饵料,左下角为稀有饵料;若饵料集中在左上角,则表明种内个体间食物组成差异较大,而当饵料集中在右下角,则表明种内个体间食物组成差异较小,重叠程度大;饵料集中在特定饵料丰度轴中线下侧,且沿x轴方向分布,为广食性策略;而饵料集中在中线上侧,为狭食性策略。
2 结果
2.1 浮游动物丰度
2021—2022年4次调查中,各龙潭中浮游动物饵料丰度均维持在极低水平,仅2021 年6 月14 日1号龙潭中的丰度达18.4 ind.·L-1(表1)。
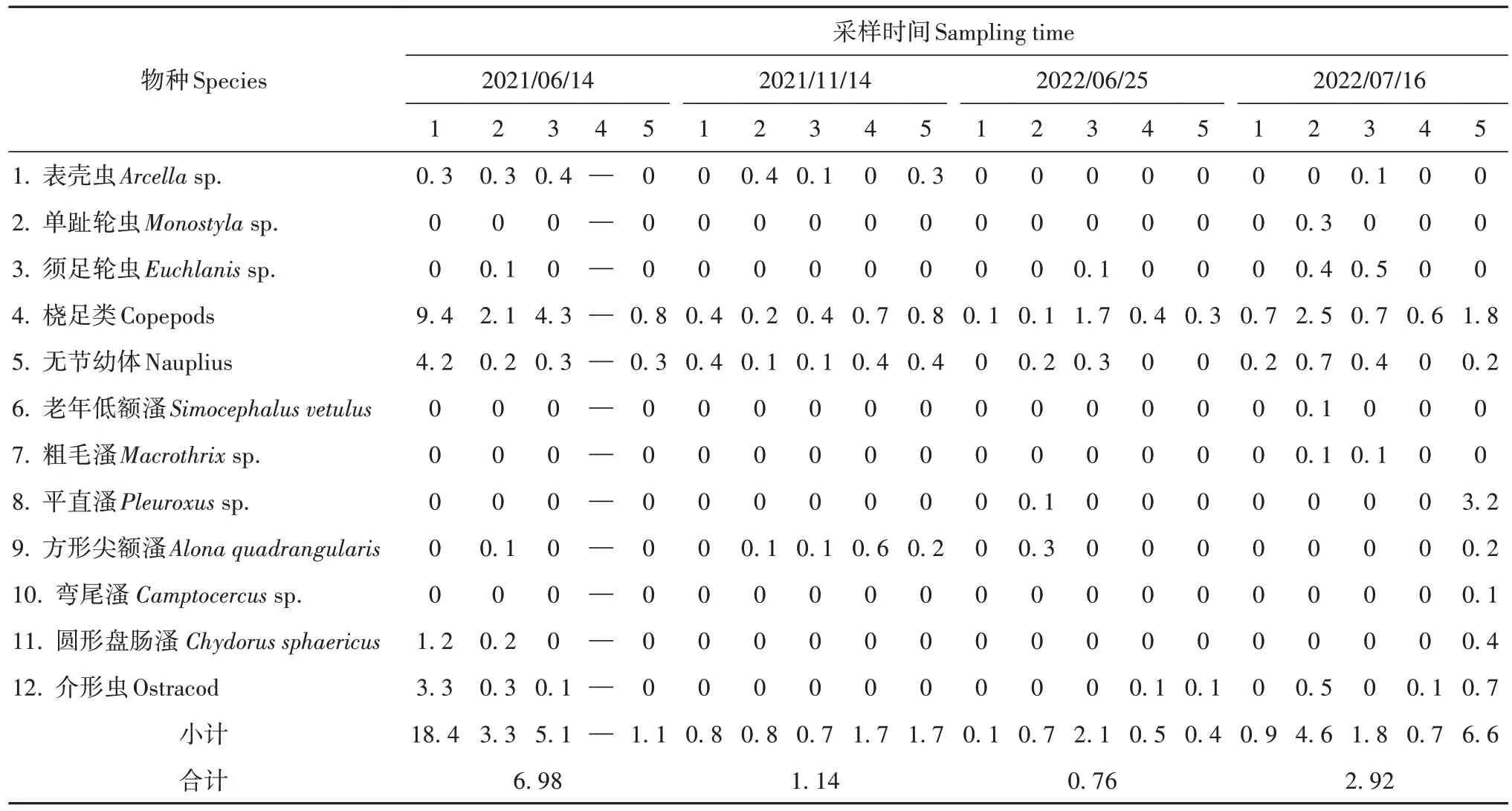
表1 “天中”龙潭群浮游动物丰度Table 1 Zooplankton abundance in the Tianzhong springs
各次调查总体均值未超过7 ind.·L-1,分别为6.98 ind.·L-1、1.14 ind.·L-1、0.76 ind.·L-1和2.92 ind.·L-1,周年均值为2.95 ind.·L-1。浮游动物中桡足类及其无节幼体占绝对数量优势,4 次调查分别占77.4%、68.4%、81.6%和53.4%。
2.2 食物组成
3 号龙潭2021 年、2022 年食蚊鱼仔稚鱼消化道食物充塞度指数依次为0.78±0.09、0.72±0.13,4 号龙潭的为0.41±0.26、0.66±0.12。整体均值为0.70±0.16,食物充塞度水平较高。
食蚊鱼仔稚鱼消化道中共检出31 种食物类型:桡足类、卵及幼体在出现频率、数量百分比上均占据了绝对优势(主要是锯缘真剑水溞Eucylops serrulatus)(表2);对桡足类的摄食强度分别为:3 号 龙 潭20.2 ind./ind.±11.7 ind./ind.(2021 年)、12.1 ind./ind.±5.7 ind./ind.(2022 年);4 号 龙 潭4.1 ind./ind.±3.8 ind./ind.(2021 年)、4.1 ind./ind.±5.4 ind./ind.(2022年)。
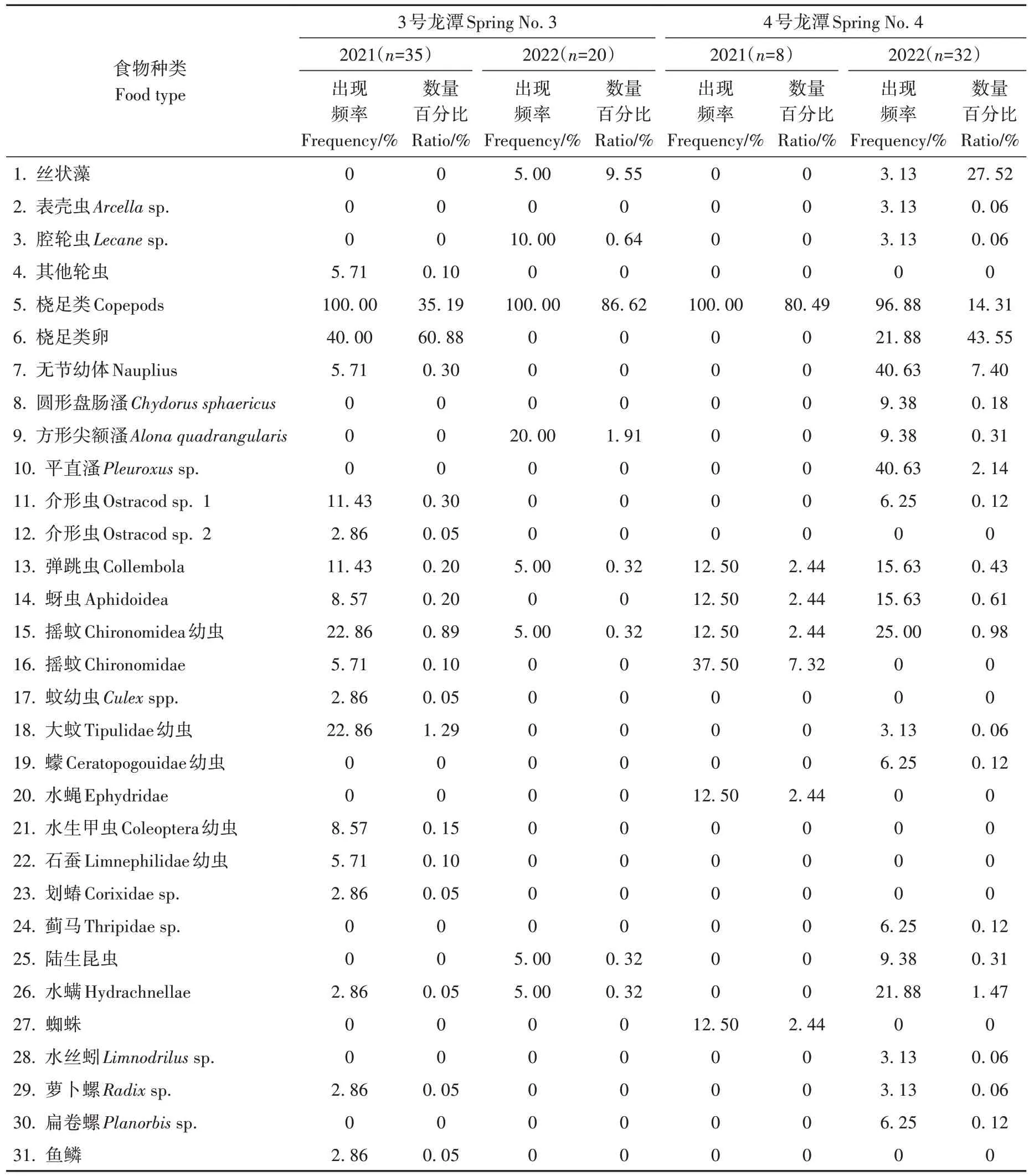
表2 “天中”龙潭群食蚊鱼仔稚鱼食物组成Table 2 Food composition of larval-juvenile mosquitofish in Tianzhong Springs
3号龙潭中食蚊鱼仔稚鱼在2021年、2022年检出的食物多样性指数分别为0.45±0.35(0~1.39)、0.13±0.26(0~0.94)。4号龙潭为0.54±0.44(0~1.30)、0.56±0.39(0~1.39)。在所有样品中未检出空肠个体(食物充塞度为0),而食物多样性指数为0(仅摄食1种食物)的个体则达24.3%。
单因素相似性分析(ANOSIM)显示,3、4 号龙潭中食蚊鱼仔稚鱼各自年际间食物组成并未检出具有统计学意义的差异(P>0.05),R值分别为0.057、0.145;但两地点间具有显著差异(P<0.01),R=0.201。
2.3 摄食策略
2021年,3号龙潭中食蚊鱼仔稚鱼的主要食物是桡足类及其卵,其余食物种类虽多,但为稀有食物,表现出以桡足类及其卵为主要食物的狭食性策略(图1:a);4 号龙潭中,桡足类表现出极高的特定饵料丰度,呈现明显的狭食性策略(图1:b)。

图1 3号(a、c)和4号(b、d)“天中”龙潭中食蚊鱼仔稚鱼摄食策略分析Fig.1 Feeding strategy of larval-juvenile mosquitofish in No.3 (a,c)and No.4 (b,d)Tianzhong Springs
2022 年,3 号龙潭中食蚊鱼仔稚鱼食物种类少,桡足类为主要食物,呈现明显的狭食性策略(图1:c);4 号龙潭中,食蚊鱼仔稚鱼食物种类较多,且多数食物在特定饵料丰度中线下侧,偏向于广食性策略(图1:d)。
2.4 肥满度
在3 号、4 号龙潭中,食蚊鱼仔稚鱼在2021 年、2022 年的肥满度指数分别为2.1±0.2、2.2±0.2、2.2±2.6、2.1±2.8,总体均值为2.1±2.4。地点、年份间均无统计学差异。
3 讨论
3.1 食蚊鱼仔稚鱼对低饵料丰度生境的适应
当浮游动物饵料丰度低于一定水平,仔稚鱼的存活就会受到严重影响(Houde,1987;Milleret al.,1988;殷名称,1991)。对大鳞副泥鳅Paramisgurnus dabryanus(李新蕊等,2018)、花鲈Lateolabrax japonicus(谢仰杰等,2004)、大眼狮鲈Sander vitreus仔鱼(Zornet al.,2020)的研究均显示其饵料密度需大于100 ind.·L-1才能存活或获得良好的生长状况。美国密歇根湖湖湾春季大型浮游动物丰度较长时间低于5 ind.·L-1(Zornet al.,2020),是造成大眼狮鲈仔鱼补充困难引发年际种群数量波动的关键因素(Mayet al.,2021)。
研究显示,“天中”龙潭群水体浮游动物丰度维持在极低水平,周年总体均值不超过3 ind.·L-1。这种饵料丰度水平对其他鱼类仔稚鱼的生存是危险因素,但对食蚊鱼仔稚鱼则并未构成明显的威胁。与金锦锦等(2020)的数据比较,“天中”龙潭种群食蚊鱼仔稚鱼肥满度指数与饵料较为丰富的滇池花红园种群(>300 ind.·L-1)并无统计学差异(2.1±2.4 vs.2.7±3.5);且食物充塞度情况显著高于后者(0.70±0.16 vs.0.55±0.18,P<0.05)。解剖镜下仔稚鱼肠道皱褶充盈饱满,未检出遭受饥饿胁迫所呈现的皱褶松弛、溃散现象;它们主要摄食的桡足类(锯缘真剑水溞)形体也饱满匀称,是生长状态良好的饵料。这些都表明在低浮游动物饵料丰度条件下,食蚊鱼仔稚鱼仍获得良好生长。
极强的早期摄食能力(Pyke,2005;陈国柱等,2008b)可能是食蚊鱼仔稚鱼能够适应低饵料丰度生境的关键。食蚊鱼为卵胎生鱼类,初产仔鱼(稚鱼)在产出后2 h 左右便可摄食外源食物,口裂宽可达2 mm(陈国柱等,2008b),这些特点可以扩大其饵料供应范围,是对不同生境形成适应性的关键生物学基础。而完善的身体发育状况赋予食蚊鱼仔稚鱼早期极强的游动能力,支持其对水体大范围搜索,提高了其在低饵料丰度环境中仍能获取足够食物的几率。对鲱Clupea harengus(Munk& Kiorboe,1985)和大西洋鳕Gadus morhua仔鱼(Puvanendran & Brown,2002)的研究显示,仔鱼能通过增加觅食时间,扩大搜索范围来应对低饵料丰度条件胁迫。尽管如此,要确切厘清食蚊鱼仔稚鱼对低饵料丰度条件的摄食适应机制,尚需开展更深入的实验观察。
3.2 桡足类对食蚊鱼仔稚鱼生存与种群扩张潜力的重要性
桡足类在营养条件较低、捕食压力较大的环境下仍能保持较强的种群稳定能力,与其自身生物学特点(快速运动能力和高繁殖力)密切相关。在许多水体中,桡足类成为关键性饵料类群(杨关铭等,1999),如在抚仙湖,桡足类及其幼体占甲壳类浮游动物数量比的60%以上(潘继征等,2009);在新丰江水库即使加入轮虫,其数量比也超过60%(赵帅营,韩博平,2007);在“天中”龙潭中,其数量比周年变化为53.4%~81.6%。桡足类从无节幼体到成蚤,形成连续尺寸的食物资源,是仔幼鱼及小型鱼类的可持续利用的重要资源(朱成德,1986)。“天中”龙潭的食蚊鱼仔稚鱼食物组成年际间未检出显著差异,显示以桡足类为核心的食物链在该区域形成了稳定的运行系统,保证了食蚊鱼仔稚鱼狭食性策略能够实现。
桡足类在世界各地均是食蚊鱼(G.affinis、G.holbrooki)的常见食物之一,对其仔稚鱼的生存及种群扩展均具有重要作用:在匈牙利Hévíz 湖中食蚊鱼幼鱼消化道桡足类的出现频率约为5%(Specziár,2004);在土耳其农田生境的为9.0%(Sokolov & Chvaliova,1936);在法国卡玛格湿地,7 月可达73.4%(Crivelli & Boy,1987);在昆明花红园,8 月 为59.6%,10—11 月 为74.7%(金 锦 锦 等,2020);而本研究区域的则高达100%。在印度奈尼塔尔湖,桡足类及其无节幼体在食蚊鱼幼鱼消化道的出现频率分别为20%、53.7%(Singh & Gupa,2010)。而以成鱼及幼鱼群体一起统计的研究显示,桡足类在食蚊鱼消化道的出现频率在中国广州的最高值可达100%(潘炯华等,1980);在西班牙巴塞罗那巴尼奥莱斯湖的周年均值为46.1%(García-Berthou,1999);在土耳其塞伊汗大坝湖则为9.8%(Erguden,2013)。从摄食量来看,食蚊鱼幼鱼在葡萄牙蒙德戈河谷稻田摄食桡足类的均值为14.6 ind./ind.(Cabralet al.,1998);在昆明花红园 为2.7 ind./ind.(8 月),3.5 ind./ind.(10—11 月)(金锦锦等,2020);在本研究区域为4.1~20.2 ind./ind.。显然,桡足类在许多生境中都是食蚊鱼重要食物资源,是其能广泛扩散的重要物质基础。
另一方面,桡足类是许多高海拔贫营养水体的优势浮游动物物种(如滇西北横断山区),且物种多样性较高(舒树森等,2013)。本文研究结果提示,有桡足类等食物资源支持,使得食蚊鱼具有向高海拔贫营养水体区域入侵扩散的重要潜力及现实性。滇西北海拔3 000 m 以上等尚未被食蚊鱼入侵的区域,避免从食蚊鱼入侵区域引入养殖鱼类与放流物种是阻止食蚊鱼入侵的关键策略。
猜你喜欢
水产科学(2022年1期)2022-01-26 08:43:00
渔业科学进展(2021年6期)2022-01-04 06:26:42
中华民居(2020年2期)2020-09-12 06:52:28
读友·少年文学(清雅版)(2019年1期)2019-05-09 02:35:30
河北渔业(2019年1期)2019-02-13 01:35:34
水利技术监督(2017年3期)2017-06-09 06:55:34
海洋渔业(2016年6期)2016-04-16 03:00:32
人间(2015年19期)2016-01-04 12:46:46
应用海洋学学报(2015年3期)2015-11-22 07:39:22
河南科技(2015年21期)2015-03-26 21:21:21
