
麦类作物粒色形成机制及基因定位研究进展
2023-05-29 15:21王燕姚晓华安立昆姚有华白羿雄吴昆仑
安徽农业科学 2023年9期
关键词:花青素
王燕 姚晓华 安立昆 姚有华 白羿雄 吴昆仑
摘要 有色麦类作物的形成是由于籽粒的种皮或糊粉层中积累了花青素,从而导致其籽粒呈现出多种颜色。有色麦类作物中含有丰富的蛋白质、维生素、膳食纤维及矿物微量元素等,兼具抗氧化、延缓衰老、调理肠胃等多方面保健功能。综述了国内外有关学者对于有色麦类作物的主要营养与功能组成成分、植物花青素的合成调控机制以及籽粒颜色基因定位3个方面的相关研究进展,探讨了目前有色麦类作物粒色基因定位过程中存在的问题,展望了有色麦类作物的发展趋势,以期为今后有色麦类作物籽粒颜色的调控机制研究提供理论依据,也为能够筛选有利基因,创制有色麦类新种质提供参考。
关键词 麦类作物;籽粒颜色;花青素;基因定位
中图分类号 S512 文献标识码 A 文章编号 0517-6611(2023)09-0001-06
doi:10.3969/j.issn.0517-6611.2023.09.001
Abstract Coloured wheat crops are formed due to the accumulation of anthocyanins in the seed coat or dextrin layer of the seeds, resulting in a variety of colours in the seeds. Coloured wheat crops are rich in protein, vitamins, dietary fibre and mineral trace elements, and have various health functions such as antioxidant, antiageing, and gastrointestinal care. This paper reviews the progress of research on the main nutritional and functional components of coloured wheat crops, the regulatory mechanism of phycocyanin synthesis and the gene localisation of grain colour, discusses the problems in the gene localisation of coloured wheat crops, and outlooks the development trend of coloured wheat crops. We hope to provide a theoretical basis for future research on the regulatory mechanisms of grain colour in coloured wheat crops, and to provide valuable reference information for the screening of favourable genes and the creation of new and superior coloured wheat germplasm.
Key words Wheat crop;Grain colour;Anthocyanin;Gene targeting
基金项目 国家自然科学基金(31960427);国家大麦产业技术体系项目(CARS-05);国家重点研发项目(2020YFD1001403)。
作者简介 王燕(1996—),女,四川遂宁人,硕士研究生,研究方向:作物遗传育种。
*通信作者,副研究员,硕士生导师,从事青稞遗传育种研究。
随着人民生活水平的日益提高,麦类作物消费向多元化的发展,普通麦类作物已经不能够完全满足人们的生活消费所需。近年来人们越来越多地通过食用全麦类食品来获得健康益处,且诸多研究也关注于谷物组织中植物化学物质的组成、分布和功能特性,使麦类作物的品质改良需要向着更多新方向进行探索和发展。有色麦类作物中富含碳水化合物、多酚、膳食纤维、矿物质及其他营养物质,被认为是一种理想的人类健康食品,因此开发有色麦类作物或许可以作为麦类作物研究的一个新方向[1]。此外,有色麦类作物是珍贵的麦类种质资源,其颜色的呈现与有色麦类作物中富含的天然花色苷类化合物的形成有关[2-3]。目前,较为常见的有色麦类籽粒颜色主要有蓝色、黑色和紫色,它们分别受不同的显性基因的控制[4]。再者,有色麦类作物中含有氨基酸、维生素、天然色素及多种微量矿质元素等多种人体稀缺的营养成分。这些成分目前已被广泛应用于食品工业、生物制药、化妆品等多个领域,具有较大的潜在开发应用前景[5-7],因此进一步开发有色麦类作物的营养资源受到了国内外学者的广泛关注。该研究概述了国内外学者对于有色麦类作物的主要营养和功能组成成分、花青素的合成调控机制及有色麦类作物粒色基因定位等相关研究,旨在为今后更深入地研究有色麦类作物的粒色形成机制和挖掘粒色控制基因提供理论基础。
1 有色麦类作物主要营养和化学组分概述
近年来,有色麦类作物因其具有特殊营养价值和保健功能而备受青睐。有色麦类作物较普通麦类作物而言含有更为丰富的营养成分和化学组分,尤其是花色苷、蛋白质和氨基酸、维生素、膳食纤维、膳食纤维、矿物微量元素等。这些营养组分在食品健康、医疗保健、美容等领域受到了广泛关注。
1.1 花色苷
花色苷是一种天然存在的水溶性色素,它能夠促使许多谷物类、水果和蔬菜等呈现出不同颜色[8-9]。花色苷之所以引起人们的兴趣,主要有两方面的原因,首先是它们可以作为一种天然着色剂应用于工业生产技术领域;其次是在人类健康领域具有重要作用,如有色麦类作物中所含有的天然花色苷种类较为丰富,在促进人体健康和预防心血管疾病等方面的作用效果显著[10-11]。叶琳[12]通过采用超高效液相色谱-四极杆-飞行时间质谱仪测定了紫粒小麦科兴617、蓝粒小麦科兴611以及普通白粒小麦济麦22中的花青素种类与含量,结果发现,3种小麦中共鉴定得到14种花色苷单体,尤以紫粒小麦科兴617中的花色苷种类最多,共计11种;蓝粒小麦科兴611中的花青素种类次之,含有9种;普通白粒小麦济麦22花青素种类最少,仅含3种。赵善仓等[13]应用极管阵列检测器液相色谱法(HPLC-PDA)和串联超高效液相色谱-串联四级杆质谱(UPLC-MS/MS)分别对蓝粒小麦、紫粒小麦紫繁3、白粒小麦远5987-88和黑小麦76 4种有色小麦籽粒中的花色苷提取物进行全波段扫描,共鉴定出14种不同类型的花色苷,其中黑小麦76和蓝粒小麦包含了9种花色苷;紫粒小麦紫繁3包含了8种花色苷,白粒小麦远5987-88仅包含了7种花色苷。Abdel-Aal等[14]通过对黄色大麦和紫色大麦进行花色苷种类与含量检测,结果发现2种有色大麦中共检测出17种花色苷单体,均包含飞燕草花色素-3-葡萄糖苷、牵牛花色素-3-葡萄糖苷等重要花色苷成分,但紫色大麦中含有4种已知花色苷成分和其他9种未被识别的花色苷成分,而黄色大麦中仅检测到5种花色苷成分。Zhang等[9]利用液相色谱-质谱(LC-MS)对紫青稞麸皮中的花青素提取物进行成分与表征分析,结果共发现6种花色苷。可见,虽然花色苷在不同颜色麦类作物中所含成分略有差异,但与白粒相比,有色麦类作物中花色苷成分多样且含量丰富。
研究表明,花色苷具有抗氧化[15]、抗肿瘤[16-17]、降低心血管疾病[18]、抗阿尔兹海默症[19]等多种功能,符合人们对健康、营养、绿色食品的发展要求。另外,以往研究中也证实了花色苷在防止紫外光照射,减缓辐射造成的皮肤表面损伤等方面效果显著,可被用于化妆品与相关美容制品的生产[20]。此外,花色苷作为一种天然的色素,还可作为多种颜色的着色剂应用于食品加工领域[21]。因此,开发有色麦类作物中的花色苷成分对于麦类作物资源的开发和利用具有重要意义。
1.2 蛋白质和氨基酸
有色麦类作物中的蛋白质与氨基酸含量总体高于普通麦类作物,营养价值较为丰富。研究表明[22],黑粒小麦76的氨基酸总量比普通白粒小麦增加了29.8%,显著高于普通小麦;且黑粒小麦76所含的人体所需8种必需氨基酸以及9种非必需氨基酸含量与普通小麦相比增加了约30%。袁园园等[23]通过测定有色小麦中蛋白质含量发现,蓝、紫粒小麦中蛋白质含量分别达到15.98%和1538%,普通小麦为12.38%,其蛋白质含量显著优于普通小麦。张帅等[24]测定了7份白粒青稞品种、6份黑粒青稞品种以及2份紫粒青稞品种的蛋白质含量,发现有色麦类品种中的蛋白质平均含量比白色青稞品种中的蛋白质平均含量高出了19.85%。综合分析以上研究发现,有色麦类作物中蛋白质与氨基酸含量及种类显著高于普通麦类作物。蛋白质和氨基酸作为人类生理机能中细胞、组织、器官中的重要组成成分,它具有供给生长、修补和更新所需营养等多种生理功能[25]。因此,人体为了满足自身的生长发育需要,保证摄入足够的构成人体蛋白质和多种氨基酸原料,有色麦类作物是一个很好的选择。
1.3 维生素
有色麦类作物中维生素的总体含量较高,在人体营养中起着重要作用。Lachman等[26]采用高效液相色谱-荧光法测定了彩粒小麦的生育甾醇含量,结果发现紫粒和蓝粒小麦品系中含有较高的β+γ维生素E。Guo等[27]通过研究紫小麦的营养特性,以1个白小麦品种作为对照,比较了7个紫小麦品系中的41种营养物质,结果发现紫小麦中所含的β+γ维生素E超过了对照的300%。研究表明,丰富的β+γ维生素E具有抗氧化、对抗自由基及抑制过氧化脂质生成等功能,被人体吸收后可有效祛除黄褐斑、抑制酪氨酸酶活性,达到减少黑色素生成的功效[27]。梁寒峭等[28]通过对黑青稞中的维生素种类进行检测,结果发现黑青稞中维生素种类多样,尤其是B族维生素较为突出,总量为9.53 mg/100 g,其中烟酸的含量最高,通常B族维生素参与人体新陈代谢中的关键代谢反应。可见,与白色麦类相比,有色麦类作物中维生素种类和含量较为丰富。有色麦类在补充人体所需的关键维生素、有效增强人体抗氧化能力以及参与人体内的关键代谢功能等方面发挥重要作用。
1.4 膳食纤维
在现代营养学中,膳食纤维被称为“第七大营养素”,它在预防肥胖、减缓心血管疾病、糖尿病、甚至癌症等现代“文明病”中发挥着重要作用[29-30]。安震[31]测定了24份蓝、紫色青稞籽粒的花青素、膳食纤维、β-葡聚糖和总黄酮等营养成分含量,结果发现供试青稞资源的膳食纤维含量在14.70~21.70 g/100 g,平均含量达到17.98 g/100 g,具有较高含量的膳食纤维组分。Rakha等[32]测定了黑小麦的总膳食纤维,含量为13%~16%,主要由阿拉伯糖基木聚糖(6.8%)、果聚糖(2.3%)、纤维素(2.1%)等成分组成。且黑小麦籽粒中的膳食纤维主要由可溶性多糖与水溶性纤维素源组成,含量是普通浅色小麦的2~3倍[33]。虽然这些有色麦类籽粒中膳食纤维含量不一,但普遍高于普通麦类作物,可以基本满足人体所需膳食纤维需要,这对于提高人体肠胃蠕动和生长代谢具有重要意义。
1.5 矿物微量元素
矿物微量元素对于人体内的酶类活性催化及生理功能具有重要意义。研究表明,彩色小麦中纯天然的微量元素以铁、锌、硒、碘为主,其含量超过目前国家正推行的“7+1”营养强化面粉中的微量元素的规定[34]。苏东民等[35]发现黑小麦的矿物质元素尤以硒、锰与碘的含量比较丰富。可见,有色麦类作物能够形成丰富的矿物微量元素,长期食用可有效地补充人体所需的大部分矿物微量元素。
2 有色麦类花青素的合成调控
2.1 花青素的种类及功能
花青素(anthocyanins)属于类黄酮化合物,是植物中的一组水溶性植物色素,赋予许多花、果实和叶片多种颜色,包括粉红色、红色、紫色和蓝色等[36]。花青素主要由一个3碳单位连结(C6-C3-C6)2个苯环构成[37]。目前已被报道的花青素种类约有100多种,以锦葵花青素、天竺葵花青素、飞燕草花青素、芍药花青素、牵牛花花青素和矢车菊花青素6种在植物内较为常见[38-39]。再者,自然界中花青素很少以游离态存在,通常是以结合单糖或低聚糖形成花色苷结构存在,在植物整个生长周期中扮演着重要角色,如减轻寒冷、耐干旱和抗紫外线照射等方面,以及吸引传粉昆虫(如蜜粉)和相关动物(如鸟类)进行授粉和种子传播等方面发挥重要作用[40]。同时,花青素在对人体保健方面也起着一定的作用,花青素具有抗衰老、抑菌、抗炎和保护视力等多種功效,能够降低动物机体内的氧化应激[41-42]和炎症[6]反应,还可通过血脑屏障送达大脑组织起到保护神经的作用。
2.2 与籽粒颜色相关的花青素合成调控机制
有色麦类作物籽粒颜色的多样性主要受到了花青素合成途径的影响,目前已被广泛证实。研究发现,有色麦类作物籽粒颜色如紫粒、蓝粒、红粒、黄粒、黑粒等主要与花青素合成过程中花青素产生途径、原花青素产生途径及鞣红(鞣酐)产生途径等代谢合成途径有关[43-45]。例如,小麦的黄色籽粒与原花青素的合成代谢途径有关,大麦的黑色籽粒与鞣红(鞣酐)产生合成代谢途径有关,小麦和大麦紫粒、蓝粒、红粒的形成主要与花青素产生途径有关[44-46]。同时,已有诸多研究表明花青素的合成途径主要是发生在内质网膜上,受到了结构基因、调节基因和转运因子相关的基因所调控[45]。其中,调节基因主要包含了R2R3-MYB、WD40和MYC家族的bHLH多药和有毒化合物排出家族(multidrug and toxic compound extrusion,简称MATE)、转运子基因包括谷胱甘肽转移酶(glutathioneS-transferases,简称GST)等[46-51]。
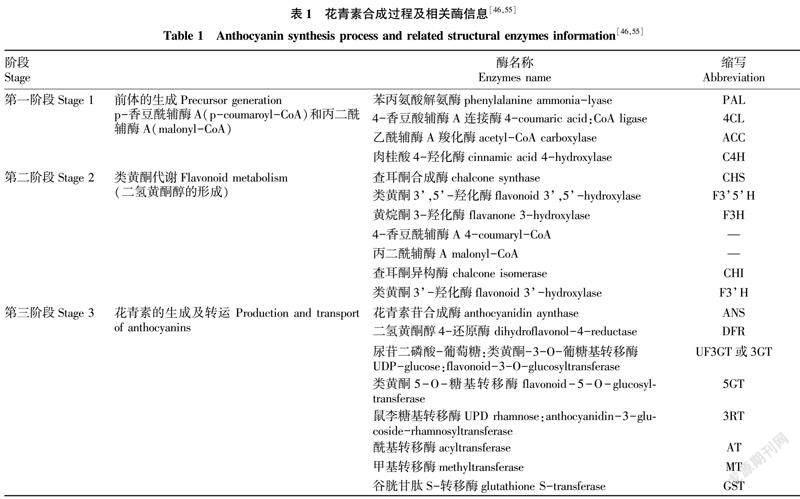
花青素通过细胞质中苯丙素和类黄酮途径合成的苯丙氨酸被转运到植物液泡中[52-53]。其生物的合成主要分为3个阶段:第一阶段为苯丙氨酸(phenylalanine)和乙酸(aceticacid)在一系列的催化转运酶的基础上催化转化合成为花青素直接前体;第二阶段为类黄酮合成代谢途径,从p-香豆酰辅酶A(p-coumaroyl-CoA)和丙二酰辅酶A(malonyl-CoA)开始,直到形成二氢黄酮醇;第三阶段为花青素的生成阶段,即二氢黄酮醇经过二氢黄酮醇4-还原酶(dihydroflavonol-4-reductase,DFR)的催化生成无色花色苷,再经过花色苷合成与转化途径的相关酶进行催化形成有色花色素[54-55]。最后,这些修饰的花色素被谷胱甘肽转移酶(GST)从细胞质中运输到液泡中[55]。此外,对于有各个阶段所涉及的酶或结构基因在不同阶段受到了相关酶或结构基因的调控,目前已报道调控籽粒颜色相关结构酶有19个(表1):第一阶段所涉及的酶主要包括了苯丙氨酸解氨酶(phenylalanine ammonia-lyase,PAL)、4-香豆酸辅酶A连接酶(4-coumaricacid:CoA ligase,4CL)等;第二阶段主要涉及的酶有查耳酮合成酶(chalcone synthase,CHS)、类黄酮3’,5’-羟化酶(flavonoid 3’,5’-hydroxylase)、黄烷酮3-羟化酶(flavanone 3-hydroxylase)等;
第三阶段涉及的酶有二氢黄酮醇4-还原酶(dihydroflavonol-4-reductase,DFR)、花青素苷合成酶(anthocyanidin aynthase,ANS)、尿苷二磷酸-葡萄糖等[55-57]。
花青素的合成、转运和积累还受到多种转录因子(如MYB、MYC、b HLH、b ZIP、WD40、WRKY等)的特异性调控[55,58]。这些转录因子通过与结构基因的协作方式共同调控花青素的生物合成[55]。它们通过识别结构基因启动子特定区域与启动子结合,从而达到促进或抑制花青素生物合成的作用,且不同植物花青素合成过程中转录因子调控的结构基因有所差异[58]。
3 麦类作物粒色基因定位研究进展
基因定位是现代分子育种的基础,基因定位的方法有多种,如体细胞杂交、原位杂交、连锁分析基因定位等,其中连锁分析基因定位分为初步定位和精细定位[59]。初步定位是将数量性状基因座定位在染色体上某个较大的区域内,为10~30 cM,可以定位到一个或多个基因座,往往不能确定该目标基因位点在基因组上的具体位置[59-60]。精细定位是在初步定位的基础上,针对目标基因位点所在区域,构建遗传背景相同的次级分离群体,在已构建的遗传图谱的基础上探索新的分子标记,对现有的图谱进行补充,不断增大分子标记的密度,将目标基因位点控制在一个更精确的區间内[59-60]。基因定位的工具包括多种分子标记技术,如SSR、SNP、In Del、RAPD、AFLP、SRAP、SCAR等,利用分子标记构建遗传连锁图谱及高密度遗传连锁图谱,从而将目标性状基因定位于染色体的相应位置,完成基因的定位[60]。目前在麦类作物中应用最为广泛的主要有RAPD、SSR、SNP标记等[61],用于基因定位的高通量测序技术主要包括转录组测序(RNA-seq)、代谢组测序、SLAF-seq(Specific-locus amplified fragment sequencing)、GBS-seq(Genotyping- by-sequence)等技术[62-65]。
在大麦籽粒颜色基因定位研究中,Lundqvist等[66]对紫大麦的自然突变体进行遗传分析,结果发现紫大麦受到了Pre1和Pre2 2个显性互补基因调控,2个基因分别位于2HL,距大麦中的六棱穗状花序基因(vrs1)和无原花色素基因(ant2)的遗传距离分别为17.1和11.2 cM。Lundqvist等[66]对黑粒大麦的粒色基因进行遗传性状分析,结果发现黑大麦主要由1对显性基因(Blp)调控,当显性基因(Blp1.b)发挥调控作用时所控制的黑大麦籽粒颜色较深,当显性基因(Blp1.mb)所发挥调控作用时黑大麦颜色中等,而当携带的显性基因(Blp1.g)发挥调控作用时黑大麦则显现出浅黑色或灰色。此外,Shoeva等[43]利用紫色、黑色和无色大麦近等基因系NILs(Near-isogenic lines)的遗传模型,借助微卫星基因分型技术将大麦中的紫粒基因(HvPLP)定位到了1H染色体上,将黑粒基因(HvBLP)定位到了2H染色体上。Yao等[44]利用紫粒与白粒青稞构建的DH(doubled haploid)系,利用简化基因组测序(GBS-seq)技术构建得到了青稞高密度的遗传图谱,在7H上获得了6个与青稞紫粒相关的位点。
在小麦植物籽粒颜色基因定位研究中,彭琴等[67]采用蓝粒小麦GLM170与4个白粒品种(贵农19、农麦30、中燕96-3和绵麦301)分别进行组配正反交得到其产生的F1和F2子代,用660K基因芯片和标记筛选对在贵农19×GLM1701组合中选取的F2纯蓝粒和纯白粒各40个单株叶片分别组建的颜色性状混池及亲本进行基因定位,结果发现控制小麦蓝色籽粒的基因位于4D染色体上,特异性标记Xgwm165-4D与目的基因的遗传距离仅为5.8 cM。Jiang等[68]通过转录组分析紫色果皮中花青素合成相关基因的分子机理,鉴定出TaPpm1和TaPpb1 2类转录因子为紫色果皮中花色苷合成的激活因子,其中TaPpm1为MYB 类调控蛋白,与之最近的多态性SSR标记为Xgwm44和Xgwm411;TaPp1为MYC类调控蛋白,与之最近的多态性SSR 标记为Xwmc632和Xgwm47。毕婵等[69]利用紫色籽粒小麦“农大3753”与非紫色籽粒突变体564进行杂交,得到F1子代和F2子代的分离群体,发现与紫粒性状紧密连锁的位于7D染色体上的SSR标记(Xbarc126),获得紫粒性状相关基因与7D-5-5和7D-4标记的遗传距离分别为4.2和2.6 cM。在2017年,西北农林科技大学Diddugodage Chamila Jeewani[70]使用 InfiniumTM i Select 90K芯片,结合F2群体粒色性状进行了QTL分析。结果表明,蓝色性状在4D染色体上2个SNP标记IWB18525和IWB16381之间有1个主效QTL位点。
目前利用基因定位技术对麦类作物籽粒颜色相关基因进行了挖掘,获得了控制大麦和小麦紫色、黑色和蓝色的候选区段(基因)。总体来看,对于麦类作物基因的挖掘首先利用不同粒色的父母本进行杂交,获得不同世代的群体,进行遗传分析,再借助包括SSR、SNP、RAPD等基因定位手段进行定位;也有通过转录组、代谢组测序等方法挖掘得到控制颜色相关基因。因此,随着现代分子生物学技术的不断发展,多种测序手段相结合的定位方法会进一步加快麦类作物粒色相关基因的挖掘与证实,从而推动有色麦类作物的新种质筛选与创制。
4 展望
麦类作物长期以来供给了人们的生活所需,且籽粒颜色相关的研究也已成为国内外麦类作物资源开发和研究的热点,随着人们生活水平的提高,开发以有色籽粒内的花青素、类黄酮等高营养物的研究越来越受到关注。目前,以分子标记技术为代表的基因作图技术已应用于遗传图谱构建、DNA指纹图谱与品种鉴定、分子标记辅助育种等方面[71]。通过对麦类籽粒颜色形成的相关基因进行的定位,极大地补充了人们对于粒色形成机制的认识。对于麦类作物的品种选育方法目前虽仍以常规遗传杂交育种方式为主,但随着现代分子生物学技术的发展,以分子标记技术为主的基因定位也逐渐融入诸多麦类新品种选育的过程中。并且,随着高通量测序技术的快速发展,转录组研究从以前的微阵列技术、SAGE(serial analysis of gene expression)技术等的低通量模式切换至RNA-seq等的高通量模式。例如Yao等[44]利用紫粒与白粒青稞构建的DH(doubled haploid)系,通过简化基因组测序(GBS-seq)技术构建得到了青稞高密度的遗传图谱,在7H上获得了6个与青稞紫粒相关的位点,在色青稞相关基因的挖掘上取得了又一步进展。目前被广泛应用的RFLP、PAPD等传统基因定位技术具有技术较为复杂、生产成本高且定位到的候选基因区段过大等劣势。利用分子标记结合高通量测序技术对于有色麦类作物颜色基因的挖掘与直接进行有色麦类籽粒颜色相关基因耦合的研究相对较少,特别是对于部分特殊生境条件下的有色麦类作物的研究更是少之甚少。因此,未来对于深度挖掘麦类籽粒颜色相关基因应与诸多分子标记技术结合高通量测序技术(如转录组测序RNA-seq、SLAF-seq、GBS-seq等)相结合,进行有色麦类籽粒颜色的精确定位。另外,未来随着现代分子生物学技术的发展,还可借助在麦类植物基因组水平与转录组水平上进行调控籽粒颜色相关基因的挖掘,并将其与基因编辑技术(CRISPR)相结合起来,直接通过基因编辑的方式对作物品种进行修饰和改造,以产生性状更为优良的新品种。
现今人们的物质需求日益增长,更加需要以高营养价值为目的的麦类育種改良与食品加工。然而,目前关于麦类籽粒颜色的研究仍主要集中在其形成的机制,重点关注如何提高麦类作物籽粒的产量等经济价值。虽已对有色麦类中的花色苷和多酚类物质为主的组成成分进行了研究,证实了有色麦类籽粒中花色苷和酚类种类较多具有较高的含量[72],但对于有色麦类作物中其他品质性状分析研究较少,例如蛋白质含量和质量、淀粉特性、浸出率、糖化力等,尤其是有色麦类作物中的维生素和抗氧化剂。通常,多种维生素和抗氧化剂在麦类作物中形成,能够显著增加麦类颜色作物的营养价值,以创造均衡的膳食平衡。已有诸多研究报道认为蓝色、紫色等特殊粒色小麦中的微量元素、氨基酸等营养物质种类多样且含量丰富[23-34]。因此,未来对于有色麦类作物进行育种和品种选育时,还可进一步鼓励对这些性状进行深入的研究,使之能够更好地应用于生产实际中。此外,目前虽已利用全套染色体形态性状标记,将有色基因定位到对应的染色体上,并将有色籽粒基因进行了遗传分析和定位,甚至克隆表达了部分控制麦类籽粒颜色的相关基因,明确了部分作用机理。但对于开发有价值的有色麦类作物的营养资源还远远不够,未来仍还需要进一步挖掘使用传统麦类作物(如小麦和大麦)和未充分利用的麦类作物(如燕麦),以筛选优化获得具有更高营养价值和适应性能力的品种,进一步实现人类饮食的多样性与营养性,这将是一项潜在的战略,在战略实现后可进一步确保粮食安全,保障粮食的可持续发展以及促进人类食品安全和饮食健康的均衡良性发展。
参考文献
[1] WANG X,ZHANG X C,HOU H X,et al.Metabolomics and gene expression analysis reveal the accumulation patterns of phenylpropanoids and flavonoids in different coloredgrain wheats(Triticum aestivum L.)[J].Food research international,2020,138:1-14.
[2] ABDELAAL E S M,HUCL P,RABALSKI I.Compositional and antioxidant properties of anthocyaninrich products prepared from purple wheat[J].Food chemistry,2018,254:13-19.
[3] 时玉晴,苏东民,陈志成.彩色小麦品质特性及其开发应用[J].粮食与油脂,2014,27(11):1-4.
[4] 华为,朱靖环,尚毅,等.有色大麦研究进展[J].植物遗传资源学报,2013,14(6):1020-1024.
[5] LI W D,BETA T.Flour and bread from black,purple,and bluecolored wheats[M]//PREEDY V R,WATSON R R,PATEL V B.Flour and breads and their fortification in health and disease prevention.Amsterdam:Elsevier,2011:59-67.
[6] SURIANO S,IANNUCCI A,CODIANNI P,et al.Carotenoids and tocols content in genotypes of colored barley[J].Journal of cereal science,2020,96:1-8.
[7] VARGA M,JJRT R,FNAD P,et al.Phenolic composition and antioxidant activity of colored oats[J].Food chemistry,2018,268:153-161.
[8] DE PASCUALTERESA ,SANCHEZBALLESTA M T.Anthocyanins:From plant to health[J].Phytochemistry reviews,2008,7(2):281-299.
[9] ZHANG Y Z,YIN L Q,HUANG L,et al.Composition,antioxidant activity,and neuroprotective effects of anthocyaninrich extract from purple highland barley bran and its promotion on autophagy[J/OL].Food chemistry,2021,339[2021-11-15].https://doi.org/10.1016/j.foodchem.2020.127849.
[10] 唐晓珍.彩粒小麦营养加工品质与色素研究[D].泰安:山东农业大学,2008.
[11] 杨晓梦,杜娟,李霞,等.大麦籽粒总花色苷含量的遗传变异及其与粒色的相关分析[J].核农学报,2020,34(2):273-280.
[12] 叶琳.彩色小麦营养成分及其色素基因表达模式的研究[D].西宁:青海师范大学,2018.
[13] 赵善仓,刘宾,赵领军,等.蓝、紫粒小麦籽粒花色苷组成分析[J].中国农业科学,2010,43(19):4072-4080.
[14] ABDELAAL E L S M,YOUNG J C,RABALSKI I.Anthocyanin composition in black,blue,pink,purple,and red cereal grains[J].Journal of agricultural and food chemistry,2006,54(13):4696-4704.
[15] HUGHES N M,CONNORS M K,GRACE M H,et al.The same anthocyanins served four different ways:Insights into anthocyanin structurefunction relationships from the wintergreen orchid,Tipularia discolor[J].Plant science,2021,303:1-7.
[16] ZHOU W,ZHANG T,ZHANG B L,et al.Addition of sucrose during the blueberry heating process is good or bad? Evaluating the changes of anthocyanins/anthocyanidins and the anticancer ability in HepG2 cells[J].Food research international,2018,107:509-517.
[17] DAI J,GUPTE A,GATES L,et al.A comprehensive study of anthocyanincontaining extracts from selected blackberry cultivars:Extraction methods,stability,anticancer properties and mechanisms[J].Food & chemical toxicology,2009,47(4):837-847.
[18] IGWE E O,CHARLTON K E,ROODENRYS S,et al.Anthocyaninrich plum juice reduces ambulatory blood pressure but not acute cognitive function in younger and older adults:A pilot crossover dosetiming study[J].Nutrition research,2017,47:28-43.
[19] 迪娜·吐爾洪,刘新莲,李建光.花色苷抗阿尔兹海默症的研究进展[J].中华中医药杂志,2019,34(4):1614-1617.
[20] 张永祥,国石磊,彭利沙,等.黑果腺肋花楸花色苷对人皮肤成纤维细胞紫外辐射损伤的保护作用[J].天然产物研究与开发,2016,28(7):1088-1092.
[21] 宋会歌,刘美艳,董楠,等.食品中花色苷降解机制研究进展[J].食品科学,2011,32(13):355-359.
[22] 谢景梅,马永甫,胡雪琴,等.蓝、紫粒小麦籽粒蛋白营养及抗氧化性研究[J].西南师范大学学报(自然科学版),2010,35(3):197-203.
[23] 袁园园,徐晓琳,李将,等.紫、蓝粒小麦籽粒营养成分分析与鉴定[J].农业科技通讯,2018(6):111-113.
[24] 張帅,吴昆仑,姚晓华,等.不同粒色青稞营养品质与抗氧化活性物质差异性分析[J].青海大学学报,2017,35(2):19-27.
[25] 叶忠孝.蛋白质的十大生理功能[J].中国保健食品,2011(7):37.
[26] LACHMAN J,HEJTMNKOV A,ORSK M,et al.Tocotrienols and tocopherols in coloredgrain wheat,tritordeum and barley[J].Food chemistry,2018,240:725-735.
[27] GUO Z F,ZHANG Z B,XU P,et al.Analysis of nutrient composition of purple wheat[J].Cereal research communications,2013,41(2):293-303.
[28] 梁寒峭,李金霞,陈建国,等.黑青稞营养成分的检测与分析[J].食品与发酵工业,2016,42(1):180-182,188.
[29] 章中.膳食纤维在慢性病防治中的作用[J].农业科学研究,2010,31(3):75-77.
[30] HARRIS P J,FERGUSON L R.Wheat and rice dietary fiber in colorectal cancer prevention and the maintenance of health[M]//WATSON R R,PREEDY V R,ZIBADI S.Wheat and rice in disease prevention and health.Amsterdam:Elsevier,2014:201-210.
[31] 安震.西藏日喀则市蓝、紫色青稞4项营养成分差异性分析[J].大麦与谷类科学,2020,37(4):30-33.
[32] RAKHA A,MAN P,ANDERSSON R.Dietary fiber in triticale grain:Variation in content,composition,and molecular weight distribution of extractable components[J].Journal of cereal science,2011,54(3):324-331.
[33] ZHANG M W,GUO B J.Nutritional functions and exploitation of black cereal and oil food resources[J].Journal of South China Normal University(natural science edition),2001(2):114-120.
[34] 魏壹纯,陈志成.彩色小麦营养组分及应用的研究进展[J].粮食与食品工业,2015,22(2):53-56,60.
[35] 苏东民,齐兵建,赵仁勇,等.漯珍一号黑小麦营养成分的初步评价[J].粮食与饲料工业,2000(8):1-2.
[36] ZHANG D W,LIU L L,ZHOU D G,et al.Genomewide identification and expression analysis of anthocyanin biosynthetic genes in Brassica juncea[J].Journal of integrative agriculture,2020,19(5):1250-1260.
[37] JAAKOLA L.New insights into the regulation of anthocyanin biosynthesis in fruits[J].Trends in plant science,2013,18(9):477-483.
[38] ANDERSEN M J,JORDHEIM M.Basic anthocyanin chemistry and dietary sources[M]//WALLACE T C,GIUSTI M M.Anthocyanins in health and disease.Boca Raton,FL:CRC Press,2013:13-89.
[39] SAKATA K,SAITO N,HONDA T.Ab initio study of molecular structures and excited states in anthocyanidins[J].Tetrahedron,2006,62(15):3721-3731.
[40] WINKELSHIRLEY B.Flavonoid biosynthesis.A colorful model for genetics,biochemistry,cell biology,and biotechnology[J].Plant physiology,2001,126(2):485-493.
[41] TANAKA Y,OHMIYA A.Seeing is believing:Engineering anthocyanin and carotenoid biosynthetic pathways[J].Current opinion in biotechnology,2008,19(2):190-197.
[42] CAO N,FU Y H,HE J H.Mechanical properties of gelatin films crosslinked,respectively,by ferulic acid and tannin acid[J].Food hydrocolloids,2007,21(4):575-584.
[43] SHOEVA O Y,MOCK H P,KUKOEVA T V,et al.Regulation of the flavonoid biosynthesis pathway genes in purple and black grains of Hordeum vulgare[J].PLoS One,2016,11(10):1-16.
[44] YAO X H,WU K L,YAO Y H,et al.Construction of a highdensity genetic map:Genotyping by sequencing(GBS)to map purple seed coat color(Psc)in hulless barley[J].Hereditas,2018,155(1):1-11.
[45] 苏乐平,姚晓华,吴昆仑,等.大麦(青稞)籽粒颜色相关研究进展[J].江苏农业科学,2019,47(18):70-74.
[46] 赵启明,李范,李萍.花青素生物合成关键酶的研究进展[J].生物技术通报,2012(12):25-32.
[47] 王艳花.大黄油菜粒色性状候选基因的定位克隆及功能分析[D].西宁:青海大学,2017.
[48] 闫瑞霞.紫山药花青素合成积累相关基因分子特征分析[D].南京:南京农业大学,2014.
[49] 石晓芳.甘薯花青素合成途径基因表达差异的研究[D].重庆:西南大学,2013.
[50] 亓燕红.彩色玉米籽粒中参与花青素合成基因的表达分析[D].泰安:山东农业大学,2011.
[51] 邹丽秋,王彩霞,匡雪君,等.黄酮类化合物合成途径及合成生物学研究进展[J].中国中药杂志,2016,41(22):4124-4128.
[52] DOONER H K,ROBBINS T P,JORGENSEN R A.Genetic and developmental control of anthocyanin biosynthesis[J].Annual review of genetics,1991,25:173-199.
[53] JEONG S T,GOTOYAMAMOTO N,KOBAYASHI S,et al.Effects of plant hormones and shading on the accumulation of anthocyanins and the expression of anthocyanin biosynthetic genes in grape berry skins[J].Plant science,2004,167(2):247-252.
[54] ABDELAAL E L S M,YOUNG J C,RABALSKI I.Anthocyanin composition in black,blue,pink,purple,and red cereal grains[J].Journal of agricultural and food chemistry,2006,54(13):4696-4704.
[55] 梁立軍,杨祎辰,王二欢,等.植物花青素生物合成与调控研究进展[J].安徽农业科学,2018,46(21):18-24.
[56] KOES R,VERWEIJ W,QUATTROCCHIO F.Flavonoids:A colorful model for the regulation and evolution of biochemical pathways[J].Trends in plant science,2005,10(5):236-242.
[57] XIA D,ZHOU H,WANG Y P,et al.How rice organs are colored:The genetic basis of anthocyanin biosynthesis in rice[J].The crop journal,2021,9(3):598-608.
[58] WANG Y C,CHEN S,YU O.Metabolic engineering of flavonoids in plants and microorganisms[J].Applied microbiology and biotechnology,2011,91(4):949-956.
[59] 田玉,马春红,宋丽华,等.数量遗传性状基因定位方法研究进展[J].河北农业科学,2021,25(5):88-91,97.
[60] 周萌萌,王佳楠,田丽波,等.葫芦科作物重要性状基因定位研究进展[J].热带作物学报,2018,39(3):606-613.
[61] 周喜旺,刘鸿燕,王娜,等.DNA分子标记技术在小麦遗传育种中的应用综述[J].甘肃农业科技,2017(5):64-68.
[62] 余其,王振波,沈真辉,等.分子标记技术及其在大麦遗传育种中的应用[J].大麦与谷类科学,2016,33(2):1-5.
[63] 张艳俊.基于转录组测序的小麦TaNAC069基因抗叶锈性分析与功能解析[D].保定:河北农业大学,2020.
[64] 孙亚林,王直新,季群,等.基于SLAF-seq技术开发芋肉纤维颜色性状的分子标记[J].中国蔬菜,2021(6):49-54.
[65] 张学军,杨文莉,张永兵,等.采用GBS-seq技术构建甜瓜高密度遗传图谱[J].新疆农业科学,2019,56(10):1828-1838.
[66] LUNDQVIST U,FRANCKOWIAK J,KONISHI T.New and revised descriptions of barley genes[J].Barley Genet Newsl,1996,26:22-43.
[67] 彭琴,周军,徐如宏,等.小麦种质GLM 1701蓝粒性状的遗传定位分析[J].种子,2020,39(12):25-31.
[68] JIANG W H,LIU T X,NAN W Z,et al.Two transcription factors TaPpm1 and TaPpb1 coregulate anthocyanin biosynthesis in purple pericarps of wheat[J].Journal of experimental botany,2018,69(10):2555-2567.
[69] 毕婵,尤明山.小麦紫色籽粒性状的基因定位及其遗传分析[C]//中国作物学会.第十届全国小麦基因组学及分子育种大会摘要集.北京:中国作物学会,2019:231.
[70] DIDDUGODAGE CHAMILA JEEWANI.蓝粒小麦中花青素生物合成基因的基因定位及转录组分析[D].杨凌:西北农林科技大学,2017.
[71] 徐丽芳,陈吉炎,罗光明.分子标记技术及其在植物育种中的应用[J].食品与药品,2007,9(10):43-46.
[72] BETA T,QIU Y,LIU Q,et al.Purple wheat(Triticum sp.)seeds:Phenolic composition and antioxidant properties[M]//PREEDY V R,WATSON R R,PATEL V B.Nuts and seeds in health and disease prevention.California,USA:Academic Press,2020:103-125.
猜你喜欢
山东青年报·教育周刊学生版上半月(2023年2期)2023-06-06
北方农业学报(2019年6期)2019-02-21
天然产物研究与开发(2018年11期)2018-11-30
热带作物学报(2018年12期)2018-05-14
天然产物研究与开发(2018年3期)2018-05-07
天然产物研究与开发(2018年1期)2018-02-02
中成药(2017年5期)2017-06-13
中成药(2016年8期)2016-05-17
中国康复理论与实践(2015年10期)2015-12-24
食品科学(2013年24期)2013-03-11
