
滩湖杂羊多胎基因(FecB)TaqMan 探针SNP 分型的研究
2023-05-25 02:07马丽娜侯鹏霞梁小军
中国畜禽种业 2023年5期
李 丹,马丽娜,施 安,王 燕,侯鹏霞,梁小军*
(1.西北农林科技大学动物科技学院,陕西西安 712100;2.宁夏农林科学院动物科学研究所,宁夏银川 750002)
Booroola (FecB)基因是在绵羊中鉴定出的第一个高繁殖力主效基因[1],其遗传效应是增加排卵数和产羔数[2]。目前,FecB 突变作为高产羔数的分子标记已广泛应用于绵羊育种改良中。湖羊繁殖力高、性成熟早且含有FecB 基因;而宁夏滩羊虽然肉质鲜美,但产羔率低[3-4]。为提高滩羊繁殖率,同时发挥湖羊多胎高产的优势,通过滩羊公羊和湖羊母羊进行杂交生产滩湖杂羊,通过横交固定得到滩湖杂羊群体G0、G1、G2代。本研究应用TaqMan 探针技术,对滩湖杂交羊G0,G1,G2代血液样本中的FecB 基因进行分析,将传统的常规表型选择育种方法与FecB 基因分子标记辅助选择育种技术相结合,旨在加快滩羊多胎品种(系)的选育步伐。
1 材料与方法
1.1 试验动物
选配方法:试验羊来自同心荣华养殖场,滩羊公羊与湖羊母羊杂交产滩湖F1代,滩湖F1代母羊与滩羊公羊杂交产滩湖F2代,滩湖F2代母羊与滩湖F2代公羊横交固定产滩湖横交固定一世代(G0代),进一步繁殖产滩湖横交固定二世代(G1代)、滩湖横交固定三世代(G2代)。
2022 年6 月,选取核心群横交固定三个世代的滩湖杂交羊共计242 只作为试验动物,其中G0代78 只,G1代142 只,G2代22 只。
1.2 血样采集
每只羊颈静脉采血5mL,保存于EDTA 抗凝管中,采血后颠倒混匀以防止血液凝固,于-20℃冷冻保存。
1.3 DNA 提取与纯度检测
使用天根全血基因组DNA 提取试剂盒(DP318)提取,严格按照试剂盒说明书操作,使用0.8%的琼脂糖凝胶电泳和紫外分光光度计对得到的DNA 片段的浓度和纯度进行检测。
1.4 引物及探针序列
利用Taqman 探针检测FecB 基因多态性的方法 (CN105176982A)进行FecB 检测分型 (表1)。BB 型代表FecB 基因CDS 区第746 位碱基发生A 至G 的突变(g.A746G),导致FecB 蛋白第249 的谷氨酰胺突变为精氨酸(p.Q249R),++型代表FecB 基因的CDS 区第746 位碱基未发生改变,仍为A。

表1 引物与探名序列
1.5 RT-PCR 反应体系和反应程序
RT-PCR 反应体系为6μL:基因组DNA 1μL,2×Master Mix 3μL,10μmol/L 正向、反向引物各0.3μL;10μmol/L 探针P-G 和P-A 各0.15μL,去离子水补齐至6μL。
RT-PCR 反应程 序:95℃ 10min,95℃30s,60℃60s,40 个循环。
2 结果分析
2.1 DNA 提取及检测
本试验中提取的血液基因组DNA 的OD 值为1.7~1.9,并经0.8%的琼脂糖凝胶110V 电泳20min 可见各泳道都有一条清晰的带(图1),说明基因组DNA 质量良好、无降解也无RNA 污染,可直接用于后续探针分型。

图1 DNA 琼脂糖凝胶电泳
2.2 滩湖杂羊FecB 基因TaqMan 探针分型结果
由图2 可见,FecB 基因TaqMan 探针分型显示FecB 基因在滩羊群体中具有3 种基因型,分别为BB、B+、++。
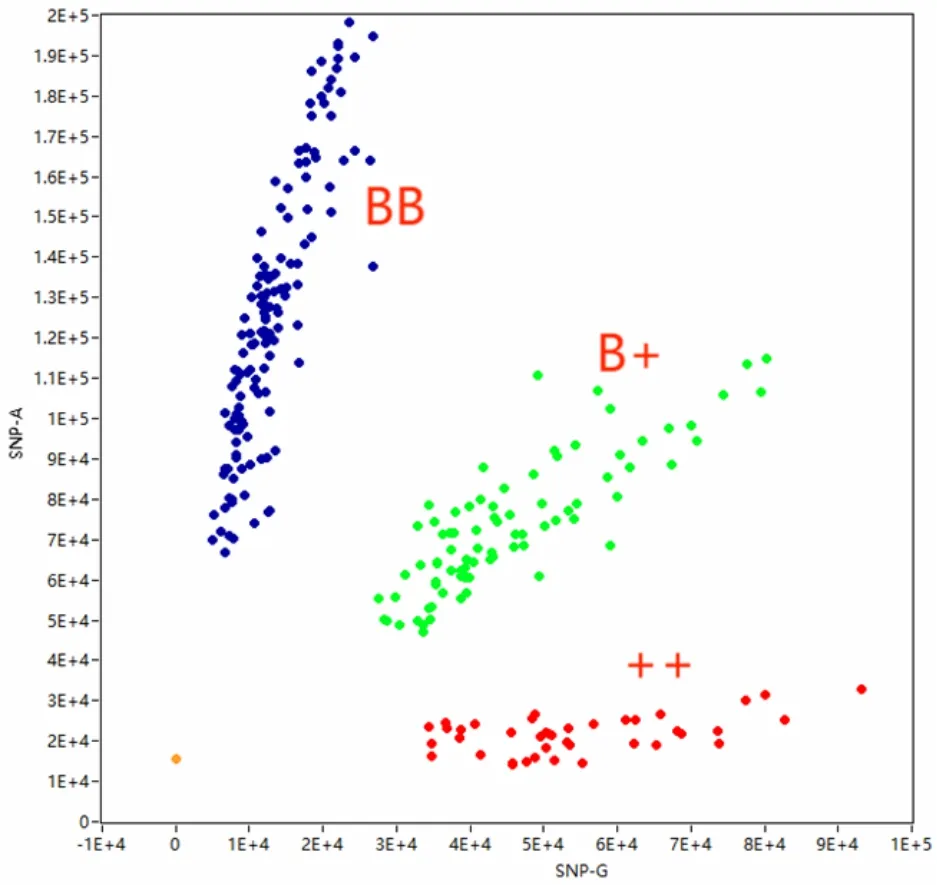
图2 FecB 基因基因分型散点图
2.3 滩湖杂羊FecB 基因基因型结果
由表2 可知,G0代、G1代、G2代的FecB 基因的等位基因频率与基因型频率,G0代群体中BB、B+、++基因型频率分别为2.56%、14.10%和83.33%,B、+等位基因频率分别为9.62%、90.38%。G1代群体中BB、B+、++基因型频率分别为28,17%、38.73%和33.10%,B、+等位基因频率为47.54%和52.46%。G2代群体中BB、B+、++基因型频率分别为18.18%、54.54%和27.27%,B、+等位基因频率分别为45.45%、54.54%。

表2 不同横交固定世代FecB 基因分布情况
3 讨论
刘秋月等[5]建立的检测FecB 突变的TaqMan探针法,是一种通过检测PCR 过程中及结束后产生的荧光信号来区分等位基因类型的SNP 检测技术[6]。该方法可进行大批量的高效检测,也是近年来广泛应用于绵羊FecB 突变的检测技术。Zhou 等[7]利用TaqMan 探针法检验FecB 突变,并统计分析了不同基因型群体的产羔率,发现FecB突变显著影响绵羊的繁殖性能。马丽娜等[8]认为FecB 基因可以作为滩羊多胎性选育的分子遗传标记,并能有效提高滩羊个体平均产羔数,通过基因型选育,还可以加快滩羊高繁品系的建立。李君等[9]研究发现,FecB 基因在黄淮杜泊羊群体中存在BB、B+、++等3 种基因型,其中B+型(杂合型)是群体中的优势基因型,BB 和B+基因型产羔数极显著高于++基因型。郭立宏[10]利用SSCP 标记对东北细毛羊FecB 基因进行检测,认为FecB 基因是控制多羔性状的主效基因,选择BB 和B+基因型母羊留作种用,可以加快东北细毛肉羊多胎品系的育成速度。
本试验利用TaqMan 探针对242 只滩湖杂交后代FecB 基因进行SNP 分型检测,结果表明,滩湖杂交后代GO、G1、G2代中存在BB、B+、++3 种基因型。随着横交固定体系的持续开展,B+基因型频率依次升高,++基因型频率依次降低,而BB 基因型理论上与B+型的规律应一致保持协同增长趋势,但G2代的BB 型低于G1代次,可能是此次试验抽取的G2群体数量太少,样本没有代表性容易被偏差个体影响整体试验结果,影响了BB 基因型在G2代群体中的分布频率。因此还需进一步扩充G2代样本量为品系选育选配提供稳定的FecB 突变背景和遗传材料。随着横交固定阶段的延伸,等位基因频率B、+等位基因变化规律与基因型频率类似。
罗生金[11]将湖羊的FecB 基因导入哈萨克羊群体中,用湖哈F1代公母羊(基因型均为B+)横交,产羔率达到172%,湖哈F1代公羊(基因型为B+)与湖哈F1代母羊(基因型为BB)横交,产羔率达到200%,而湖哈F1代公羊(基因型为B+)与湖哈F1代母羊(基因型为++)横交,产羔率为109%。李彬等[12]在滩羊群体中导入小尾寒羊的FecB 基因,古丽格娜等[13]在吐鲁番黑羊群体中导入策勒黑羊的FecB 基因,均增加了后代的产羔率。
这些成功的实例证明,在繁殖力较低的群体中导入FecB 基因,能有效地提高绵羊群体的繁殖力。
4 结论
本研究在滩羊群体中导入湖羊的FecB 基因,并采用基因检测技术选留含有B 基因的个体,提高群体的B 基因频率。因此,利用基因型选育技术,结合传统选种方法,通过选留含有B 基因的公母羊繁殖,可显著改善滩羊FecB 基因的基因型结构和基因频率,提高群体的产羔数,加快滩羊高繁品种(系)的建立。
猜你喜欢
美食(2021年11期)2021-12-04
中国饲料(2021年17期)2021-11-02
现代畜牧科技(2021年8期)2021-10-13
现代畜牧科技(2021年7期)2021-07-28
现代畜牧科技(2021年6期)2021-07-16
现代畜牧科技(2021年2期)2021-03-19
现代畜牧科技(2018年6期)2018-10-21
山东农业科学(2018年12期)2018-03-04
畜牧兽医科技信息(2018年12期)2018-02-14
新农业(2017年24期)2018-01-30
