
不同光强和光周期对山药珠芽生长发育的影响
2023-05-26 01:46:14李锦超刘淑祺师晨玮李跃凡贾凯旋刘晓清杨太新葛淑俊
种子 2023年3期
李锦超, 刘淑祺, 师晨玮, 李跃凡, 贾凯旋, 刘晓清, 杨太新, 葛淑俊
(河北农业大学农学院/教育部华北作物种质资源研究与利用重点实验室, 河北 保定 071000)
山药为薯蓣科多年生缠绕性藤本薯蓣(DioscoreaopposataThunb.),地下根茎具有重要的食用与药用价值[1],药用历史始载于《神农本草经》,列为上品,富含皂苷、黏液质、糖蛋白等多种有效成分,具有降血糖、降血脂、延缓衰老等药理作用[2]。山药在北方几乎不结种子,无法利用种子来进行更新[3]。山药珠芽,在薯蓣中常叫做“零余子”,俗称“山药豆”[4],为变态腋生块茎,具有与根茎相似的成分与功效,珠芽成熟后能自主发育为新的植株,是山药种苗繁育、脱毒快繁和遗传转化研究的重要组织材料[5]。
山药珠芽是由多个副芽分化而成的一种腋生复合结构,具有多个分生区和可以生根或出芽的疣突。珠芽解剖学结构从外到内由表皮、薄壁组织、初生加厚分生组织、内生薄壁组织和散生在其中的有限外韧维管束组成[6]。其形成和发育与蔗糖、淀粉的代谢以及激素含量的变化有重要关系[7]。当山药茎蔓枯黄时,珠芽中糖分和干物质积累完全,表皮完全变褐,发育成熟[8]。在离体条件下珠芽诱导和发育受激素、蔗糖浓度和培养条件等因素影响。李明军等[9]用怀山药带芽茎段作为外植体,在含有60 g/L 蔗糖的培养基和完全黑暗的培养条件中能诱导珠芽产生;彭晓英等[10]将盾叶薯蓣带顶芽茎段竖插于MS+4.0 mg/L 6-BA+1.0 mg/L KT+6%蔗糖培养基中,光照条件为1 500~2 000 lx,14 h/d,建立了珠芽离体诱导体系;将脚板薯脱毒试管苗双节茎段接种在MS+2.0 mg/L 6-BA+0.5 mg/L NAA+8%蔗糖培养基中,8 h/d光照下,珠芽离体诱导率可达100%,且数量、鲜重和直径均高于其他光照处理[11]。由此可见,高浓度的蔗糖是离体茎段形成珠芽的关键因素。但光强和光周期对山药珠芽形态发生的影响尚未有系统研究。
本试验以河北省道地山药小白嘴为材料,以带芽茎段为外植体,研究离体条件下光照强度和光周期对珠芽诱导和生长发育的影响,为解析山药珠芽发育机理和种苗高效快繁提供基础。
1 材料与方法
1.1 实验材料
将小白嘴山药根茎切段栽种于育苗盒中,培养至幼苗嫩茎具7~8个节段备用。
珠芽诱导选用MS固体培养基,蔗糖浓度6%,琼脂浓度0.8%,pH值5.8~6.0。121 ℃高温高压灭菌20 min备用。
1.2 光环境设置
对实验材料进行光照强度和光周期处理。其中光照强度设3个处理,光合有效辐射(Photosynthetically active radiation,PAR)分别为20 μmol/(m2·s)、95 μmol/(m2·s)、195 μmol/(m2·s)。在每个光强下分别设置4 h/d、8 h/d、10 h/d、12 h/d、16 h/d等5个光照时长,共15个光强和光周期组合(表1)。

表1 光强和光周期的处理组合Table 1 Processing combinations of light intensity and photoperiod
1.3 接种及调查
剪取山药植株带芽嫩茎,流水冲洗30 min,无菌水冲洗3遍,用75%酒精消毒30 s,0.1%氯化汞溶液消毒8 min,无菌水冲洗5次。接种到培养基中,每瓶接3段,每处理15瓶,每天光照10 h,温度(25±1)℃。待珠芽肉眼可见时(约15 d)转移到上述光调控条件下。在处理0 d(D 0)、10 d(D 10)、20 d(D 20)、30 d(D 30)后,调查以下项目:分枝数;叶面积及叶色;茎节数;叶腋数;珠芽数量;珠芽着生位置;珠芽长度;珠芽须根数和须根长;珠芽表皮颜色变褐面积;珠芽出芽率。其中,珠芽着生位置将接种外植体所含节定为初节(0),腋芽新生节根据生长次序依次定为第1、2、3、4节。

注:A为接种1 d茎段;B~C为培养15 d的茎段;B为长新生茎段外植体;C为未长新生茎段;D~G为由左至右第一排为处理1~5,第二排为处理6~10,第三排为处理11~15;D~E为光调控10 d;F~H为光调控20 d;I~G为光调控30 d;D、F、I为带叶片植株;E、H、G为剪除叶片植株。比例尺为1 cm。图1 山药植株的生长形态变化Fig.1 Morphological changes of yam plantlets
1.4 数据分析
数据统计及图表制作使用Microsoft Office Excel 2016软件,采用IBM SPSS Statistics 25.0统计软件进行分析,用新复极差测验法(Duncan test)进行处理间差异显著性比较。不同小写字母之间表示显著性差异(p<0.05),同一列中不同字母代表差异显著。
2 结果与分析
2.1 光强和光周期对山药外植体生长状态的影响
将消毒后的山药带芽茎段接种到培养基上,茎节处芽点会继续分化发育,部分腋芽生长迅速,长出新生茎段,新叶呈绿色,约15 d后在叶腋处有乳白色珠芽着生,有不定根迅速形成并伸长(图1 B);有些腋芽则不再伸长生长,仅在原始茎节叶腋处分化形成乳白色珠芽,同样有不定根迅速形成并伸长(图1 C)。
带有珠芽的茎段转移到不同光强和光周期组合条件下继续培养,叶片和分枝等逐渐发生变化。弱光[PAR 20 μmol/(m2·s)]下培养,新生叶片一直呈深绿色,中度光强[PAR 95 μmol/(m2·s)]下出现程度较弱的黄化现象,而在强光条件[PAR 195 μmol/(m2·s)]下培养叶片黄化极明显,且随着培养时间和光照时数的延长黄化现象更严重,20 d后16 h/d光照下会出现叶片枯萎,30 d后在12 h光照下造成叶片枯死。表明山药叶片在强光条件下随着培养时间的增长而出现黄化现象,不利于外植体的生长,产生光胁迫(图1)。
光照强度对离体培养的山药茎段分枝数、茎节数和叶腋数没有显著影响,处理前后均为1、2个和4个,而光照时间影响较大,与较短日照时数相比,8 h和16 h的光照使茎、叶数量产生显著差异,尤其在16 h/d强光照射下分枝生长受到抑制,分枝数、茎节数和叶腋数(分别为0.93、2.23、3.80)均显著低于其他处理。分析表明,15 d内山药茎段的茎、叶分化已经定型。因此,各处理间差异不显著,而光强和光周期的变化影响了山药叶片的生长状态,长时间强光照射会抑制植株生长,进一步对珠芽的生长产生影响。
2.2 光调控对山药珠芽的影响
2.2.1光强和光周期对山药珠芽着生位置的影响
经过统计分析,山药珠芽的着生位置在各处理中未产生较大差异,最初珠芽集中着生在初叶节和新生枝的第1节,平均占比47%和43%,随着处理时间的推移,第2节和第3节处的珠芽开始生长,第4节仅在个别生长健壮的植株中少有珠芽着生(图2)。但在强光处理4、5和中等光强处理10中随着光照时间的增加,初叶节珠芽占比大于50%,说明长时间强光照射抑制了山药新生枝条及其附生珠芽的生长。
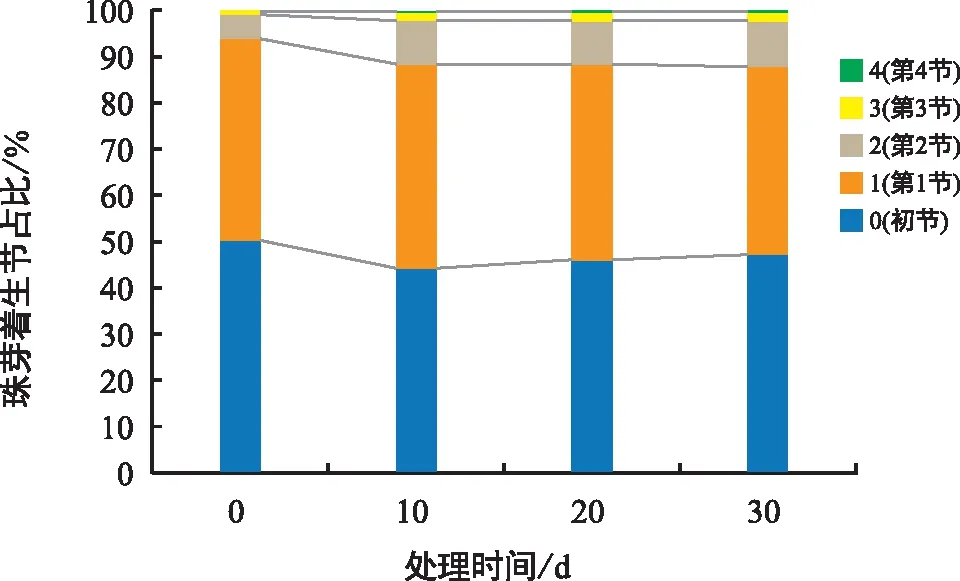
图2 山药珠芽着生位置统计Fig.2 Statistics of bubil position of yam
2.2.2光强和光周期对山药珠芽数量的影响
随着在不同处理下山药生长状态的变化,珠芽数量也随之变化,0~10 d是珠芽数量增长最快的阶段,在处理10 d后数量趋于稳定,平均每株4个珠芽,之后的20 d增长速度减缓。在强光和中等光强下,珠芽在光照8~16 h内随着时长的增加数量有所减少,而在4 h光照下分别低于8 h、8~12 h的珠芽数量;弱光条件下趋势相反,4~12 h光照下珠芽数量呈增长趋势,而16 h处理低于10~12 h光照的数量(表2)。分析可得,山药珠芽生长数量与植株的叶腋数呈极显著正相关,而山药珠芽的起始发育与光合有效辐射的积累有关,4 h/d的195 μmol/(m2·s)光强照射基本满足珠芽的发育需求,8 h时长下珠芽数量最多,而在20 μmol/(m2·s)光强下需要10 h光照时长才能满足,并在12 h的光照条件积累下达到最高值。

表2 光照处理中山药珠芽数量的变化Table 2 The change of the number of bulbil in each light treatment
2.2.3光强和光周期对山药珠芽长度的影响
图3表明,山药珠芽在处理7、8中30 d平均长度超过4 mm,显著优于其他处理,弱光下平均长度也能达到3.5 mm左右,而在强光下珠芽整体长度仅达到3 mm,其生长显著受到抑制。山药珠芽随着处理时间的延长,伸长生长量亦有所差别,不同光照条件下达到生长高峰的时间不同,强光处理整体抑制了珠芽的生长,处理前期0~10 d生长受阻,中期10~20 d适应后生长较快,后期20~30 d生长迟缓滞缓;弱光下前期生长迅速,中后期生长速度减缓;中等光强下珠芽生长量显著优于其他处理(图3)。通过分析光周期和珠芽生长量的关系发现,在各光强下,4 h/d光照均不能满足珠芽快速生长的需要,并且16 h/d的光照不同程度的抑制生长,而每天8 h和10 h的光照时长适宜珠芽的生长。

图3 光照处理中山药珠芽长度变化Fig.3 The length of yam bulbil changed under each treatment
2.2.4光强和光周期对山药珠芽表皮颜色的影响
在不同光照处理中培养发现,珠芽在最初萌发时为乳白色,随着培养时间的延长,在处理10 d后中等以上光强均会促使珠芽50%以上的表皮变成褐色,强光下的变化稍早于中等光强,20 d后达到70%以上,30 d后达90%以上。在弱光条件下,表皮褐化程度和速度与光照时间的增加呈正相关,4 h/d光照使珠芽颜色一直处于显著较低的褐化程度。通过分别分析光强和光周期对表皮颜色的影响发现,弱光条件下珠芽褐化程度一直低于同期强光照射,并且中等光强与强光变化程度趋于一致;在不同光周期下,4 h/d光照有利于保持珠芽较浅的表皮颜色,16 h/d光照显著促进表皮变褐(图4)。分析表明,山药珠芽表皮对光反应极为敏感,在弱光条件下亦产生褐化,而褐化程度标志着珠芽成熟度的变化,减少光照辐射会减缓褐变速度,使珠芽能够持续生长,为获得更高质量的珠芽提供可能性。

图4 光强和光周期对山药珠芽表皮颜色的影响Fig.4 The effect of light intensity and photoperiod on the epidermis color of bulbil
2.2.5光强和光周期对山药珠芽须根生长的影响
珠芽萌发时伴随着1~2条须根的生长,随着处理时间的延长,弱光条件下,每个珠芽根数达到平均6条以上,平均根长17 mm左右(表3);强光明显抑制根的萌发和生长,随着光照时长的增加,根数和根长下降趋势明显(图5);中等光强配合10 h以下光照时间有利于根的生长,16 h抑制根的生长。根的生长决定了珠芽吸收养分的多少,从而影响整体的生长趋势。综合来看,弱光和短日照条件是促进山药珠芽须根生长的有利因素。

图5 光照处理0~30 d山药珠芽不定根长度变化Fig.5 The length of bulbil adventitious roots varied from 0 to 30 days under light treatments
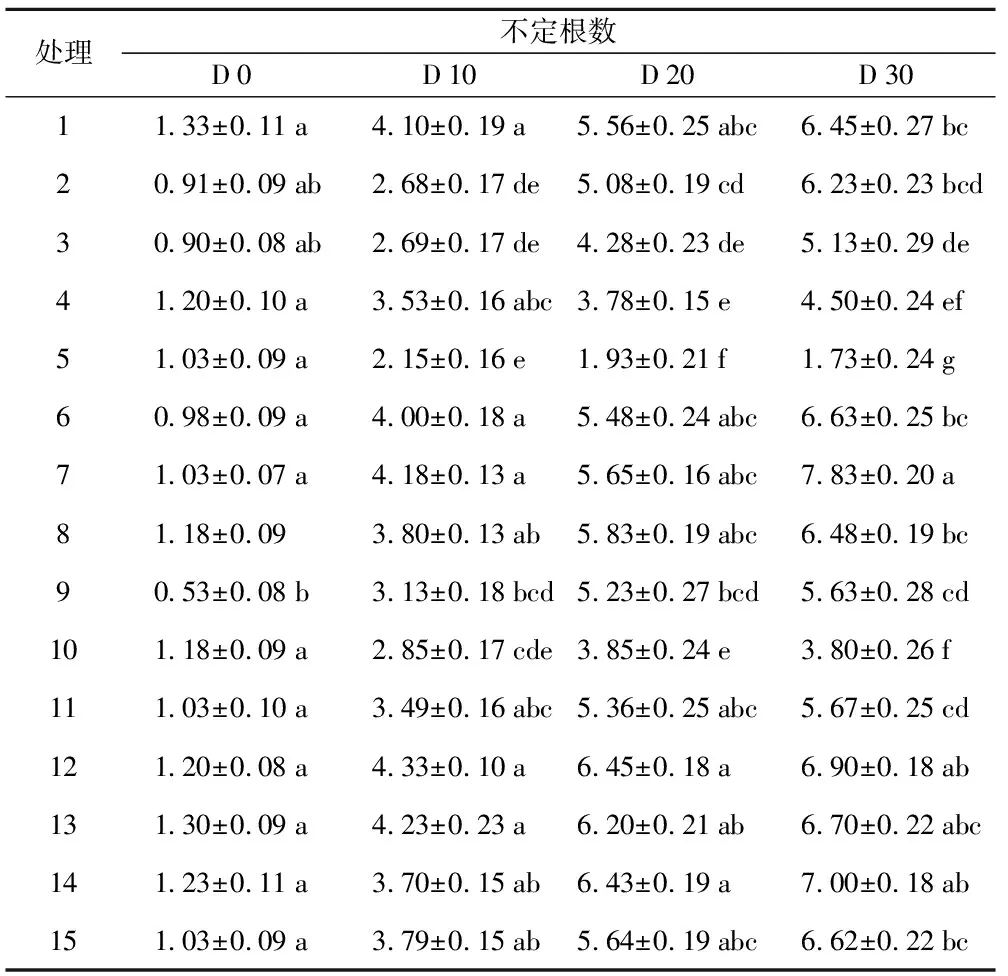
表3 光照处理中山药珠芽不定根数量变化Table 3 Adventitious root number changes of yam bulbil under light treatmentt
2.2.6光强和光周期对山药珠芽发芽率的影响
山药组织培养体系中,珠芽是诱导再生植株的理想材料。本研究发现,山药茎段在培养25 d后珠芽在不同光照条件下均有出芽现象,弱光下发芽较早,在处理10 d后平均发芽率达到72.20%,而强光明显抑制出芽,处理后10 d发芽率仅39.17%,处理30 d后发芽率约50%,中等光强发芽率达60%~70%,弱光则提升发芽率达83.82%左右,光周期对发芽率的影响不显著(图6)。分析表明,光强是影响山药珠芽隐芽萌发的重要因素,受光周期的调控较弱。

图6 光强和光周期对山药珠芽发芽率的影响Fig.6 Effects of light intensity and photoperiod on the rate of bulbil shoots
3 讨 论
光照是绿色植物生长的重要环境条件之一,山药珠芽作为一种变态茎,且处于光照环境中,除了生长方向为向重力性生长以外,似乎沿袭了茎在光照环境中的习性。在植物学中,山药珠芽和地下根茎均起到贮藏作用,作为“库”器官吸收转化“源”器官输送的光合产物,其自身不能进行光合作用。在田间由于各种种植系统的不同经常发生荫蔽现象,导致光照条件不匀,田间试验发现遮荫会降低箭叶兰芋(Xanthosomasagittifolium)、甘薯 (Ipomoeabatatas)、山药(Dioscoreaesculenta)、木薯 (Manihotesculenta) 和香芋 (Colocasiaesculenta)等主要热带块根和块茎作物的下表皮气孔密度,导致产生表面积更大但更薄的叶子[12],在山药外植体培养过程中,长时间强光照射导致叶片加速衰老,而弱光下枝叶茂密,具有更多的分枝和茎节。Hendrik等[13]发现,许多植物表型特征随着日常光积分(DLI)以指数方式增加,比如本研究中外植体根、茎、叶等可在光调控下产生明显的差异。山药对光周期的微小变化非常敏感,Marcos等[14]评估辐射利用效率(RUE)随种植日期的变化及其对水山药(DioscoreaalataL.)生长和产量的影响发现,受作物发育和光周期控制的源库相互作用,最大RUE与光周期呈负相关。Vaillant等[15]研究了光周期对山药幼苗发育的影响,发现地下块茎仅在12 h光周期下诱导形成和发育,相反,16 h的光周期抑制了块茎的形成并刺激了藤蔓和叶子的生长。Yasunori等[16]研究了光周期对山药(DioscoreaoppositaThunb. cv. Ichoimo)块茎发育的影响,发现长日光周期促进了枝叶的发育,相比之下,块茎受到16 h和24 h光周期的严重抑制,本研究中珠芽的生长同样受到16小时强光照射的抑制。
由于珠芽是一些植物在特殊生境下为保障繁衍而进化出的器官,因此植物珠芽的形成与环境因子也有一定的关系。珠芽繁殖是特基拉龙舌兰对弱光环境的适应性选择,在弱光条件下,龙舌兰的花茎上产生混合分生组织,在小苞片处萌生珠芽[17]。不同植物的珠芽形成对光周期有不同的要求,珠芽艾麻和楼梯草在短日照条件下形成珠芽,珠芽景天在长日照条件下形成珠芽。短日照、弱光环境能促进台闽苣苔产生相对更多的珠芽[18]。由此可见,光照会显著影响一些植物珠芽的形成,不同植物对光强和光周期有不同的适应性,山药珠芽的伸长也受到光环境的调控从而产生适应性生长。
光照是植物组织培养体系中的重要条件之一,自1955年首次报道在向日葵等组织培养中引进光因子以来[19],有关光调控在植物组织培养中的应用研究已引起人们关注。在前人的研究中,短波光在较低强度下对烟草芽发生有促进作用,而长波光需要在较高的光强下才能促进芽的发生[20]。不同植物对光的接受能力不同,兰花组培苗在中等光强[270 μmol/(m2·s)]下类胡萝卜素含量高于低光强[175 μmol/(m2·s)]或高光强[450 μmol/(m2·s)],高光强下光合色素会加速分解而使其含量降低,而类胡萝卜素可以吸收过剩的光子能量保护叶绿素不被氧化,减少光抑制[21],但人参组培苗在60 μmol/(m2·s)条件下叶绿素和类胡萝卜素含量最高[22]。延长光照时间可促进地黄组培苗的生长[23]。不同的植物在离体培养中对光的需求也不同,光强、光质和光照时间的不同对外植体的生长产生不同的影响,根据培养目的不同创造合适的光环境能达到事半功倍的效果。同样,在本研究中通过光环境调控发现,弱光和短日照条件促进山药珠芽持续伸长、出芽和根的生长,而强光和长日照条件使珠芽表皮颜色褐化加快,促进成熟,抑制芽和根的生长。
本研究利用山药带芽茎段为外植体,研究在培养过程中光强和光周期对茎、叶及珠芽伸长生长的影响。结果表明,长时间强光照射会抑制茎叶生长和珠芽的伸长;珠芽的发育与光合有效辐射的积累有关;在各光强下8 h和10 h光周期适宜珠芽的生长;珠芽表皮对光反应极为敏感,在弱光条件下亦产生褐化;弱光和短日照条件促进珠芽根的生长;光强是影响珠芽出芽的重要因素。综合来看,如培养目的为获得更高质量的种苗即促进珠芽的伸长生长可选择95 μmol/(m2·s)光照8 h或10 h,抑制生根和萌发可采用195 μmol/(m2·s)光照12 h或16 h,提高萌发率和促进须根生长可选用20 μmol/(m2·s)照射4~12 h,为山药种质研究和快繁体系的优化提供参考。
猜你喜欢
贵州农业科学(2023年10期)2023-10-27 00:05:42
遗传(2023年9期)2023-09-25 09:31:44
农业科技通讯(2023年1期)2023-02-12 07:08:50
草地学报(2021年6期)2021-07-05 03:50:08
特种经济动植物(2021年2期)2021-02-25 05:47:34
长江丛刊(2020年13期)2020-11-19 02:42:32
少儿科学周刊·少年版(2017年7期)2017-09-29 08:02:41
少儿科学周刊·儿童版(2017年7期)2017-09-29 02:22:58
机电信息(2014年17期)2014-02-27 15:53:01
食品科学(2013年17期)2013-03-11 18:26:53
