
过表达CBX7调控PTEN/Akt信号通路干预上皮-间充质转化抑制胃癌细胞迁移、侵袭
2023-05-24 06:40刘剑波李芳芳钟月圆王秀丽郴州市第一人民医院中心医院消化内科湖南郴州423000
局解手术学杂志 2023年5期
谭 斌,刘剑波,李芳芳,彭 艳,钟月圆,邓 晖,王秀丽 (郴州市第一人民医院中心医院消化内科,湖南 郴州 423000)
胃癌是消化道恶性肿瘤之一,其发病率在我国恶性肿瘤中居第2位,病死率居第3位[1]。近年来,医学技术的进步、手术方法的改进、免疫治疗及新辅助化疗的应用极大地延长了胃癌患者的生存期,然而肿瘤转移和复发仍不可避免,导致胃癌患者预后差、生存质量低[2-3]。因此,深入研究胃癌发展、转移的分子机制极为必要。上皮-间充质转化(epithelial-mesenchymal transition,EMT)是肿瘤远处转移过程中的关键步骤之一,临床上,EMT与癌症预后不良密切相关[4-5]。染色体盒蛋白同源物7(chromosome box protein homologue 7,CBX7)是多梳蛋白家族中的一员,属于黑腹果蝇的异染色质蛋白在人类细胞中的同源物,主要分布于细胞核的染色质中,通过与染色质结合发挥作用[6]。CBX7的表达与多种癌症的不良预后和侵袭转移相关,在癌症发生和进展中起着重要的作用[7-8]。研究发现,CBX7在胃癌组织中低表达,其低表达与胃癌患者的不良预后和生存率低显著相关[9],CBX7通过p16和Akt/NF-κB/miR-21信号通路调节胃癌细胞的干细胞样特性[10]。但CBX7对胃癌细胞迁移、侵袭及EMT的影响尚不明确。因此,本研究就CBX7对胃癌细胞迁移、侵袭及EMT的影响展开探讨,并分析其潜在作用机制。
1 材料与方法
1.1 实验材料
人胃癌细胞SGC7901(中国科学院细胞库);CBX7过表达质粒和阴性对照质粒、CBX7 siRNA和阴性对照siRNA(上海吉玛生物公司);RPMI-1640培养基(深圳华晨阳科技有限公司);Lipofectamine 2000转染试剂盒(美国Invitrogen公司);CCK-8试剂盒、BCA蛋白定量试剂盒、ECL试剂盒(上海碧云天生物技术公司);结晶紫染色液(武汉卡诺斯科技有限公司);Matrigel基质胶、Transwell小室(美国BD公司);CBX7和GAPDH引物(上海捷瑞生物工程有限公司);RNA提取试剂盒、反转录试剂盒、SYBR Premix Ex Taq试剂盒(大连宝生物工程有限公司);兔源GAPDH、E-钙黏蛋白(E-cadherin)、N-钙黏蛋白(N-cadherin)、波形蛋白(Vimentin)、磷酸酶张力蛋白同源物基因(phosphatase and tensin homolog deleted on chromosometen,PTEN)、蛋白激酶B(Akt)、磷酸化Akt(p-Akt)单克隆抗体及生物素偶联的IgG二抗(英国Abcam公司)。
Heracell VIOS 160i细胞培养箱(美国Thermo Fisher公司);PT-3502PC多功能酶标仪(北京普天新桥技术有限公司);WMF-3690型倒置荧光显微镜(上海无陌光学仪器有限公司);JY-ZY5型电泳仪(济南君意生物科技有限公司);RTQ-960 Pro qRT-PCR仪[艾康生物技术(杭州)有限公司];KETA GL型凝胶成像系统(北京好亿科技发展有限公司)。
1.2 细胞培养
人胃癌细胞SGC7901置于RPMI-1640培养基中培养,培养基中添加10%胎牛血清和1%青霉素-链霉素,培养箱温度为37 ℃,CO2浓度为5%。
1.3 细胞转染与分组
按照Lipofectamine 2000转染试剂盒说明书,将CBX7过表达质粒和阴性对照质粒、CBX7 siRNA和阴性对照siRNA转染至对数生长期的SGC7901细胞,依次记为pcDNA-CBX7组、pcDNA-NC组、si-CBX7组和si-NC组,同时将正常培养的细胞作为阴性对照组(NC组)。转染48 h后检测转染效率。
1.4 qRT-PCR法检测细胞中CBX7 mRNA表达水平
收集各组SGC7901细胞,采用TRIzol试剂提取RNA并进行定量,用反转录试剂盒反转录合成cDNA,按照SYBR Premix Ex Taq试剂盒说明书配置实验体系,进行qRT-PCR检测。引物序列如下:CBX7上游引物为5'-CATGGAGCTGTCAGCCATC-3',下游引物为5'-CTGTACTTTGGGGGCCATC-3';GAPDH 上游引物为5'-ACCCAGAAGACTGTGGATGG-3',下游引物为5'-CAGTGAGCTTCCCGTTCAG-3'。 反 应条件:95 ℃3 min,95 ℃ 15 s,60 ℃ 20 s,72 ℃ 10 s,共40 次循环。实验重复 3次,采用 2-ΔΔCt法进行分析。
1.5 CCK-8法检测细胞增殖活性
各组SGC7901细胞按1×104/mL接种至96孔培养板,设置5个复孔,同时设置对照孔和空白孔,培养24 h后,每孔加入10 μL CCK-8试剂孵育4 h,采用酶标仪测定各孔在450 nm波长处的吸光度(OD)值。实验重复3次,细胞存活率(%)=(实验孔OD值-空白孔OD值)/(对照孔OD值-空白孔OD值)×100%。
1.6 细胞划痕实验检测细胞迁移能力
各组SGC7901细胞按1×105/mL接种至6孔培养板,培养至细胞贴壁生长后,以200 μL移液枪枪头垂直划痕,做好标记,PBS冲洗,加入RPMI-1640培养基继续培养24 h。实验重复3次,细胞迁移率(%)=(0 h划痕距离-24 h划痕距离)/0 h划痕距离×100%。
1.7 Transwell小室检测细胞侵袭能力
收集各组SGC7901细胞,取100 μL浓度为2×105/mL的细胞悬液接种至含有Matrigel基质胶包被膜的Transwell小室上室,下室添加600 μL含10%胎牛血清的RPMI-1640培养基,孵育24 h后,清除上室细胞,用4%多聚甲醛固定穿过膜的细胞,并用0.1%结晶紫染色20 min,倒置显微镜下观察,计数侵袭细胞数目,实验重复3次。
1.8 Western blot法检测细胞CBX7蛋白、EMT相关蛋白和PTEN/Akt信号通路蛋白表达水平
蛋白裂解液提取各组SGC7901细胞总蛋白,BCA试剂盒定量,SDS-PAGE分离蛋白样品,并湿转至PVDF膜上,用5%脱脂牛奶封闭膜2 h,TBST洗膜,将膜与 CBX7、E-cadherin、N-cadherin、Vimentin、PTEN、Akt、p-Akt、GAPDH 一抗(1∶1 000)在 4 ℃孵育过夜,TBST洗膜,加入IgG二抗(1∶5 000)室温孵育1 h,ECL试剂显影,分析蛋白条带灰度值。实验重复3次。
1.9 统计学方法
采用SPSS 25.0软件进行统计学分析,实验数据以均数±标准差(-x±s)表示,多组间比较采用单因素方差分析,进一步两两比较采用LSD-t检验,P<0.05表示差异有统计学意义。
2 结果
2.1 SGC7901细胞中CBX7表达水平
与NC组比较,pcDNA-NC组和si-NC组SGC7901细胞中CBX7 mRNA和蛋白表达量变化均无统计学差异(P>0.05);与pcDNA-NC组比较,pcDNA-CBX7组SGC7901细胞中CBX7 mRNA和蛋白表达量均显著增加(P<0.05);与 si-NC组比较,si-CBX7组 CBX7 mRNA和蛋白表达量均显著下降(P<0.05),见图1,提示转染效果良好。

图1 SGC7901细胞中CBX7表达水平
2.2 过表达CBX7抑制SGC7901细胞增殖、迁移和侵袭
与NC组比较,pcDNA-NC组SGC7901细胞存活率、迁移率及侵袭数变化均无统计学差异(P>0.05);与pcDNA-NC组比较,pcDNA-CBX7组SGC7901细胞存活率、迁移率及侵袭数均显著下降/减少(P<0.05),见图2,提示过表达CBX7抑制SGC7901细胞增殖、迁移和侵袭。

图2 过表达CBX7抑制SGC7901细胞增殖、迁移和侵袭
2.3 过表达CBX7抑制SGC7901细胞EMT
与NC组比较,pcDNA-NC组SGC7901细胞中E-cadherin、N-cadherin、Vimentin蛋白表达量变化均无统计学差异(P>0.05);与pcDNA-NC组比较,pcDNACBX7组SGC7901细胞中E-cadherin蛋白表达量显著升高(P<0.05),N-cadherin、Vimentin蛋白表达量显著下降(P<0.05),见图 3,提示过表达 CBX7抑制SGC7901细胞EMT进程。
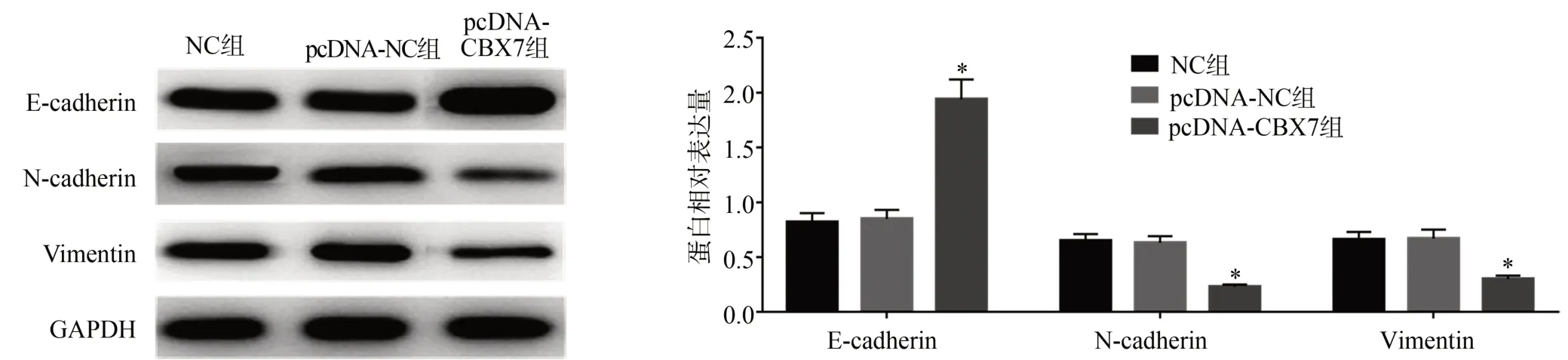
图3 过表达CBX7抑制SGC7901细胞EMT
2.4 下调CBX7表达促进SGC7901细胞增殖、迁移和侵袭
与NC组比较,si-NC组SGC7901细胞存活率、迁移率及侵袭数变化均无统计学差异(P>0.05);与si-NC组比较,si-CBX7组SGC7901细胞存活率、迁移率及侵袭数均显著升高/增加(P<0.05),见图4,提示下调CBX7促进SGC7901细胞增殖、迁移和侵袭。

图4 下调CBX7表达促进SGC7901细胞增殖、迁移和侵袭
2.5 下调CBX7表达促进SGC7901细胞EMT
与NC组比较,si-NC组SGC7901细胞中E-cadherin、N-cadherin、Vimentin蛋白表达量变化均无统计学差异(P>0.05);与si-NC组比较,si-CBX7组SGC7901细胞中E-cadherin蛋白表达量显著下降(P<0.05),N-cadherin、Vimentin蛋白表达量显著升高(P<0.05),见图5,提示下调CBX7促进SGC7901细胞EMT进程。
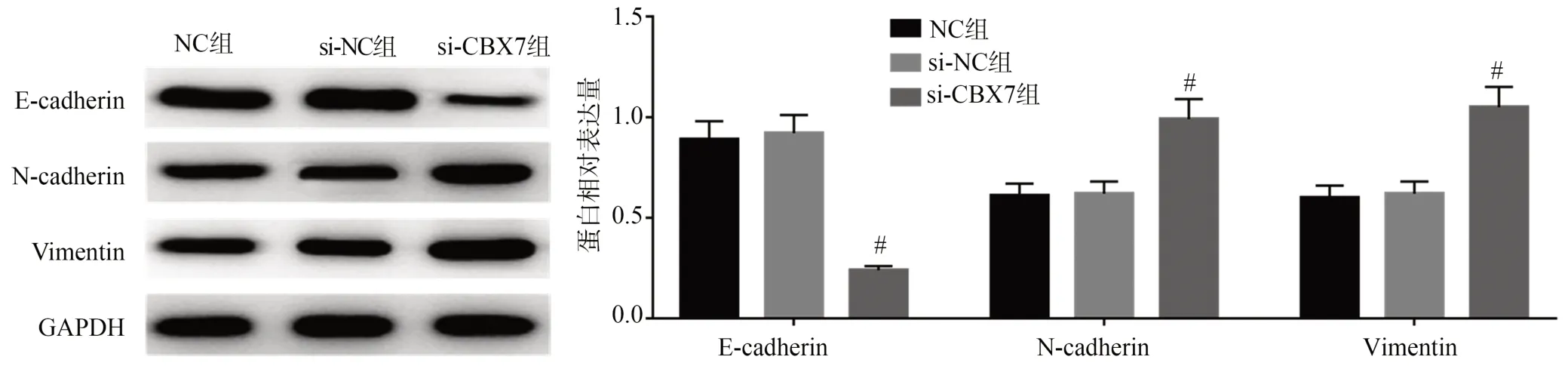
图5 下调CBX7表达促进SGC7901细胞EMT
2.6 CBX7对SGC7901细胞中PTEN/Akt信号通路蛋白表达的影响
与NC组比较,pcDNA-NC组和si-NC组SGC7901细胞中PTEN蛋白表达量、p-Akt/Akt变化均无统计学差异(P>0.05);与pcDNA-NC组比较,pcDNA-CBX7组SGC7901细胞中PTEN蛋白表达量显著升高(P<0.05),p-Akt/Akt显著下降(P<0.05);与si-NC组比较,si-CBX7组SGC7901细胞中PTEN蛋白表达量显著下降(P<0.05),p-Akt/Akt显著升高(P<0.05),见图6,提示过表达CBX7通过上调PTEN抑制Akt信号通路活性。

图6 CBX7对SGC7901细胞中PTEN/Akt信号通路蛋白表达的影响
3 讨论
胃癌起源于胃壁最表层的黏膜上皮细胞,可发生于胃的各个部位,是具有高度侵袭和转移特性的恶性肿瘤。胃癌早期患者临床症状不明显,多数胃癌患者确诊时已发生远处转移,而胃癌细胞发生转移是导致胃癌患者病死率高和预后差的主要原因。因此,探究胃癌侵袭、转移的相关机制,对降低胃癌患者病死率和改善预后具有重要意义。CBX7在哺乳动物大脑、心脏及骨骼肌组织等中均有表达,可维持机体组织的正常生长发育。在细胞中,CBX7通常作为表观遗传调控因子调节基因表达,然而,在病理状态下CBX7的异常表达可导致基因表达失衡,这与肿瘤的发生、发展密切相关。在人类肿瘤中,CBX7起着双重作用,一方面其通过抑制抑癌基因促进某些癌症的进展;另一方面,其通过与不同分子的相互作用调节相关蛋白合成,进而抑制癌症进展[11-12]。在宫颈癌中,CBX7过表达可抑制细胞增殖、迁移和侵袭,诱导细胞凋亡,作为抑癌因子发挥作用[13]。在神经胶质瘤中,CBX7通过沉默CCNE1诱导细胞G1/S期阻滞,可作为神经胶质瘤预后标志物[14]。此外,CBX7可作为miR-19及miR-18a等多种microRNAs的靶基因,对卵巢癌、非小细胞肺癌等肿瘤细胞迁移、侵袭及EMT发挥调控作用[15-16]。但CBX7对胃癌细胞迁移、侵袭的影响尚不清楚。本研究通过调控CBX7的表达,分析CBX7对胃癌细胞增殖、迁移和侵袭的影响,结果显示,过表达CBX7可抑制胃癌SGC7901细胞增殖、迁移和侵袭,下调CBX7表达可促进胃癌SGC7901细胞增殖、迁移和侵袭,提示CBX7可通过调控细胞增殖、迁移和侵袭而调控胃癌进展。
EMT是一个复杂的生物学过程,不仅包括细胞-细胞连接的溶解,还包括顶端外侧极性的丧失。在EMT过程中,上皮细胞失去细胞黏附蛋白(如E-cadherin)和紧密连接蛋白的表达,并同时表达丰富的N-cadherin和Vimentin等间充质标志物,从而完成从上皮到间充质的表型转化,转化后的细胞表现出细胞间黏附减少和运动性增加,使肿瘤细胞获得较强的迁移性和侵袭性[17]。研究报道,复合物CBX7-PRMT1在调节E-cadherin表达和细胞迁移中起关键作用[18]。Tian等[19]报道,CBX7表达降低与宫颈癌EMT和不良预后相关。本研究探讨CBX7表达变化与胃癌EMT的相关性,结果显示,过表达CBX7后SGC7901细胞中E-cadherin蛋白表达升高,N-cadherin、Vimentin蛋白表达下降,下调CBX7表达后SGC7901细胞中E-cadherin蛋白表达下降,N-cadherin、Vimentin蛋白表达升高,提示CBX7可调控胃癌细胞SGC7901的EMT进程。
肿瘤的发生、转移过程涉及多个信号途径和多种分子机制,其中PTEN/Akt通路是近年来研究较多的相关通路之一。PTEN是一种磷酸酶,通过负调节Akt信号通路调节各种转录因子和信号分子的活性,进而调控细胞周期、细胞凋亡,并参与细胞增殖、迁移和侵袭。在胃癌发生、发展进程中,PTEN/Akt信号通路参与胃癌血管生成以及胃癌细胞增殖、迁移、凋亡和EMT等生物学过程。Zhou等[20]报道,靶向miR-21通过PTEN/Akt信号通路抑制胃癌细胞增殖和迁移。Wu等[21]报道,miR-616-3p通过PTEN/Akt/mTOR通路促进胃癌血管生成和EMT。Qiang等[22]报道,姜黄素通过调控miR-21/PTEN/Akt通路,与PD98059协同诱导人胃癌细胞凋亡。还有研究显示,CBX7可通过调节PTEN/Akt信号通路抑制癌细胞增殖、迁移和侵袭[23]。本研究结果显示,过表达CBX7显著上调SGC7901细胞中PTEN蛋白表达,使Akt磷酸化减少;而下调CBX7表达则使SGC7901细胞中PTEN蛋白表达显著下调,Akt磷酸化增加。提示CBX7可通过调控PTEN/Akt信号通路参与胃癌细胞EMT进程及细胞迁移和侵袭。
综上所述,本研究证实过表达CBX7可抑制胃癌细胞增殖,并可通过抑制EMT抑制胃癌细胞迁移、侵袭,其机制可能是与调控PENT/Akt信号通路有关。因此,CBX7可能是胃癌转移和预后的相关标志物及潜在治疗靶点。
猜你喜欢
学苑创造·A版(2020年12期)2020-01-07
中国外汇(2019年15期)2019-10-14
作文教学研究(2016年1期)2016-07-05
中国卫生标准管理(2015年3期)2016-01-14
法医学杂志(2015年4期)2016-01-06
法医学杂志(2015年4期)2016-01-06
医学研究杂志(2015年9期)2015-07-01
医学研究杂志(2015年8期)2015-06-22
中国当代医药(2015年20期)2015-03-01
中国中医药现代远程教育(2014年22期)2014-03-01
