
基于MaxEnt预测未来气候条件下钻叶紫菀在中国的潜在适生区
2023-05-30 09:17:23李建宇陈燕婷郭燕青何玉仙傅建炜史梦竹
植物保护 2023年2期
李建宇 陈燕婷 郭燕青 何玉仙 傅建炜 史梦竹

摘要 鉆叶紫菀Symphyotrichum subulatum是一级恶性入侵植物,其传播速度快,繁殖能力强,严重影响本地草本植物多样性。本研究利用生态位模型MaxEnt和地理信息系统软件ArcGIS,根据收集的钻叶紫菀分布点和环境变量,预测目前气候条件和未来气候变化情景下钻叶紫菀在中国的潜在适生区,并推测环境变量对钻叶紫菀适生区的影响。结果表明,钻叶紫菀的潜在适生区主要位于亚热带湿润型气候大区的低海拔区域,其中高适生区的分布范围为21°~40°N,主要位于广东、福建、江西、江苏、湖南、湖北、四川东南地区和安徽东北地区等。影响钻叶紫菀潜在适生区分布的关键环境变量为最干旱月份降水量、最湿润月份降水量、10月平均温度、3月最高温度、8月最低温度和6月太阳辐射量。在未来气候变化情景SSP126、SSP245、SSP370和SSP585条件下,钻叶紫菀的低、中和高适生区面积占比将分别增加3.54%、4.80%、5.73%和3.29%。整体而言,未来气候变化有利于钻叶紫菀适生区的扩张。到21世纪中叶,钻叶紫菀的高适生区呈现向高海拔地区和中高纬度(具体为由南向北和西北方向)扩张的趋势。钻叶紫菀适生区分布范围较广,建议加强适生区的果蔬、苗木等农产品调运的检验检疫工作,以防止钻叶紫菀的进一步扩散。
关键词 钻叶紫菀; MaxEnt模型; 气候变化; 适生区; 入侵植物
中图分类号: Q948
文献标识码: A
DOI: 10.16688/j.zwbh.2022344
Abstract Symphyotrichum subulatum, as a malignant invasive plant, has a rapid spread and strong reproductive capacity, which seriously affected the diversity of local herbaceous plants. Based on the known distribution points and environmental variables collected, MaxEnt model and ArcGIS were used to predict the potential geographical distribution of S.subulatum in China and the changes of its suitable areas under future climate scenarios. The relationship between the distribution of S.subulatum and environmental variables was also investigated. The results showed that the suitable areas of S.subulatum were mainly distributed in subtropical humid climate regions. The highly suitable areas of S.subulatum distributed from 21°N to 40°N, including Guangdong, Fujian, Jiangxi, Jiangsu, Hunan, Hubei, southeast Sichuan and northeast Anhui. The key environmental variables that determine the distribution location of S.subulatum are precipitation of driest month, precipitation of wettest month, mean temperature of October, maximum temperature of March, minimum temperature of August and solar radiation of June. Under future climate scenarios (SSP126, SSP245, SSP370 and SSP585), the percentage of lowly, moderately, and highly suitable areas of S.subulatum will increase by 3.54%, 4.80%, 5.73% and 3.29%, respectively. By the middle of the 21st century, the suitable area of S.subulatum will expand to middle and high latitude (from south to north and northwest) and higher elevation area. Overall, future climate change benefit to the expansion of S.subulatum. It is necessary to strengthen inspection and quarantine in the trade of fruits, vegetables, seedlings and other agricultural products to prevent its further spread.
Key words Symphyotrichum subulatum; MaxEnt model; climate change; suitable area; invasive plant
气候是决定物种分布的重要因素之一[1-2]。政府间气候变化专门委员会[3]发布的数据显示,1850年-1900年,全球地表平均温度已经上升约1℃,且预计2021年-2040年全球气温将上升1.5℃。许多研究表明,未来气候变化将会对外来入侵植物的空间分布格局产生重要影响。李安等[4]的研究结果显示,未来气候条件下,黄顶菊Flaveria bidentis (L.) Kuntze的适生分布范围将进一步扩大。加拿大一枝黄花Solidago canadensis L.在我国的适生区面积随时间的推移而增大[5];而南美蟛蜞菊Sphagneticola trilobata (L.) Pruski在未来总适生区面积变化不大,但分布格局发生变化,最佳适生区有向西南地区转移的趋势[6]。因此,研究气候变化背景下外来入侵植物的适宜生境以及气候变化对这些适宜生境的影响,对于采取有效措施控制外来入侵植物的传播具有重要意义,同时可为农业生产、生物多样性保护和生态系统可持续发展等提供策略依据。
物种分布模型(species distribution model,SDM)是一个基于生态位理论,根据物种的已知分布点和相关环境因素,预测物种在当前或未来气候条件下适生区的重要建模工具[7-9]。结合全球气候变化的环境因素和气体排放情景RCPs(代表性浓度路径,representative concentration pathways)、SSPs(共享社会经济路径,shared socialeconomy pathways)等,SDM能够预测未来气候背景下物种适生区的变化。常用的SDM包括BIOCLIM(bioclimate analysis and prediction system)[10]、DOMAIN[11]、GARP(genetic algorithm for rule-set production)[12]和MaxEnt(maximum entropy modeling)[13]等。相比于其他模型,MaxEnt模型对样本量的要求不高,并且当分布点与环境变量相关性不明的情况下也能得到较好的预测结果[7, 14-17]。因此,MaxEnt被广泛应用于外来入侵植物的适生区预测[4-6,18]。
钻叶紫菀Symphyotrichum subulatum (Michx.) G. L. Nesom,又名钻形紫菀,异名Aster subulatus,属于菊科Asteraceae联毛紫菀属Symphyotrichum,一年生草本植物,高25~150 cm,叶片狭长细小,叶片卵形至倒披针形、钻形;头状花序,舌状花带状,管状花黄色。钻叶紫菀能够产生大量的带冠毛瘦果,瘦果轻小,随风散布入侵,其扩散距离可超过100 m[19]。钻叶紫菀原产于北美洲,目前在全球范围内广泛分布[20-21]。1827年,钻叶紫菀入侵至我国澳门,随后在1947年扩散至湖北武昌,此后广泛分布于广西、广东、台湾、福建、湖北、江西、安徽、江苏、浙江、云南、贵州、四川、重庆、山东、河南和河北等省份,其分布北界为北京[22-23]。由于植株耐盐碱、喜湿润,易沿路边河道蔓延,快速传播并侵入河岸带生态系统[24]。钻叶紫菀全草具有化感物质,能够抑制周围植物的生长和发育,如茎叶水浸液会抑制油菜、小麦、绿豆等植物的根系生长;危害秋收作物(棉花、大豆、甘薯等),有时在田埂边形成单优势种群,大量消耗土壤营养,使得其他植物难以生存。该杂草也常侵入浅水湿地,影响湿地生态系统及其景观[25]。因此,环境保护部办公厅于2014年将其列入中国外来入侵物种名单(第三批)[26]。
目前对钻叶紫菀的研究主要集中在特定地区的新发报道[20,27-28]、生物学特性[19,29-30]以及提取利用[31-32]等,而关于钻叶紫菀的潜在分布预测研究则较少。Guan等[33]采用集合预测的方法模拟不同气候情景下包括钻叶紫菀在内的11种入侵物种的范围变化,预测结果发现,受全球气候变暖的影响,11种入侵植物的适生范围均向北扩展。目前尚未见有利用MaxEnt模型结合全球不同时期的环境变量,对钻叶紫菀在中国的适生区进行预测的研究报道。因此,本研究基于MaxEnt模型对钻叶紫菀在中国的适生区进行预测,同时运用刀切法和环境变量响应曲线评估影响钻叶紫菀分布的环境因子,分析二者之间的关系,为钻叶紫菀的预测预报和有效防控提供参考。
1 材料与方法
1.1 数据来源与处理
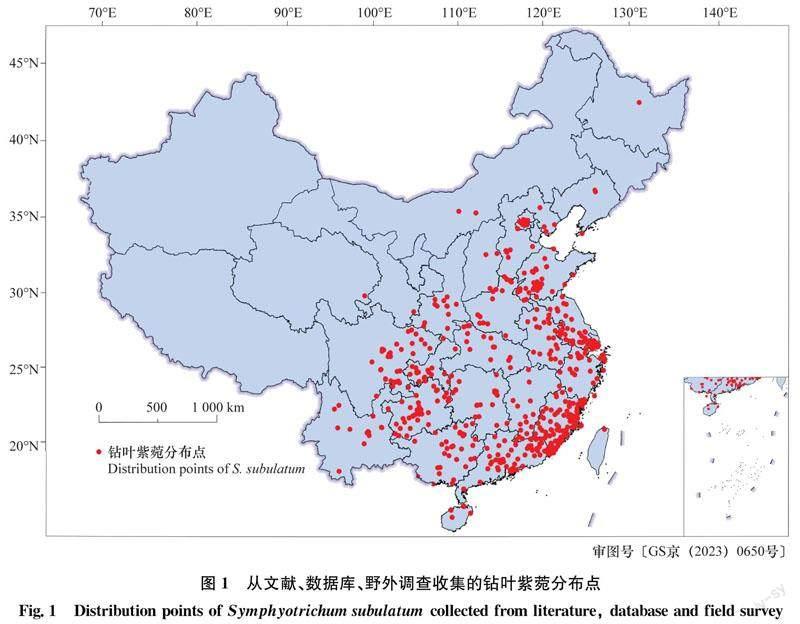
入侵植物钻叶紫菀分布点主要来自:1)野外调查数据:调查方法和样地点选择参考《外来草本植物普查技术规程》(NY/T 1861-2010),調查数据均记录于福建省农业科学院数字农业研究所研发的农林科学数据云采集农业大数据平台(http:∥www.datacatcher.cn);2)平台数据:主要采集自中国数字植物标本馆(CVH,https:∥www.cvh.ac.cn/)、国家标本平台(NSII,http:∥www.nsii.org.cn/2017/)、中国自然标本馆(CFH,https:∥cfh.ac.cn/)和中国植物图像库(PPBC,http:∥ppbc.iplant.cn/);3)文献资料查阅:通过查阅国内外公开发表的文献,获得该物种的分布信息。
基于上述数据进行进一步的过滤和筛选,去除位置信息不明确的分布点。本文所选择的环境变量分辨率为2.5弧分(arc-minute),设置缓冲区为2.5 km,缓冲区内仅保留1个样本点[34]。
野外调查数据包含210个钻叶紫菀的分布点;从CVH、NSII、CFH和PPBC网站上获得585个分布点;通过文献查阅获得12个分布点,共计807个分布点。经过数据过滤和筛选,去除重复分布点,共获得639个钻叶紫菀的有效分布点,如图1所示。
1.2 环境变量数据
本研究选取的环境变量包括1970年-2000年的11个温度相关环境变量(Bio1~Bio11)、8个降水量相关环境变量(Bio12~Bio19)和84个其他环境变量(如每月的太阳辐射、风速和水蒸气压等)(表1)。未来气候数据选取的是第6次国际耦合模型相互比较项目(coupled model intercomparison projectphase 6,CMIP6)中的北京气候中心气候系统模式中等分辨率版本(BCC-CSM2_MR),包括未来3个时期(2041-2060、2061-2080和2081-2100)的4种SSPs最新排放情景数据,即SSP126、SSP245、SSP370和SSP585[34]。环境变量均下载自WorldClim(https:∥worldclim.org/),分辨率为2.5弧分。
1.3 地图数據
本研究采用的中国地图来源于国家测绘地理信息局标准地图服务网站(http:∥bzdt.ch.mnr.gov.cn)。
1.4 模型构建方法
1.4.1 MaxEnt模型建模及精度检测
将获得的钻叶紫菀分布点坐标及环境变量导入MaxEnt模型(Version 3.4.1,https:∥github.com/mrmaxent/Maxent)中,随机选取75%的分布点用于模型构建,剩余25%的分布点用于模型验证与测试,迭代500次,其他参数设定值为默认。
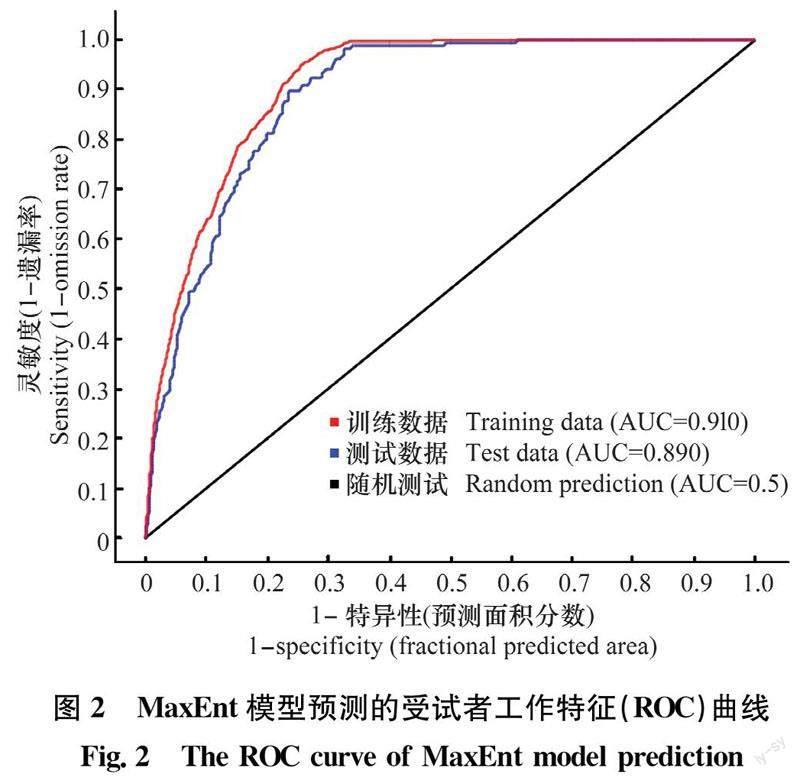
MaxEnt模型预测的准确性以受试者工作特征曲线(receiver operating characteristic curve,ROC曲线)的曲线下面积(area under curve,AUC)表示。AUC值为0.5~1,越接近1,预测精度越高。当AUC≥0.9时,模型预测最优;0.8≤AUC<0.9时,模型预测较好;0.7≤AUC<0.8时,模型预测一般;0.6≤AUC<0.7时,模型预测较差;0.5≤AUC<0.6时,模型预测失败[35]。
1.4.2 环境变量相关性检验和筛选
通过MaxEnt模型对103个环境变量进行运算,获得各环境变量的贡献率,保留贡献率大于1.0%的环境变量。环境变量过多,各因子间容易产生多重共线性(过拟合)问题,影响模型运算结果。因此,利用ArcMap软件和SPSS数据统计软件对环境变量进行Pearson相关性检验(Pearsons r),当两个变量之间的|r| > 0.8,说明二者存在共线性关系,保留贡献率高的环境变量,去除贡献率低的环境变量,当二者贡献率相同时,选择与其他环境变量共线性关系小的环境变量。贡献率> 1.0%且无共线性的环境变量均用于MaxEnt模型建模。采用刀切法(Jackknife method)分析环境变量对钻叶紫菀分布产生影响的重要程度。
1.4.3 适生区等级划分
基于MaxEnt输出的钻叶紫菀的存在概率逻辑值(P),范围为0~1,利用ArcGIS软件的重分类(reclassify)工具将其适生区等级划为4个等级:高适生区(P≥0.5)、中适生区(0.25≤P<0.5)、低适生区(0.05≤P<0.25)、非适生区(P<0.05),获得钻叶紫菀适生区图。
2 结果与分析
2.1 MaxEnt模型精确性分析
受试者工作特征(ROC)曲线如图2所示,训练数据AUC值为0.910,测试数据AUC值为0.890,模拟结果表明MaxEnt模型预测钻叶紫菀适生区精度较高。
2.2 环境变量筛选结果
经过MaxEnt建模,选择贡献率>1.0%的环境变量共14个,对这些变量进行Pearson相关性分析后(图3),最终筛选出7个主要的环境变量:最湿月份降水量(Bio13)、最干月份降水量(Bio14)、10月降水量(Prec10)、6月太阳辐射(Srad06)、10月平均温度(Tavg10)、3月最高温度(Tmax03)和8月最低温度(Tmin08)。
分析各环境变量对钻叶紫菀分布影响的贡献率,发现影响钻叶紫菀分布的主要因素是降水量,其次是温度。降水量贡献率达63.3%,分别为最干月份的降水量(Bio14,45.7%)和最湿月份降水量(Bio13,17.6%);温度贡献率占27.2%,分别为10月平均温度(Tavg10,16.7%)、8月最低温度(Tmin08,7.9%)和3月最高温度(Tmax03,2.6%)。上述5个环境变量是影响钻叶紫菀分布的关键因子。
2.3 环境变量对钻叶紫菀适生区预测的影响
通过Jackknife分析可知(图4):仅有单一环境变量时,正规化训练收益影响的环境变量由高到低为:Tavg10 > Tmin08 > Bio14 > Bio13 > Tmax03 > Prec10 > Srad06。表明影响钻叶紫菀地理分布的主要环境变量为:气温变量(10月平均温度和8月最低温度)和降水变量(最干月份降水量和最湿月份降水量)。
根据钻叶紫菀对环境变量的响应情况,设置存在概率阈值为0.5,当存在概率大于0.5时,对应的环境变量适合钻叶紫菀的生长,10月平均温度高于15℃时(图5a),适合钻叶紫菀的生存,最适平均温度为24℃;8月最低温度高于21℃时(图5b),适合钻叶紫菀的生存,高于26℃时,存在概率最高;最干月份降水量范围在18~72 mm(图5c),当降水量为23 mm时,钻叶紫菀存在的概率最高(0.88);最湿月份降水量范围在150~420 mm(图5d),钻叶紫菀存在概率最高时(0.78)的降水量为250 mm;3月最高温范围在11~24℃(图6e),最适温度为17℃;10月降水量范围在38~152 mm(图5f),最适降水量为49 mm;6月太阳辐射高于19 200 kJ/(m2·d)时(图5g),钻叶紫菀存在概率低,当太阳辐射低于14 900 kJ/(m2·d)时,最适合钻叶紫菀的生存。
2.4 钻叶紫菀目前在中国的适生区预测
当前钻叶紫菀的适生区分布在18.1°N~44.4°N,91.0°E~125.5°E,总适生面积大约为307.82×103 km2,占我国国土面积的32.10%。其中,高适生区面积为95.50×103 km2,占总适生区面积的31.02%,主要分布在我国广东、福建,江西、江苏、湖南、湖北、四川东南地区和安徽东北地区等;中适生区面积为128.04×103 km2,占总适生区面积的41.60%,主要分布在我国四川东部、贵州、河南、山东、湖北和安徽西南地区等;低适生区面积为84.29×103 km2,占总适生区面积的27.38%,主要分布在我国云南、陕西、河北北部地区和辽宁等(图6)。
2.5 未来气候情景下钻叶紫菀的适生区预测
統计不同气候情景下适生区面积及变化(表2),结果如下:总适生区面积预计为312.77 ×103 ~325.46×103 km2,比当前分布增加1.61%~5.73%。2041年-2060年,SSP126、SSP245、SSP370和SSP585情景下钻叶紫菀的适生区面积分别占我国国土总面积的33.24%、33.64%、33.94%和33.16%,与当前的适生区面积相比,分别增加了3.54%、4.80%、5.73%和3.29%(图7a~d)。相比当前的潜在分布面积,2061年-2080年钻叶紫菀的潜在分布面积分别增加了3.77%、2.93%、5.73%和3.75%(图7e~h)。2081年-2100年的潜在分布面积则分别增加了3.57%、3.07%、1.61%和3.29%(图7i~l)。
适生区总面积增幅最大的是2061年-2080年SSP370情景下,增加了17×103 km2;增幅最小的为2081年-2100年SSP370情景下,为4.95×103 km2。高适生区增幅最大的是2041年-2060年及2081年-2100年SSP585情景下的,增幅达到95.68%;增幅最小的为2081年-2100年SSP245情景下的,增幅为86.01%。由图7可知:未来的钻叶紫菀适生分布区在不断增加,除海南、云南、四川、部分山东、部分河南和陕西外,大部分省份几乎变为高适生区;藏东南地区由低适生区提高到高适生区,新增低适生区有新疆准噶尔盆地至塔里木盆地边缘。钻叶紫菀在河北北部和辽宁西南地区适生程度降为非适生区。
2.6 未来气候模式下钻叶紫菀的适生区的海拔变化情况
从表3中可知,未来情景下适生区总面积增加1.61%~5.73%,且各年份及SSPs间的适生区总面积差异不大,2081年-2100年SSP126的适生区变化居于中间值,因此本文选取该年预测结果来分析钻叶紫菀未来的适生区海拔变化情况。
海拔1 164 m以下,未来气候情况下,钻叶紫菀的中适生区面积减少(图8b),高适生区面积增加(图8a),总适生区海拔变化不明显(图8d)。由此可知,未来气候更适宜钻叶紫菀的生长,其高适生区有向高海拔扩张的趋势。
3 结论与讨论
3.1 钻叶紫菀在中国的适生区
预测结果显示,钻叶紫菀适生区分布主要受到降水量的影响(贡献率63.3%),分布区主要在400 mm年等降水量线(大兴安岭—张家口—兰州—拉萨—喜马拉雅山脉)的东南地区。高适生区和中适生区主要分布于湿润区,低适生区则主要分布于半湿润区。其次适生区受到温度的影响(贡献率27.2%),高适生区大部分在亚热带地区,中适生区分布于亚热带及暖温带,低适生区分布于热带及其他温度带边缘交界处。
华北平原年均温度较低的地方仍然是钻叶紫菀的高适生区,说明在海拔较低的地区,海拔高度可能是影响钻叶紫菀分布的重要因素之一。钻叶紫菀的高适生区包含了华北平原、长江中下游平原、四川盆地和武夷山脉、南岭等低海拔地区。中适生区包含了云贵高原贵州地区、秦岭以东和太行山脉以东地区。低适生区分布于云贵高原、黄土高原、太行山脉及台湾山脉等地区。青藏高原地区(西藏、青海和部分新疆、甘肃、四川、云南)和内蒙古地区地处干旱区和半干旱区,且海拔高;黑龙江、吉林、辽宁东北地区地处中温带,温度低;内蒙古高原海拔高,地处中温带。因此,钻叶紫菀在这些地区的适生度低,且适生区面积小。
不同纬度区域的气候类型不同,一般来说纬度越低,气温越高[36]。本研究预测结果显示钻叶紫菀适生区虽广,但最适宜生长的分布区较为集中,高适生区主要集中在21°N~40°N,说明纬度可能对钻叶紫菀的分布产生一定的限制作用。因此,对钻叶紫菀集中分布区的小范围生境的分析,有助于揭示钻叶紫菀的入侵机制。
3.2 环境变量对钻叶紫菀适生区的影响
最干月份降水量(Bio14)和最湿月份降水量(Bio13)这两个环境变量与钻叶紫菀适生区的关系最为密切。模型预测结果表明:最干月份降水量为23 mm时,钻叶紫菀存在概率最高,当最干月份降水量超过160 mm时,钻叶紫菀存在概率较低。最湿月份降水量的适宜范围为150~420 mm,当最湿月份降水量高于740 mm时,钻叶紫菀存在概率仅为0.18。虽然当前没有研究揭示降水量与钻叶紫菀分布的关系,但是大多数研究表明降水量对植物的增长有显著的影响[2,33,37-38]。
3月最高温度对钻叶紫菀
的分布有一定的影响,3月最高温适宜范围为11~24℃,钻叶紫菀的存在概率最高,当3月最高温为17℃时,存在概率为0.84。有文献记载云南省有钻叶紫菀分布和危害[20-21],而本研究结果显示钻叶紫菀在云南省内以低适生区为主,北部地区为中适生区,这可能与云南省南北温差较大有关,平均温度在5~24℃,南北气温相差可达19℃左右。在福建和广东等省份,则由于全省各地温差较小,全省均是高适生区。因此,在对不同区域具体分析时,还应考虑是否有其他特定因素对钻叶紫菀分布造成影响。
3.3 未來气候变化下钻叶紫菀分布区变迁与防治策略
研究表明,气候变暖使得大多数动植物在纬度梯度向两极方向迁移[39-40],但是,这种物种分布纬度变化的方向和幅度却存在许多争议[41-42]。本研究发现,未来气候条件下,钻叶紫菀的适生区明显扩张,且总体趋势以低海拔向高海拔、中低纬度向中高纬度(具体为由南向北和西北方向)扩张为主,这一结果与Guan等[33]的预测基本一致。有研究证明由降水主导和由温度主导的物种移动方向不同[43],钻叶紫菀在温暖多雨的气候下生长较好,而不适应高温干旱。气候变化中水分条件和水热关联性的指标对钻叶紫菀生长与分布均有很大的影响。海拔高度的变化影响地区的水热条件,海拔越高,温度越低,水汽增加。在未来气候变化情景下,全球温度呈现上升趋势,钻叶紫菀分布区很可能往高海拔地区扩散。
在未来气候情景下,钻叶紫菀的适生区增加了新疆区域。目前尚未在新疆发现钻叶紫菀分布,而且从中国地形看,钻叶紫菀通过自然扩散方式进入新疆区域的概率较小,但由于新疆旅游业的发展,吸引大批量游客,因此需预防游客或其他人员无意带入。对于适生区,加强检疫,精选种子,防止通过作物种子夹带;同时,在已发生区域对该杂草进行有效控制,在钻叶紫菀开花前进行人工铲除或使用除草剂,并配合深耕土壤,将植株埋入土中腐败,深埋种子能够抑制种子的萌发[21]。
参考文献
[1] OCONNOR B, BOJINSKI S, RSLI C, et al. Monitoring global changes in biodiversity and climate is more important than ever as ecological crisis intensifies [J/OL]. Ecological Informatics, 2020, 55: 101033. DOI: 10.1016/j.ecoinf.2019.101033.
[2] 陈禹光, 乐新贵, 陈宇涵, 等. 基于MaxEnt模型预测气候变化下杉木在中国的潜在地理分布[J]. 应用生态学报, 2022, 33(5): 1207-1214.
[3] IPCC. Climate change 2021: The physical science basis [EB/OL]. (2021 -08-09) [2022-06-18]. https:∥www.ipcc.ch/report/ar6/wg1.
[4] 李安, 李良涛, 高萌萌, 等. 基于MaxEnt模型和气候变化情景入侵种黄顶菊在中国的分布区预测[J]. 农学学报, 2020, 10(1): 60-67.
[5] 潘铭心, 朱思睿, 张震. 外来入侵植物加拿大一枝黄花在中国的适生区预测[J]. 西安文理学院学报(自然科学版), 2022, 25(1): 90-96.
[6] 肖雨沙, 郑洁宁, 李红春, 等.气候变化背景下南美蟛蜞菊在中国的潜在适生区预测[J].生态科学, 2021, 40(4): 75-82.
[7] 乔慧捷, 胡军华, 黄继红. 生态位模型的理论基础、发展方向与挑战[J]. 中国科学: 生命科学, 2013, 43(11): 915-927.
[8] GUISAN A, ZIMMERMANN N E. Predictive habitat distribution models in ecology [J]. Ecological Modelling, 2000, 135(2/3): 147-186.
[9] 黄敏毅, 孔晓泉, 段仁燕, 等. 虚拟物种的基本原理及其在物种分布模型评估中的应用[J]. 生态学报, 2016, 36(9): 2460-2470.
[10]NIX H A. A biogeographic analysis of Australian elapid snakes [M]∥LONGINORE R. Atlas of elapid snakes of Australia. Canberra: Australian Government Publishing Service, 1986: 4-15.
[11]CARPENTER G, GILLISON A, WINTER J. DOMAIN: A flexible modelling procedure for mapping potential distributions of plants and animals [J]. Biodiversity and Conservation, 1993, 2(6): 667-680.
[12]STOCKWELL D. The GARP modelling system: problems and solution to automated spatial prediction [J]. International Journal of Geographical Information Science, 1999, 3(2): 143-158.
[13]PHILLIPS S J, ANDERSON R P, SCHAPIRE R E. Maximum entropy modeling of species geographic distributions [J]. Ecological Modelling, 2006, 190(3/4): 231-259.
[14]ELITH J, GRAHAM C H, ANDERSON P R, et al. Novel methods improve prediction of species distributions from occurrence data [J]. Ecography, 2006, 29(2): 129-151.
[15]WISZ M S, HIJMANS R J, LI J, et al. Effects of sample size on the performance of species distribution models [J]. Diversity and Distributions, 2008, 14(5): 763-773.
[16]朱耿平, 刘强, 高玉葆. 提高生态位模型转移能力来模拟入侵物种的潜在分布[J]. 生物多样性, 2014, 22(2): 223-230.
[17]陈新美, 雷渊才, 张雄清, 等. 样本量对MaxEnt模型预测物种分布精度和稳定性的影响[J]. 林业科学, 2012, 48(1): 53-59.
[18]郭燕青, 史梦竹, 李建宇, 等. 基于MaxEnt模型的假臭草潜在分布区预测[J]. 热带亚热带植物学报, 2019, 27(3): 250-260.
[19]诸葛晓龙, 朱敏, 季璐, 等. 入侵杂草小飞蓬和钻形紫菀种子风传扩散生物学特性研究[J]. 农业环境科学学报, 2011, 30(10): 1978-1984.
[20]莫训强, 孟伟庆, 李洪远. 天津3种外来植物新记录一长芒苋, 瘤梗甘薯和钻叶紫菀[J]. 天津师范大学学报(自然版), 2017, 37(2): 36-38.
[21]徐海根, 强胜. 中国外来入侵生物[M]. 北京: 科学出版社, 2018: 517-518.
[22]潘玉梅, 唐赛春, 岑艳喜, 等. 钻形紫菀开花期种群构件的生物量分配[J]. 热带亚热带植物学报, 2010, 18(2): 176-181.
[23]贺萍, 邓禹君, 胡潇尹, 等. 钻形紫菀对小麦与白菜潜在化感作用的研究[J]. 草业学报, 2019, 28(10): 101-109.
[24]苏思. 伊洛河流域外来草本植物分布格局及其对不同干扰强度的响应[D]. 郑州: 河南大学,2010.
[25]许桂芳, 刘艳侠. 钻形紫菀化感作用及危害评价[J]. 安徽农业科学, 2006,34(16): 4032-4033.
[26]中华人民共和国生态环境部. 关于发布中国外来入侵物种名单(第三批)的公告[EB/OL].(2014-08-15) [2022-06-18]. http:∥www.mee.gov.cn/gkml/hbb/bgg/201408/t20140828_288367.htm.
[27]刘传才, 崔力拓, 韩桂君. 河北省15种新记录植物[J]. 中国环境管理干部学院学报, 2011, 21(5): 36-37.
[28]曾宪锋, 邱贺媛, 齐淑艳, 等. 环渤海地区1种新记录入侵植物——钻形紫菀[J]. 广东农业科学, 2012, 39(24): 189.
[29]HU Xin, ZHANG Yun, LUO Jun, et al. Accumulation and quantitative estimates of airborne lead for a wild plant (Aster subulatus) [J]. Chemosphere, 2011, 82(10): 1351-1357.
[30]YUAN Yongqiang, YU Shen, BAUELOS G S, et al. Accumulation of Cr, Cd, Pb, Cu, and Zn by plants in tanning sludge storage sites: opportunities for contamination bioindication and phytoremediation [J]. Environmental Science and Pollution Research, 2016, 23(22): 22477-22487.
[31]AYAZ F, KKBOYACI N, DEMIRCI B. Essential oil composition and antimicrobial activity of Aster subulatus Michx. from Turkey [J]. Records of Natural Products, 2017, 11(4): 389-394.
[32]CHENG Huiyuan, WANG Shu, WEI Mei, et al. Effect of leaf water extracts of four Asteraceae alien invasive plants on germination performance of Lactuca sativa L. under acid deposition [J]. Plant Ecology, 2021, 222(4): 433-443.
[33]GUAN Bicai, GUO Hanjing, CHEN Sisi, et al. Shifting ranges of eleven invasive alien plants in China in the face of climate change [J/OL]. Ecological Informatics, 2020, 55: 101024. DOI: 10.1016/j.ecoinf.2019.101024.
[34]刘婷, 曹家豪, 齐瑞, 等. 基于GIS和MaxEnt模型分析气候变化背景下紫果云杉的潛在分布区[J]. 西北植物学报, 2022, 42(3): 481-491.
[35]SWETS J A. Measuring the accuracy of diagnostic systems [J]. Science, 1988, 240(4857): 1285-1293.
[36]马世炎, 于洪春, 赵奎军, 等. 基于MaxEnt模型的大豆蚜全球潜在地理分布分析[J]. 昆虫学报, 2022, 65(5): 630-637.
[37]王书越, 潘少安, 王明睿, 等. 基于MaxEnt模型评估刺五加在东北地区的空间分布[J]. 生态学报, 2019, 39(9): 3277-3286.
[38]塞依丁·海米提, 努尔巴依·阿布都沙力克, 许仲林, 等. 气候变化情景下外来入侵植物刺苍耳在新疆的潜在分布格局模拟[J]. 生态学报, 2019, 39(5): 1551-1559.
[39]LENOIR J, SVENNING J C. Latitudinal and elevational range shifts under contemporary climate change [J]. Encyclopedia Biodiversity, 2013, 4(1): 599-611.
[40]PARMESAN C. Ecological and evolutionary responses to recent climate change [J]. Annual Review of Ecology Evolution and Systematics, 2006, 37(1): 637-669.
[41]GUO Fengyi, LENOIR J, BONEBRAKE T C. Author correction: Land-use change interacts with climate to determine elevational species redistribution [J/OL]. Nature Communications, 2020, 11: 3485.DOI: 10.1038/s41467-020-17319-w.
[42]LENOIR J, GGOUT J C, MARQUET P A, et al. A significant upward shift in plant species optimum elevation during the 20th century [J]. Science, 2008, 320(5884): 1768-1771.
[43]BELLARD C, BERTELSMEIER C, LEADLEY P, et al. Impacts of climate change on the future of biodiversity [J]. Ecology Letters, 2012, 15(4): 365-377.
(責任编辑:杨明丽)
猜你喜欢
四川林业科技(2023年6期)2024-01-07 06:16:12
英语世界(2023年10期)2023-11-17 09:19:12
中国野生植物资源(2023年9期)2023-10-20 09:11:34
中国中医药信息杂志(2023年6期)2023-06-13 02:05:36
西北植物学报(2022年3期)2022-04-18 01:40:50
资源节约与环保(2022年12期)2022-02-02 02:39:16
中华环境(2021年9期)2021-10-14 07:51:16
植物资源与环境学报(2020年2期)2020-04-20 07:18:02
竹子学报(2018年2期)2018-10-25 02:02:12
生态文明新时代(2018年4期)2018-04-25 11:57:12
