
不同种源的毛建草珠芽形成比较研究
2021-07-05 03:50:08田旭平郭庆浩孙鸣屿
草地学报 2021年6期
田旭平, 郭庆浩, 孙鸣屿, 李 倩, 路 通
(山西农业大学林学院, 山西 太谷 030801)
珠芽是植物在长期的物种进化中,为更好地适应不同的生境而形成的保护自我、繁衍后代的“胎生器官”,能较好的保留母体特征,较少变异和退化,是一种特殊器官[1]。高海拔地区的植物周期性的开花结实是一种极具风险的繁殖方式,为了避免种群更新失败,高山植物采用多种营养繁殖方式,这种方式的重要性常随海拔升高而增加[1]。很多高山植物在花期会产生营养芽(珠芽或吸芽),珠芽为地上茎叶腋间自然发生的短缩、肥厚呈莲座状的短枝,当珠芽长到一定大小时,其下部能自然生根,从母株上剥离能栽植成功。珠芽的落地生根,可以使得母株迅速传播并得以广泛分布,具有扩大种群数量并占据较多生长空间的潜在能力[1-2]。
毛建草(又名岩青兰,DracocephalumrupestreHance)是唇形科(Labiatae)青兰属(Dracocephalum)毛建草系多年生草本植物,主要生于海拔650~2 000 m(青海达2 650~3 100 m)的高山草原、草坡或疏林下向阳处[3],中国辽宁、内蒙古、河北、山西、青海等为其主要产地,在这些省份的一些地方,常用毛建草地上部制作具有地方风味的茶叶。在栽培毛建草时,发现一些毛建草植株在开完花后,花茎的叶腋间会产生一些珠芽或新的花茎。目前尚未见到有关毛建草珠芽的报道。为了了解毛建草珠芽的繁殖特性,本文比较了采集地与人工栽培地珠芽的形成差异及扦插生根情况,以期为研究毛建草种群繁殖提供科学基础,为气候变化对毛建草的生物进化影响提供理论依据。
1 材料与方法
1.1 珠芽形成的观察
在2017年从山西省宁武县芦芽山大洼村周围(111°59′29″ E,38°47′30″ N,平均海拔2 360 m)和娄烦县云顶山顶(111°38′14″ E,37°53′14″ N,平均海拔2 708 m)的草地采集毛建草植株;在2018年分别从交城关帝山(112°6′34″ E,37°37′15″ N,平均海拔2 216 m)、浑源恒山(113°43′ 56″ E,39°40′25″ N,平均海拔1 280 m)、辽宁朝阳县苏家店村半截沟山(120°13′ 56″ E,41°16′ 46″ N,平均海拔380 m)采集。在5个种源地采集的毛建草都种植在山西农业大学林业站试验地(112°35′19″ E,37°26′7″ N,海拔780 m)。
在2018—2020年对种植的毛建草统计产生珠芽的植株数量,并观察珠芽的形成发育过程。在2019年对种植的毛建草植株产生的珠芽数量进行比较,对珠芽最多的和最少的材料进行了计量。在2018年9月和2019年10月分别对芦芽山、关帝山和恒山采集地的毛建草珠芽着生情况进行调查。
1.2 珠芽原位生根观察
在2018年10月20日,对种植的源自芦芽山的毛建草珠芽在母株周围的原位生根情况进行调查,统计生根的珠芽数量,并拍照记录珠芽原位生根状态。
1.3 珠芽扦插试验
在2019年8月22日,根据珠芽球状木质部直径及叶片长度的不同,将源自芦芽山的毛建草的珠芽分为大中小3类。大珠芽:直径0.5 cm以上,叶长6 cm以上;中珠芽:直径0.3~0.5 cm以内,叶长4~6 cm以内;小珠芽:直径0.3~0.5 cm以内,叶长4 cm以内。大中小珠芽各扦插150 颗,基质采用珍珠岩、蛭石、有机质按1∶1∶1的比例配制,将珠芽基部膨大的球状木质部埋进基质中,露天养护,用喷壶喷水保持湿润。
在10月10日检查珠芽存活率,有绿叶的认为成活,其他状况认为没有成活,同时随机挖30 棵珠芽检查其生根情况。对未生根的珠芽在11月10日低温来临前再次检查生根情况。
2 结果和分析
2.1 植株形成珠芽能力的不同
通过对5个种源地的毛建草产生珠芽的植株数量比较(表1),发现采自芦芽山的毛建草植株在3年栽培期内都产生珠芽;采自云顶山和恒山的毛建草产生珠芽的植株数量随着种植年限的延长而增加;采自朝阳半截沟和关帝山的毛建草产生珠芽的植株在2019和2020年都是同样的植株。
在2019年粗略性比较种植的毛建草产生的珠芽数量,发现采自芦芽山的毛建草单株产生的珠芽数量最多达到了43颗,最少的有13颗,观测到植株间产生的珠芽数量有较大差异;采自关帝山的1株毛建草产生的珠芽有19颗,有1株毛建草仅产生2颗珠芽;其他种源地采集的毛建草产生的珠芽基本在2~6颗之间。
对芦芽山、关帝山和恒山3个采集地的毛建草进行调查,发现3个采集地的植株都没有珠芽产生,与种植于林业站的毛建草植株形成了鲜明的差异。
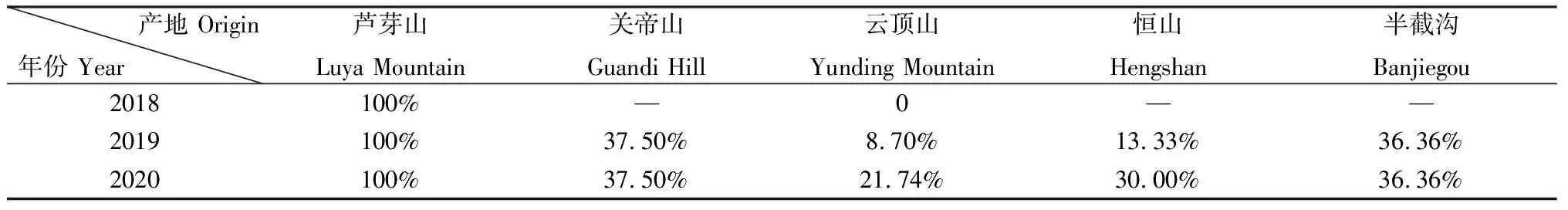
表1 不同产地的毛建草产生珠芽的植株比率Table 1 Ratio of bulbil plants of Dracocephalum rupestre collected from different habitats
2.2 珠芽形成过程
毛建草开完第一批花后,在花茎下部的叶片(有叶柄)和中上部的苞片(无叶柄,且抱茎)内的侧芽开始萌发并发育形成小枝(图1A),萌发的侧芽发育的结果不同,有些侧芽节间伸长发育成花序开花(图1B),有的侧芽节间皱缩成莲座状,叶片着生处膨大形成了珠芽(图1C,1D,1E)。在一个叶腋能产生多个侧芽,这个部位萌发的一部分侧芽形成花序开花,一部分则成为珠芽。所有的花茎都能长出珠芽,只是珠芽的数量及大小有差异。
珠芽在发育过程中,不断的通过花茎从母株上获得营养,不断抽生新叶,珠芽持续膨大,最后发育成木质化并缩短的茎块,类似鳞茎。在10月时,毛建草地上部逐渐枯黄时,花茎及珠芽上的叶片也相继干枯,珠芽顶端不再抽生新叶(图1E),进一步随着母株地上部枯黄而枯萎。

图1 毛建草珠芽发育状态Fig.1 Development status of bulbil of Dracocephalum rupestre注:A为珠芽着生部位;B为珠芽开花;C为未开花的莲座状叶序,D为珠芽膨大;E为珠芽木质化Note:A,bulbil location;B,bulbil blooming;C,rosette phyllotaxy without blossom;D,bulbil fication;E,lignified bubil
2.3 不同部位的的珠芽原地生根率不同
从芦芽山采集的毛建草植株地上部开始枯黄时检查珠芽生根情况,发现只有3颗着生在花茎基部及靠近地表的珠芽着地生根(图2A,2B箭头指向),占检查珠芽数量的1%不到,2颗珠芽在靠近地面处长出1 cm左右的气生根(图2C箭头指向),占检查珠芽数量的0.7%。在毛建草株丛中央的花茎上着生的珠芽没有生根,生根的珠芽主要是着生在株丛外围的花茎上,并且花茎为斜生状态,离地表较近,有利于珠芽底端附着在地表。

图2 毛建草珠芽原位生根状态Fig.2 In situ rooting status of the bulbils of Dracocephalum rupestre注:A为花茎基部珠芽生根;B为花茎中部的珠芽着地生根;C为花茎上部珠芽气生根Note:A,at the base of a flower stalk;B,at the middle of a flower stem by touchdown roots;C,at the top of a flower stalk by aerial roots
2.4 不同大小的珠芽扦插成活率及生根率不同
在10月10日第一次检查扦插的珠芽生根情况时,发现珠芽大小不同时,其成活及生根率也不一样。大珠芽成活率为78.45%,30棵珠芽生根率为100%,根系不仅长,而且数量多,最长7.1 cm(图3A);中珠芽成活率为75.68%,检查的30棵珠芽生根率也为100%,但其根长较短且数量少,根长为2.3~4.3 cm(图3B,3C);小珠芽成活率为58.20%,生根率为0(图3D)。在11月10日,仅检查了小珠芽,发现小珠芽也已经生根,但根系较少。那些死亡的珠芽多为腐烂或者是干枯的。

图3 珠芽扦插生根情况比较Fig.3 Rooting of cuttings in bulbil of Dracocephalum rupestre注:A为大珠芽;B,C为中珠芽;D为小珠芽Note:A,big bulbil;B,C,medium bulbil;D,little bulb
3 讨论
3.1 种源地影响珠芽发生
半夏(Pinelliaternata)不同居群,其产生珠芽的能力不同,有的根本不会产生珠芽[4],并且珠芽的发生频率与叶形有关[5]。石竹科(Caryophyllaceae)的一些植物采用部分克隆、不克隆或偶尔克隆的方式[1],增加其繁殖方式的多样性,有助于种群的繁衍。从芦芽山采集的毛建草植株全部产生珠芽,从云顶山和恒山采集的毛建草随着种植年限的增加,其产生珠芽的植株数量在增加,而从关帝山和朝阳半截沟采集的毛建草产生珠芽的植株数量却没有随着种植年份的延长而增加。说明毛建草珠芽的产生不仅与种源地有关,而且也跟植株个体基因型有关。
在3个采集地没有发现毛建草植株产生珠芽,而引种到太谷后,毛建草或多或少的产生了珠芽,这表明其生态环境及其气候环境的变化对珠芽产生起到了重要作用。高山垫状植物在温和的气候条件下依旧能保持其生长型,是由遗传特性所造成的,植物的生活型和形态在全世界所有山区都有其一致性[1]。当生长环境改变时,植物在不同环境因子作用下,其形态、生理、生态性状与繁殖策略就会发生适应性的改变[6-8]。毛建草作为一种垫状植物,从高海拔或高纬度种源地引种到低海拔地栽培时,仍然保持垫状的生长形态,但却产生了珠芽繁殖方式,增加了一种繁殖策略,但这种气候变化对珠芽的产生机制暂未清晰。
3.2 生长节律影响珠芽发生
珠芽不仅发生在叶腋处,而且也能在花序的中下部发生,花序中的珠芽是由小穗发育而来的,属于无性营养繁殖器官[9-10]。台闽苣苔(Titanotrichumoldhamii)的花序不但能结种子而且在花期结束时的花序轴下部有珠芽产生[11],珠芽蓼(PolygonumviviparumL.)的花序上不但能产生种子,同时在其花发育失败后也能产生珠芽,这表明花茎的顶端优势在抑制腋芽的萌发[12]。毛建草珠芽发生在花后和种子发育期间,表明其生长发育具有一定的时序性,或许也是受顶端优势的抑制,在种子发育期间产生珠芽,这与台闽苣苔珠芽的形成具有相似性。
珠芽蓼借助地下根状茎、分期发育形成的种子或者是花受精发育失败后形成的珠芽使得其形成了多梯级、多形式的多种繁殖系统进行繁殖[13],有性与无性繁殖的有机结合,是克隆植物长期与环境互作中形成的灵活多变的繁育策略,极大地提高了繁殖成功的机会,保证了该物种在复杂多变的环境条件中生存并延续下去[13-14]。珠芽具有繁殖系数高、生活力强等优点,且多产生于地上部分的叶腋间或花序轴而有利于其扩散和传播[15]。栽培的毛建草不仅能产生大量种子,而且种子萌发率也较高,符合珠芽类植物的繁殖策略。
3.3 珠芽生长部位影响其能否发育成新的植株
半夏珠芽在土壤中形成则会生根发芽成为新块茎[16],在珠芽萌发期间保持土壤湿度对提高珠芽萌发率和促进半夏幼苗的生长均具有重要作用[17]。通过毛建草珠芽扦插和原地生根的观察,发现毛建草珠芽在扦插时能生根,接近地面的珠芽在母株周围也能生根,并且能继续长叶发育成植株,离地面较近的珠芽能偶然产生气生根,并未深扎进土壤中,而离地面较远的珠芽则未生根。该现象说明珠芽的着生位置,其能否接触土壤,以及是否具有足够的空气湿度影响珠芽生根并发育成植株。很多偶然的高山克隆植物在某些特定的环境中能进行无性繁殖,如茎枝或花梗在被土埋或基质潮湿松软时,就会产生不定根,进而生长成一株独立的植株个体[1]。半夏的倒苗有助于珠芽落地生根,发育成新的植株,而倒苗是对不利环境条件的一种适应,倒苗次数越多,产生的珠芽总数也越多[5]。能生根的毛建草珠芽主要发生在母株外围的珠芽,而在母株株丛中心抽生的花茎上发生的珠芽则不生根,这进一步说明着生于母株基部斜生的花茎上,并且靠近土壤的珠芽接触土壤后就能生根,表明珠芽生长部位影响着其能否落地生根,这与半夏珠芽生根的研究结果一致。
半夏的传播方式无论是珠芽还是果实都缺乏有效的传播媒介,大多数情况下,珠芽和果实都掉落在母株附近,而母株的生境大多能适合其子代的生长,这种传播方式使子代过分集中,从而往往导致一些个体在生存竞争中被淘汰[4]。毛建草产生较多的珠芽,但由于在生长季不能全部落地生根,珠芽随着植株干枯,表明并不会有太多的珠芽发育成新的植株,从而产生与母株竞争的后果。而少量接触土壤生根的珠芽则在母株的外围,既可以增加种群扩大的潜力,也不会对母株产生大的竞争,有的珠芽可能会凋落地上,掉落的珠芽是否会被土埋,次年是否会生根成长为新的植株,目前尚未观察到。
3.4 珠芽大小影响珠芽生根的快慢
半夏珠芽萌发率与珠芽大小有显著的正相关,珠芽越大活力越强,萌发率越高,萌发的新个体较健壮,幼苗生长较好[18]。扦插毛建草珠芽时,珠芽大且叶多的生根较好,而珠芽小且叶少的生根较慢,在规定期限内生根率较低,这与半夏珠芽大小影响其萌发的规律有相似性。毛建草的珠芽在扦插过程中,能连续萌发新叶,与珠芽蓼的珠芽萌发一样,没有休眠特性,具有较高的发芽率[17 ]。
薯蓣(DioscoreacirrhosaL.)珠芽外形多样,其本质为茎,是一种特殊的变态茎[19],成熟珠芽最外为木栓质表皮,其内为木栓形成层形成的周皮[20]。薯蓣珠芽外观呈现嫩绿色球形小珠芽时,内部已分化出顶芽和多处根原基[2],顶芽也是埋藏在周皮以内,在外方一般见不到顶芽的存在[20]。毛建草珠芽扦插时能生根长叶,说明其珠芽也具有了根原基和顶芽。毛建草珠芽基部木质化,有助于防止其失水,因此,未生根的毛建草小珠芽在基质中仍能使叶片保持存活状态,在扦插期内能继续生长新叶,并未死亡,表明珠芽木质部能吸收水分且供应营养物质,而那些死亡的珠芽则是因为在喷水及下雨时,将蛭石冲刷,使得珠芽裸露干枯,或者是埋深造成的珠芽腐烂。
4 结论
毛建草从高海拔(或高纬度)地引种到低海拔(低纬度)平原区后,由于环境及气候条件发生了变化,使得毛建草在花后及种子发育期间形成了珠芽。毛建草种源及植株个体的差异影响珠芽产生的能力及数量。珠芽具备了在形态学和生理学上生根的能力,靠近土壤表面的珠芽能生根并发育成新的植株,有助于扩展种群生长空间,但是能生根的珠芽较少。无性与有性繁殖方式的结合使得种群既能维持基因型稳定的情况下,又有进化的可能。本研究将为毛建草的繁殖生物学及种群生态学提供新的切入点,增加了珠芽类植物的种类数量。
猜你喜欢
安徽农业科学(2023年20期)2023-11-01 02:24:34
贵州农业科学(2023年10期)2023-10-27 00:05:42
浙江农业学报(2021年4期)2021-04-27 07:40:02
特种经济动植物(2021年2期)2021-02-25 05:47:34
西藏农业科技(2019年2期)2019-07-10 09:23:12
花卉(2017年23期)2017-12-05 07:44:01
现代园艺·综合版(2016年17期)2016-10-17 04:23:16
果树实用技术与信息(2016年8期)2016-09-21 03:25:35
长江蔬菜(2014年2期)2014-01-11 04:35:24
食品科学(2013年17期)2013-03-11 18:26:53
