
四种栎属青冈亚属植物叶绿体基因组特征及系统发育研究
2023-05-22 01:55黄婷唐梦陈晓丽李卜宇张雪梅
广西植物 2023年4期
黄婷 唐梦 陈晓丽 李卜宇 张雪梅


摘 要:栎属青冈亚属植物的系统发育地位长期存在着争议,部分种的种间关系不明确。为揭示宁冈青冈(Quercus ningangensis)、曼青冈(Q. oxyodon)、毛曼青冈(Q. gambleana)、竹叶青冈(Q. neglecta)的叶绿体基因组特征及系统发育关系,该研究选择以上4种栎属青冈亚属植物的成熟叶片进行二代测序,对其叶绿体基因组结构和特征进行分析,并结合相关类群进行系统发育研究。结果表明:(1)宁冈青冈、曼青冈、毛曼青冈、竹叶青冈的叶绿体基因组序列长度分别为160 906、160 883、160 832、160 784 bp,均编码133个基因,包括88个蛋白质编码基因、37个tRNA基因、8个rRNA基因。(2)4种栎属青冈亚属植物偏好以A/T结尾的密码子,质体基因组变异区域主要存在于非编码序列。(3)通过IR边界分析得出,4种栎属青冈亚属植物存在ycf1假基因且在IRb/SSC区域发生扩张。(4)系统发育分析显示,在壳斗科中,水青冈属(Fagus)和轮叶三棱栎属(Trigonobalanus)较早分化出来,栎亚属(subg. Quercus)未形成一个单系群,叶绿体基因组建树结果和核标记、质体标记一致,栎亚属中Ilex组和Cerris组间出现穿插现象。(5)毛曼青冈的系统发育地位存在着争议,《中国植物志》及部分地方植物志将毛曼青冈作为一个单独的种处理;周浙昆、邓敏等学者根据叶的特征将毛曼青冈作为曼青冈的一个亚种处理;而该研究根据叶绿体基因組信息并结合前人的形态分析结果,支持毛曼青冈作为一个独立的种存在的观点。该研究结果为探讨栎属青冈亚属系统发育地位、栎属青冈亚属组的划分、存疑种种间关系的解决提供了基础资料。
关键词: 壳斗科, 栎属青冈亚属, 基因组比较, 叶绿体基因组, 系统发育研究
中图分类号:Q949
文献标识码:A
文章编号:1000-3142(2023)04-0741-14
Abstract:The phylogenetic status of Quercus subg. Cyclobalanopsis has long been controversial and the interspecific relationships of some species are unclear. At present, chloroplast genomes have been used to solve the phylogenetic problems of subg. Quercus and Castanea in the Fagaceae, but there are few researches on solving the phylogenetic problems of Quercus subg. Cyclobalanopsis with chloroplast genome information. To reveal the chloroplasts genome characteristics and phylogenetic relationships of Quercus ningangensis, Q. oxyodon, Q. gambleana and Q. neglecta. In this study, mature leaves of the above four species of Quercus subg. Cyclobalanopsis were selected for next-generation sequencing, and the chloroplast genome structure and characteristics were analyzed, phylogeny was studied in combination with related taxa. The results were as follows: (1) The chloroplast genome sequences of Q. ningangensis, Q. oxyodon, Q. gambleana and Q. neglecta were 160 906, 160 883, 160 832, 160 784 bp, respectively. They all encoded 133 genes, including 88 protein-coding genes, 37 tRNA genes and 8 rRNA genes. (2) The codons ending in A/T were preferred by the four species of Quercus subg. Cyclobalanopsis, and the variation regions of plastid genome mainly existed in non-coding sequences. (3) According to IR boundary analysis, ycf1 pseudogene was found in four species of Quercus subg. Cyclobalanopsis and expanded in IRb/SSC regions. (4) Phylogenetic analysis showed that Fagus and Trigonobalanus were differentiated early in Fagaceae, while subg. Quercus did not form a monophyly. The results of phylogenetic tree based on chloroplast genome were consistent with the nuclear marker plasmid markers. Interspersed between Ilex group and Cerris group in subg. Quercus. (5) The Flora of China and some local flora treated Q. gambleana as a separate species, while ZHOU Zhekun and DENG Min et al. treated Q. gambleana as a subspecies according to the characteristics of leaves. The phylogenetic status of Q. gambleana is still controversial. Based on the chloroplast genome information and previous morphological analysis results, the present study supported the idea that Q. gambleana existed as an independent species. This study provides basic data for discussing the phylogenetic status of the Quercus subg. Cyclobalanopsis, the division of its groups, and the resolution of doubtful relationships among them.
Key words:Fagaceae, Quercus subg. Cyclobalanopsis, genome comparison, chloroplast genome, phylogenetic study
叶绿体广泛存于高等植物及大部分藻类中,是绿色植物进行光合作用的场所。叶绿体自身拥有独立的遗传物质,其基因组为四分体结构,由一个大单拷贝区域(long single copy, LSC)、一个小单拷贝区域(small simple copy, SSC)及两个反向重复区域(inverted repeat, IRA and IRB)构成,基因组序列和结构都相对保守(Zhao et al., 2018)。与核基因组比较结果显示,叶绿体基因组具有结构简单、分子量小、基因的替换率较低、单拷贝等特点(Korpelainen, 2004; Kwak et al., 2019)。近年来,随着二代测序时间和成本的降低,越来越多的叶绿体基因组的测序分析结果已经用于模糊种的鉴定和系统发育研究。目前,虽然已有研究利用叶绿体基因组解决壳斗科栎亚属(subg. Quercus)、栗属(Castanea)的系统发育问题(杨颜慈, 2018;高潇潇, 2020),但利用叶绿体基因组信息解决栎属青冈亚属系统发育问题的还未多见。
壳斗科(Fagaceae)栎属青冈亚属(Quercus subg. Cyclobalanopsis)共有122种(Frodin & Govaerts, 1998)或150种(Huang et al., 1999),主要分布于亚洲热带和亚热带地区。我国是栎属青冈亚属的重要分布区和多样性中心之一(罗艳和周浙昆, 2001a),共有77种以及3变种,秦岭、淮河以南为栎属青冈亚属植物的主要分布区,为常绿阔叶林的建群种之一(陈焕镛和黄成就, 1998;Huang et al., 1999)。青冈类植物木材具有质地坚硬、耐腐蚀、耐磨损等优点,是制作船桨和其他工具的良好木材;其果实内淀粉含量丰富,含量约为50%~60%(端木炘, 1995),可用于酿酒、制作饲料、糊料和工业淀粉等;树皮中含有单宁,可制作栲胶。青冈类植物的系统发育地位长期存在着争议,部分学者根据壳斗的排列方式将青冈类植物作为一个单独的属处理(徐永椿和任宪威, 1976;郑万钧, 1985;陈焕镛和黄成就, 1998;Huang et al., 1999);而多数学者根据花粉粒解剖学等形态特征及ITS序列,支持青冈类植物作为栎属下的一个亚属处理(中国科学院植物研究所, 1972;刘兰芳和房志坚, 1986;端木炘, 1992;罗艳和周浙昆, 2001b)。由于青冈类植物生殖结构变化较大,叶的特征较为均一,不同类群间存在基因渐渗,馆藏的标本量有限,果实等重要分类特征难以观察(邓敏, 2007),因此其属下没有明确的分组,部分种的种间关系不明确,种的划分上还存在着许多争议。目前,关于栎属青冈亚属系统发育的研究主要还停留在形态学方面。因此,需要更多的分子系统学方面的证据来协助解决栎属青冈亚属的系统发育地位和种间关系。
本研究对4种栎属青冈亚属植物的成熟叶片进行二代测序,依托4种栎属青冈亚属植物的叶绿体基因组信息,拟探究以下问题:(1)4种栎属青冈亚属植物的叶绿体基因组基本特征及其差异;(2)4种栎属青冈亚属植物的密码子使用偏好性;(3)毛曼青冈和曼青冈亲缘关系如何。以期从叶绿体基因组层面为栎属青冈亚属系统发育问题解决、种间关系的划分、种的鉴定及优良青冈木材的选育提供新资料。
1 材料与方法
1.1 实验材料
在野外采集宁冈青冈(Quercus ningangensis)、曼青冈(Q. oxyodon)、毛曼青冈(Q. gambleana)、竹叶青冈(Q. neglecta)新鲜的、无病虫害的成熟叶片(表 1),用湿润的纱布擦拭后置于装有变色硅胶的密封袋中保存,干燥后置于-80 ℃的冰箱内备用。凭证标本存放于西华师范大学标本馆(Herbarium, College of Life Sciences, China West Normal University)。
1.2 基因组DNA的提取与测序
用改良的CTAB法从4种栎属青冈亚属植物叶片中提取总DNA。利用Illumina Hiseq对提取的DNA进行双端测序。叶绿体全基因组DNA的提取与测序均由南京奥维森基因科技有限公司完成。
1.3 组装与注释
利用FASTQ软件进行数据质控,将质控得到的Clean data利用GetOrganelle软件(Jin et al., 2020)进行组装,将原始数据map到组装好的序列上查看覆盖率并结合Gapcloser软件(Zuo et al., 2017),补齐组装出现的空缺(gaps)。在NCBI (https://www.ncbi.nlm.nih.gov/)上下载褐叶青冈(MN199023)的叶绿体基因组作为参考基因组,利用CPGAVAS2(Shi et al., 2019)(http://www.herbalgenomics.org/cpgavas/)进行注释,注释好的数据利用Geneious軟件(Kearse et al., 2012)进行手动调整和修改。利用OGDRAW软件(https://chlorobox.mpimp-golm.mpg.de/OGDraw.html)绘制4种栎属青冈亚属植物的叶绿体基因组物理结构图谱。最后将注释好的数据提交到NCBI,宁冈青冈、曼青冈、毛曼青冈、竹叶青冈的序列登录号分别为ON303301、ON258628、ON258629、ON258631。
1.4 叶绿体基因组比较分析
利用Geneious(Kearse et al., 2012)及Editseq软件统计4种栎属青冈亚属植物的叶绿体基因组序列SSC、LSC、IRS区域的长度、GC含量等基本信息。从NCBI上下载小叶青冈(MN199025)、褐叶青冈(MN199023)、西畴青冈(NC_036941)、青冈(NC_036930)的叶绿体基因组与本文4种栎属青冈亚属植物所测的叶绿体基因组进行比较分析。
选择以ATG为起始密码子、长度大于300 pb的非重复序列,所有物种筛选后均剩余52条CDS序列。利用Codon W软件(Sharp & Li, 1987)计算4种栎属青冈亚属植物及其近缘种的同义密码子相对使用度(relative synonymous codon usage, RSCU)、密码子适应指数(codon adaptation index, CAI)、有效密码子数(effective number of codons, ENC)、密码子偏好性指数(codon bias index, CBI)、最优密码子使用频率(frequency of optimal codons, FOP)以及每条CDS序列的ENC值、RSCU值及密码子第三位各碱基出现的概率(记为A3、T3、C3、G3),运用在线软件EMBOSS(https://www.bioinformatics.nl/emboss-explorer/)计算各CDS序列GC1、GC2、GC3的含量。中性绘图分析以GC1和GC2的平均值(GC12)为纵坐标,以GC3值为横坐标,绘制散点图,并插入y=x的函数图像。ENC-plot分析绘制的散点图将GC3值、ENC值分别作为横、纵坐标,添加理论ENC值曲线,标准曲线公式为ENC=2+GC3+29/ [GC32+(1-GC3)2]。PR2-plot分析以G3/(G3+C3)为横坐标,A3/(A3+T3)为纵坐标绘制散点图,分析A、T、C、G的使用频率和偏倚性。中性绘图、ENC-plot绘图、PR2-plot绘图均使用Origin软件。
利用mVISTA软件(https://genome.lbl.gov/vista/mvista/instructions.shtml)(Mayor et al., 2000)对4种栎属青冈亚属植物及其近缘种的叶绿体基因组进行可视化对比分析。使用IRscope(https://irscope.shinyapps.io/irapp/)(Amiryousefi et al., 2018)绘制宁冈青冈、曼青冈、毛曼青冈、竹叶青冈及其近缘种的叶绿体基因组IR边界。
1.5 系统发育分析
选取并下载在NCBI中公开释放的8种壳斗科植物的叶绿体基因组序列,包括倒卵叶青冈(Q. obovatifolia,MG356785.1)、川滇高山栎(Q. aquifolioides, KX911971.1)、刺叶高山栎(Q. spinosa,MG678038.1)、巴东栎(Q. engleriana, MZ196209.1)、栓皮栎(Q. variabilis, NC031356.1)、三棱栎(Trigonobalanus doichangensis, NC023959.1)、台湾水青冈(Fagus hayatae,MW846258.1)、米心水青冈(F. engleriana,NC036929.1)。选用台湾水青冈和米心水青冈作为外类群,将上述8种壳斗科植物的叶绿体基因组与本文组装的4种栎属青冈亚属植物的叶绿体基因组一起构建系统发育树。将所有叶绿体基因组序列运用MAFFT(Katoh & Standley, 2013)进行对比,将对比好的叶绿体基因组序列利用MEGA 7软件(Kumar et al., 2016)对序列开头和结尾未对齐部分进行适当的修剪,运用MEGA 7软件中Models功能寻找以上序列运用ML法构建系统发育树的最佳模型(GTR+GI),以最大似然法(maximum likelihood,ML),设置bootstrap为1 000构建系统发育树。另将修剪好的序列再运用MEGA 7软件以邻接法(neighbor-joining method, NJ),选用Maximum Composite Likelihood模型,设置bootstrap为10 000构建相关物种的系统发育树。
2 结果与分析
2.1 叶绿体基因组的基本特性
宁冈青冈、曼青冈、毛曼青冈、竹叶青冈的叶绿体基因组均有由一个LSC区域、两个IRs区域、一个SSC区域构成,为典型的四分体结构(图 1)。其基因组长度为160 681~160 906 bp,其中曼青冈和毛曼青冈叶绿体全基因组的长度分别为160 883 bp和160 882 bp,仅相差1个碱基。LSC区的长度范围为90 245~90 360 bp,SSC区的长度范围为18 891~18 929 bp,IRs的长度范围为25 816~25 840 bp。本实验测得的4种青冈的总GC含量(36.9%)、IRs区域的GC含量(42.8%)及CDs区域的GC含量(37.9%)均一致,LSC区域及SSC区域的GC含量也相差不大(表 2)。
宁冈青冈、曼青冈、毛曼青冈、竹叶青冈的叶绿体基因组注释结果显示,4种栎属青冈亚属植物均有133个基因,其中包含37个tRNA基因(transfer RNA genes),8个rRNA基因(ribosomal RNA genes),以及88个蛋白质编码基因(protein-coding genes),(表 2)。有15个基因有1个内含子(rpoC1、ndhA、ndhB、rpl2、rpl16、atpF、rps16、trnL-UAA、trnK-UUU、trnI-GAU、trnA-UGC、trnG-GCC、trnV-UAC、petB、petD),包含2個内含子的基因有3个(rps12、clpP、ycf3)(表3)。
2.2 密码子偏好性分析
2.2.1 密码子组成分析 4种栎属青冈亚属植物的密码子数在20 996~20 977之间;有效密码子数(effective number of codons, ENC)在49.91~49.81之间,密码子适应指数(codon adaptation index, CAI)均为0.167;宁冈青冈、毛曼青冈和竹叶青冈的密码子偏好性指数(codon bias index, CBI)为-0.099,曼青冈的密码子偏好性指数为-0.097;4种栎属青冈亚属植物的最优密码子使用频率(frequency of optimal codons, FOP)均为0.355,GC含量在37.93%~37.95%之间。4种栎属青冈亚属圆圈内外的灰色箭头表示基因转录方向;不同的颜色代表不同功能的基因;内圈深灰色表示GC含量,浅灰色表示AT含量。
Gray arrows inside and outside the circle indicate the direction of gene transcription; Different colors represent genes with different functions; Dark gray in the inner circle represents GC content, the light gray represents AT content.
如圖2所示,亮氨酸(Leu)、丝氨酸(Ser)精氨酸(Arg)有6个同义密码子,色氨酸(Trp)、甲硫氨酸(Met)仅由一种密码子编码,其余氨基酸均由2个及以上的同义密码子编码。这8种栎属青冈亚属植物的密码子偏好性相似,其中ACU、UCU、UUA、GCU、UAU、GAU、AGA的使用频率较高,而CUC、CUG、GCG、UAC、CAC、CAG、AAC、AAG、GAC、GAG、CGC、CGG、AGC、GGC的使用频率较低。在米槠(Castanopsis carlesii)、长果柯(Lithocarpus longinux)中UAA的使用频率较所列的其他植物低。
2.2.2 中性绘图分析 从4种栎属青冈亚属植物叶绿体基因组筛选出以ATG为起始密码子、长度大于300 pb的非重复序列进行中性绘图,结果(图 3)显示,宁冈青冈、曼青冈、毛曼青冈、竹叶青冈GC3和GC12的相关系数分别为0.01、0.05、0.01、0.06,回归系数分别为0.12、0.34、0.34、0.36,双尾检验结果均为P<0.01,说明GC12和GC3之间的相关性较弱,组成密码子碱基的第1、第2位和第3位之间存在显著差异。由此推测,自然选择对4种栎属青冈亚属植物叶绿体基因组密码子的使用偏好性的影响更大。
2.2.3 ENC-plot绘图分析 由图4结果显示,大部分基因的实际ENC值在标准曲线之下,说明自然选择对这部分基因的密码子使用偏好性的影响更大;少部分基因的实际ENC值在标准曲线附近或上方,说明突变对其密码子的使用偏好性影响更大。ENC比值频率分布(表 5)显示,宁冈青冈、曼青冈、毛曼青冈、竹叶青冈中位于-0.05~0.05区间的基因分别有19、18、18、19个,分别占37%、35%、35%、37%,表明突变对这些基因的影响较大。这4种栎属青冈亚属植物中有33~34个基因的实际ENC值与理论ENC值差距较大,这部分基因受自然选择的影响作用较大。ENC-plot分析结果和ENC比值频率分布显示,4种栎属青冈亚属植物的叶绿体基因组密码子偏好性虽同时受突变和自然选择的影响,但总体而言,自然选择的影响大于突变带来的影响。
2.2.4 PR2-plot绘图分析 由图5可知,分布于中心点上或附近的基因主要受突变的影响, 而大多远离中心的点则受自然选择的影响。4种栎属青冈亚属植物的PR2-plot绘图分析显示, 基因分布不均匀,大部分基因分布于右下角,说明4种栎属青冈亚属植物叶绿体基因组中密码子第3位碱基的使用频率为A>T,G>C,表明4种栎属青冈亚属植物密码子的偏好性受到了突变和自然选择的共同影响。
2.3 叶绿体基因组差异比较分析
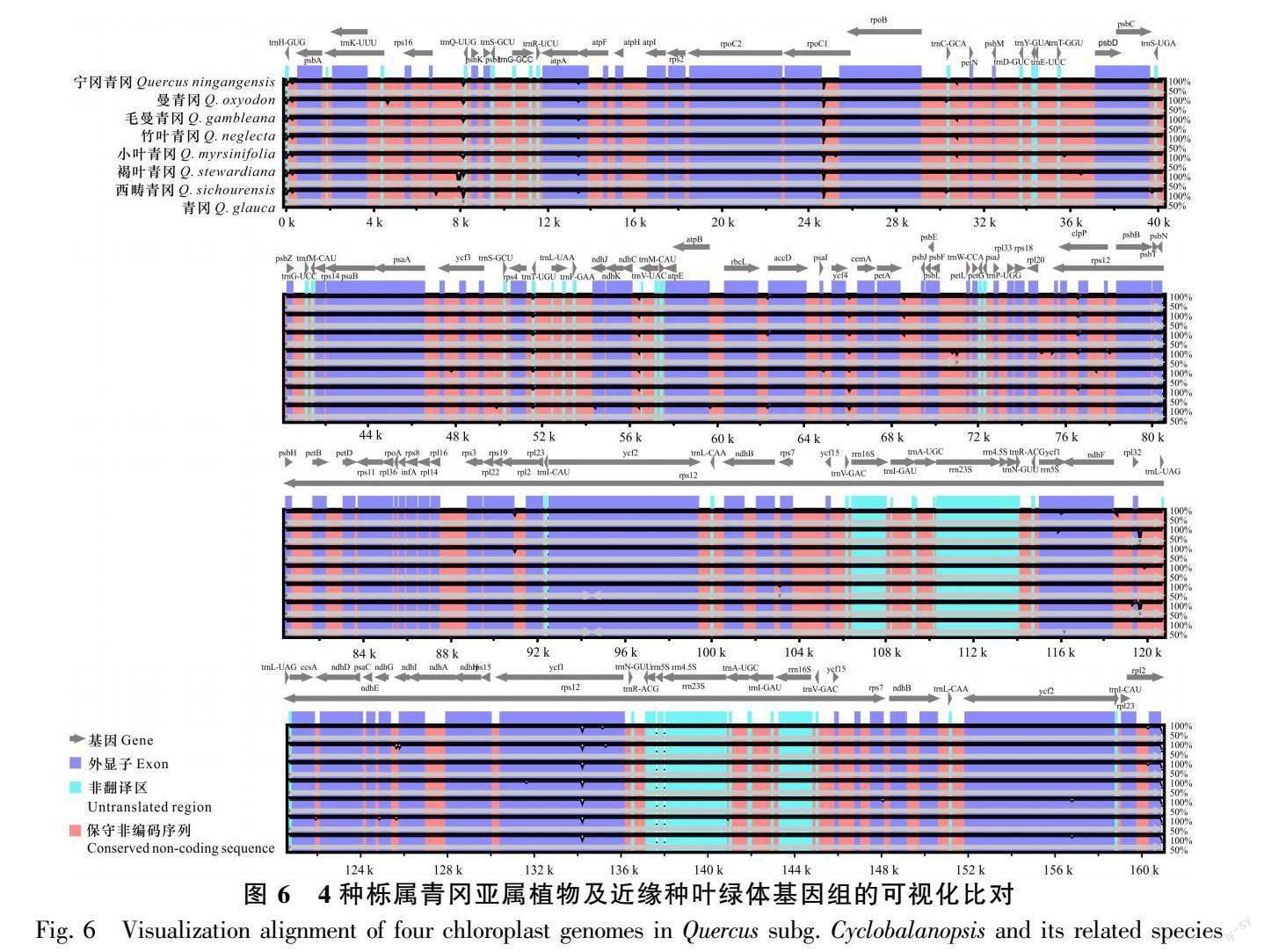
选宁冈青冈的叶绿体基因组做参考,利用mVISTA对曼青冈、毛曼青冈、竹叶青冈、小叶青冈、褐叶青冈、西畴青冈、青冈的叶绿体基因组序列进行可视化比较分析。从分析结果可以看出,栎属青冈亚属植物的叶绿体基因组具有较高的保守性,主要的变异区域位于非编码区,而ycf1基因在蛋白质编码区域发生变异较大。LSC区域的变异较大,IRs区域变异小较为保守,SSC区域的变异小于LSC区域。8种栎属青冈亚属植物的rRNA基因高度保守,几乎没有发生变异(图6)。
2.4 IR边界分析
使用在线软件IRscope分析4种栎属青冈亚属植物及其近缘种叶绿体基因组的IR扩张和收缩情况,以及识别LSC、IR、SSC的边界位置(图7)。在8种青冈中,各边界附近分布的基因一致,但距边界的距离略有不同。其中,rps19基因距SSC/IRb边界均为11 bp, 西畴青冈和青冈的trnH白色和红色分别表示较低和较高的 RSCU 值。毛曼青冈、曼青冈、竹叶青冈、宁冈青冈为本研究组装和注释,其余物种均为NCBI上下载。
White and red indicate lower and higher RSCU values, respectively. Quercus gambleana, Q. oxyodon, Q. neglecta and Q. ningangensis were assembled and annotated for the study, and the remaining species were downloaded from NCBI.
基因距离IRa/LSC边界的距离为16 bp,其余青冈为1 bp。在毛曼青冈和竹叶青冈中ndhF基因,有一个碱基位于IRb区域,其余2 255 bp均位于SSC区域,其余青冈中ndhF基因的长度为2 255 bp且均位于SSC区域。高等植物的叶绿体基因组中ycf1基因为第二长的基因,其横跨这8种青冈叶绿体基因组的SSC/IRa边界,在宁冈青冈、小叶青冈、褐叶青冈、西畴青冈、青冈中,ycf1基因有1 060 bp位于IRa区域,在曼青冈、毛曼青冈、竹叶青冈中分别有1 056、1 054、1 062 bp位于IRa区域,在8种青冈的IRb区域也分别拷贝了相应长度的ycf1片段,这部分被视为ycf1假基因(ψycf1)。8种青冈中,ψycf1基因横跨IRb/SSC,有56~58 bp位于IRb/SSC上游。
2.5 系统发育分析
选取NCBI上公开释放的8种壳斗科植物的叶绿体基因组与本研究组装的4种栎属青冈亚属植物的叶绿体基因组序列,以台湾水青冈(Fagus hayatae)和米心水青冈(F. engleriana)作为外类群,分别用ML法和NJ法构建系统发育树,来探讨栎属青冈亚属和栎亚属之间的关系及毛曼青冈和曼青冈的关系,并为栎属青冈亚属系统发育问题的解决提供基础资料。两种方法构建的系统发育树均得到了较高的支持率且获得的拓扑结构一致。系统发育树(图 8)显示,水青冈属(Fagus)和轮叶三棱栎属(Formanodendron)位于系统发育树的基部,为壳斗科中较早分化出来的类群。曼青冈和毛曼青冈成姐妹群关系,毛曼青冈为4种栎属青冈亚属植物中最早分化出来的类群。栎亚属并未形成一个单系群,川滇高山栎(Quercus aquifolioides)与栎属青冈亚属的亲缘关系较近。
3 討论与结论
3.1 叶绿体基因组比较及密码子偏好性分析
被子植物的叶绿体全基因组长度在120 000~170 000 bp之间( Tangphatsornruang et al., 2010),本研究中宁冈青冈、曼青冈、毛曼青冈、竹叶青冈的叶绿体基因组长度为160 784~160 906 bp,在叶绿体基因组的大小、结构、基因的数量、总GC含量及4个区域的长度和GC含量上均高度保守。栎属青冈亚属的叶绿体基因组基本特征与其他壳斗科植物相似,壳斗科与其他科物种相比基因密度更大,对环境的适应更强,稳定性更高(高潇潇, 2020)。从4种栎属青冈亚属植物的叶绿体全基因组比较分析可以看出,4种栎属青冈亚属植物的叶绿体基因组高度保守,变异区域主要在非编码区。栎属青冈亚属植物的鉴定主要依据壳斗的特征,在没有果实的情况下,部分种难以鉴定,可以利用叶绿体基因组筛选变异较大的区域,开发成DNA条形码,便于栎属青冈亚属植物的鉴定。通过对8种栎属青冈亚属植物叶绿体基因组的研究,筛选出trnH-GUG~trnQ-UUG、ropC1~petN、trnT-UGC~trnM-CAU、ycf4~psbJ及ycf1区域作为DNA条形码,为后续栎属青冈亚属植物的鉴定提供便利。被子植物叶绿体基因组IR区域的扩张和收缩普遍存在于植物进化过程中(Hansen et al., 2007;Davis & Soreng, 2010;Huang et al., 2014)。栎属青冈亚属植物位于IR区域的基因相同,说明栎属青冈亚属IR边界较为保守。4种栎属青冈亚属植物均出现ycf1假基因(ψycf1),在壳斗科栎亚属,栗属(Castanea)部分种中也出现了ycf1基因在IRb/SSC拷贝为假基因的现象(杨颜慈, 2018;高潇潇, 2020)。
密码子使用偏好性是生物进化的重要现象。主要受基因组的大小、碱基突变、基因漂变、自然选择、基因的表达水平、tRNA丰度、GC含量、蛋白质结构等因素影响(Romero et al., 2000;Duret , 2000;Angellotti et al., 2007)。从GC3的含量可以看出,本研究中的栎属青冈亚属植物更偏好以A/T结尾的密码子,这一现象在被子植物中广泛存在(Clegg et al., 1994;Tangphatsornruang et al., 2010;Delannoy et al., 2011)。同义密码子相对使用度结果显示,栎属青冈亚属和栎亚属植物的同义密码子使用度相似,米槠、长果柯中UAA的使用频率较所列的其他植物低,说明壳斗科植物的密码子使用偏好性相似,而不同的属之间仍存在着一定的差异。3种绘图分析结果显示,4种栎属青冈亚属植物的密码子偏好性受选择压力和突变同时影响, 但选择压力为主要影响因素, 这与米槠(江淑珍等,2021)、降香黄檀(Dalbergia odorifera)(原晓龙等, 2021)、灯盏花(Erigeron breviscapus)(李显煌等,2021)等得出的结果相似。
3.2 系统发育关系及种间关系分析
系统发育树结果显示,水青冈属和三棱栎属分化较早,与周浙昆(1999)根据化石和地理信息系统对壳斗科起源等研究的结果一致。前人根据花粉特征及核基因将川滇高山栎、刺叶高山栎、巴东栎归为Ilex组、栓皮栎归为Cerris组(Hubert F et al., 2014),而核标记、质体标记的研究结果中显示,Ilex组的亚洲种常出现穿插在Cerris组中的现象(Simeone et al., 2013;Hubert et al., 2014)。本研究中,4种栎亚属植物并未形成一个单系群,刺叶高山栎(Quercus spinosa)与巴东栎(Q. engleriana)、栓皮栎(Q. variabilis)聚在一枝,Ilex组和Cerris组之间出现穿插的现象,与核标记、质体标记的结果一致。在《中国植物志》(陈焕镛和黄成就, 1998)、《中国树木志》(郑万钧, 1985)、《云南植物志》(中国科学院昆明植物研究所, 1979)、《贵州植物志》 (蓝开敏和李永康, 1982)、 等地方植物志中,毛曼青冈和曼青冈是作为两个单独的种处理(罗艳和周浙昆,2001a)。邓敏(2007)等根据曼青冈、毛曼青冈叶背的星状毛等特征,认为毛曼青冈应作为曼青冈的一个变种处理,而刘凌燕等(2008)根据聚类分析的结果认为应将毛曼青冈作为长叶青冈(Q. longifolia)的变种处理。本研究中系统发育分析结果显示,曼青冈与竹叶青冈亲缘关系较近,曼青冈和毛曼青冈未形成姐妹群。虽然毛曼青冈和曼青冈的果实和叶型相似,但是叶背毛的颜色和形态存在差异,并且在不同地区采集的叶背毛差距很大(邓敏, 2007)。在扫描电镜下观察两者的花粉外壁纹饰纹饰上也存在区别,毛曼青冈的花粉外壁纹饰为颗粒疣状或聚合疣状,曼青冈的花粉外壁为颗粒状纹饰(王萍莉和溥发鼎, 2004)。本研究中采的毛曼青冈叶密被黄褐色星状毛,而曼青冈叶背的星状毛为白色且较毛曼青冈稀疏。综上,本研究更倾向于毛曼青冈和曼青冈作为两个种处理的观点。本研究中的毛曼青冈采自重庆,曼青冈采自广西,两地相距较远,推测可能由于生长环境存在较大差异及长期的基因交流受阻等因素导致本研究中的毛曼青冈和曼青冈逐渐形成两个种。
参考文献:
AMIRYOUSEFI A, HYVNEN J, POCZAI P, 2018. IRscope: an online program to visualize the junction sites of chloroplast genomes [J]. Bioinformatics, 34(17): 3030-3031.
ANGELLOTTI MC, BHUIYAN SB, CHEN GR et al., 2007. CodonO: codon usage bias analysis within and across genomes [J]. Nucl Acid Res, 35(Suppl. 2): W132-W136.
CHEN HY, HUANG CJ, 1998. Flora Reipublicae Popularis Sinicae: Tomus 22[M]. Beijing: Science Press: 263-334.[陈焕镛, 黄成就, 1998. 中国植物志: 第二十二卷 [M]. 北京: 科学出版社: 263-334]
CLEGG MT, GAUT BS, LEARN GH et al., 1994. Rates and patterns of chloroplast DNA evolution [J]. Proc Nat Acad Sci, 91(15): 6795-6801.
DAVIS JI, SORENG RJ, 2010. Migration of endpoints of two genes relative to boundaries between regions of the plastid genome in the grass family (Poaceae) [J]. Am J Bot, 97(5): 874-892.
DELANNOY E, FUJII S, COLAS DES FRANCS-SMALL C et al., 2011. Rampant gene loss in the underground orchid Rhizanthella gardneri highlights evolutionary constraints on plastid genomes [J]. Molec Biol Evol, 28(7): 2077-2086.
DENG M, 2007. Anatomy, taxonomy, distribution & phylogeny of Quercus subg. Cyclobalanopsis (Oersted) Schneid. (Fagaceae) [D]. Beijing: Chinese Academy of Sciences: 3-70.[邓敏, 2007. 壳斗科栎属青冈亚属的形态解剖、分类、分布及其系统演化 [D]. 北京: 中国科学院研究生院: 3-70.]
DUAN MX, 1992. Studies on the Fagaceae wood-assortment in Taiwan [J]. Fujian Sci Technol, 19(4): 65-70.[端木炘, 1992. 台湾壳斗科木材分类的研究 [J]. 福建林业科技, 19(4): 65-70.]
DUAN MX, 1995. The comprehensive utilization of the resources of Cyclobalanopsis in China [J]. J Beijing For Univ, 17(2): 109-110.[端木炘, 1995. 我国青冈属资源的综合利用 [J]. 北京林业大学学报, 17(2): 109-110.]
DURET L, 2000. tRNA gene number and codon usage in the C. elegans genome are co-adapted for optimal translation of highly expressed genes [J]. Trends Genet, 16(7): 287-289.
FRODIN DG, GOVAERTS R, 1998.World Checklist and Bibliography of Fagales (Betulaceae, Corylaceae, Fagaceae and Ticodendraceae) [M]. London: The Royal Botanic Gardens: 201-394.
GAO XX, 2020. Comparative whole chloroplast genomics of Castanea and Fagales family [D]. Xian: Northwest UNIVERSITY: 14-29.[高瀟潇, 2020. 壳斗目栗属植物比较叶绿体基因组与系统发育研究 [D]. 西安: 西北大学: 14-29.]
HANSEN DR, DASTIDAR SG, CAI ZQ, et al., 2007. Phylogenetic and evolutionary implications of complete chloroplast genome sequences of four early-diverging angiosperms: Buxus (Buxaceae), Chloranthus (Chloran-thaceae), Dioscorea (Dioscoreaceae), and Illicium (Schi-sandraceae) [J]. Mol Phylogenet Evol, 45(2): 547-563.
HUANG CJ, ZHANG YT, BRUCE B, 1999. Flora of China Vol. 4 [M]. Beijing: Science Press and Missouri Botanical Garden Press: 314-400.
HUANG H, SHI C, LIU Y et al., 2014. Thirteen Camellia chloroplast genome sequences determined by high-throughput sequencing: genome structure and phylogenetic relationships [J]. BMC Evol Biol, 14(1): 1-17.
HUBERT F, GRIMM GW, JOUSSELIN E et al., 2014. Multiple nuclear genes stabilize the phylogenetic backbone of the genus Quercus [J]. Syst Biodivers, 12(4): 405-423.
Institute of Botany Institute of Botany, The Chinese Academy of Sciences, 1972. Iconographia Cormophytorum Sinnicorum: Tomus 1 [M]. 北京: Science Press: 439-462. [中国科学院植物研究所, 1972. 中国高等植物图鉴:第一册 [M]. 北京: 科学出版社: 439-462.]
JIANG SZ, LIAN H, XIONG YF, et.al., 2021. Analysis of codon bias in chloroplast genome of Castanopsis carlesii [J/OL]. Mol Plant Breed: 1-12[2022-04-28]. http://kns.cnki.net/kcms/detail/46.1068.S.20210722.1131.006.html.[江淑珍, 连辉, 熊远芳等., 2021. 米槠叶绿体基因组密码子偏好性分析 [J/OL]. 分子植物育种: 1-12[2022-04-28]. http://kns.cnki.net/kcms/detail/46.1068.S.20210722.1131.006.html.]
JIN JJ, YU WB, YANG JB et al., 2020. GetOrganelle: a fast and versatile toolkit for accurate de novo assembly of organelle genomes[J]. Genome Biol, 21(1): 1-31.
KATOH K, STANDLEY DM, 2013. MAFFT multiple sequence alignment software version 7: improvements in performance and usability [J]. Mol Biol Evol, 30(4): 772-780.
KEARSE M, MOIR R, WILSON A et al., 2012. Geneious Basic: An integrated and extendable desktop software platform for the organization and analysis of sequence data [J]. Bioinformatics, 28(12): 1647-1649.
KORPELAINEN H, 2004. The evolutionary processes of mitochondrial and chloroplast genomes differ from those of nuclear genomes[J]. Naturwissenschaften, 91(11): 505-518.
KUMAR S, STECHER G, TAMURA K, 2016. MEGA7: molecular evolutionary genetics analysis version 7.0 for bigger datasets [J]. Mol Biol Evol, 33(7): 1870-1874.
Kunming Institute of Botany, Chinese Academy of Sciences, 1979. Flora Yunnanica: Tomus 2 (spermatophyte) [M]. Beijing: Science Press: 322-325.[中國科学院昆明植物研究所, 1979. 云南植物志:第二卷(种子植物) [M]. 北京: 科学出版社: 322-325]
KWAK SY, LEW TTS, SWEENEY CJ et al., 2019. Chloroplast-selective gene delivery and expression in planta using chitosan-complexed single-walled carbon nanotube carriers [J]. Nat Nanotechnol, 14(5): 447-455.
LAN KM, LI YK, 1982. Guizhou botanical chronicles [M]. Guiyang: Guizhou Peoples Press: 103-105.[蓝开敏, 李永康, 1982. 贵州植物志 [M]. 贵阳: 贵州人民出版社: 103-105]
LIU LF, FANG ZJ, 1986. A study on pollen morphology of Quercoideae oerst (Fagaceae) in China [J]. Guihaia, 12(4): 243-251.[刘兰芳, 房志坚, 1986. 中国壳斗科栎亚科花粉形态研究 [J]. 广西植物, 12(4): 243-251.]
LIU LY, ZHANG ML, LI JQ, et.al., 2008. A numerical taxonomic study of the genus Cyclobalanopsis oersted from China [J]. Wuhan Botan Res, 26(5): 466-475.[刘凌燕, 张明理, 李建强, 等, 2008. 国产青冈属的数量分类学研究 [J]. 武汉植物学研究, 26(5): 466-475.]
LI XH, YANG SC, XIN YX, et.al., 2021. Analysis of the codon usage bias of chloroplast genome in Erigeron breviscapus (Vant.) Hand-Mazz [J]. J Yunnan Agr Univ(Nat Sci Ed), 36(3): 384-392.[李显煌, 杨生超, 辛雅萱, 等, 2021. 灯盏花叶绿体基因组密码子偏好性分析 [J]. 云南农业大学学报(自然科学), 36(3): 384-392.]
LUO Y, ZHOU ZK, 2001a. Leaf epidermis of Quercus subgen. Cyclobalanopsis (Oerst.) Schneid. (fagaceae) [J]. Acta Phytotax Sin, 39(6): 489-501.[罗艳, 周浙昆, 2001a. 栎属青冈亚属(壳斗科)的叶表皮研究[J]. 植物分类学报, 39(6): 489-501.]
LUO Y, ZHOU ZK, 2001b. Phytogeography of Quercus subg. Cyclobalanopsis [J]. Acta Bot Yunnan, 23(1): 1-16.[罗艳, 周浙昆, 2001b. 青冈亚属植物的地理分布 [J]. 云南植物研究, 23(1): 1-16.]
MAYOR C, BRUDNO M, SCHWARTZ JR et al., 2000. VISTA: visualizing global DNA sequence alignments of arbitrary length [J]. Bioinformatics, 16(11): 1046-1047.
ROMERO H, ZAVALA A, MUSTO H, 2000. Codon usage in Chlamydia trachomatis is the result of strand-specific mutational biases and a complex pattern of selective forces [J]. Nucl Acid Res, 28(10): 2084-2090.
SHARP PM, LI WH, 1987. The codon adaptation index — a measure of directional synonymous codon usage bias, and its potential applications [J]. Nucl Acid Res, 15(3): 1281-1295.
SHI LC, CHEN HM, JIANG M et al., 2019. CPGAVAS2, an integrated plastome sequence annotator and analyzer [J]. Nucl Acid Res, 47(W1): W65-W73.
SIMEONE MC, PIREDDA R, PAPINI A et al., 2013. Application of plastid and nuclear markers to DNA barcoding of Euro-Mediterranean oaks (Quercus, Fagaceae): problems, prospects and phylogenetic implications [J]. Bot J Linn Soc, 172(4): 478-499.
TANGPHATSORNRUANG S, SANGSRAKRU D, CHANPRA-SERT J, et al., 2010. The chloroplast genome sequence of mungbean (Vigna radiata) determined by high-throughput pyrosequencing: structural organization and phylogenetic relationships [J]. DNA Res, 17(1): 11-22.
WANG PL, PU DF, 2004. Pollen morphology and biogeography of Fagaceae [M]. Guangzhou: Guangdong Sci Technol Press: 1-158.[王萍莉, 溥發鼎, 2004. 壳斗科植物花粉形态及生物地理 [M]. 广州: 广东科技出版社: 1-158.]
XU YC, REN XW, 1976. The classification and distribution of Fagaceae of Yunnan Province (2) [J]. J Syst Evol, 14(2): 73-88.[徐永椿, 任宪威, 1976. 云南壳斗科分类与分布(2) [J]. 植物分类学报, 14(2): 73-88.]
YANG YC, 2018. Plastid genomes of Quercus in China and the key genera in Fagaceae: comparative and phylogenetic analyses [D]. Xian: Northwest University: 19-90.[杨颜慈, 2018. 中国栎属植物和壳斗科主要属质体基因组比较分析和系统发育研究 [D]. 西安: 西北大学: 19-90.]
YUAN XL, LI YQ, ZHANG JF, et.al., 2021. Analysis of codon usage bias in the chloroplast genome of Dalbergia odorifera [J]. Guihaia, 41(4): 622-630.[原晓龙, 李云琴, 张劲峰, 等, 2021. 降香黄檀叶绿体基因组密码子偏好性分析 [J]. 广西植物, 41(4): 622-630.]
ZHAO ZY, WANG X,YU Y et al., 2018. Complete chloroplast genome sequences of Dioscorea: Characterization, genomic resources, and phylogenetic analyses [J]. PeerJ, 6(12): e6032.
ZHENG WJ, 1985. Sylva Sinica: Vol. 2[M]. Beijing: China Forestry Publishing House: 2199-2356.[郑万钧, 1985. 中国树木志 第2卷 [M]. 北京: 中国林业出版社: 2199-2356]
ZHOU ZK, 1999. Fossils of the Fagaceae and their implications in systematics and biogeography [J]. Acta Phytotax Sin, 37(4): 369-385.[周浙昆, 1999. 壳斗科的地质历史及其系统学和植物地理学意义[J]. 植物分类学报, 37(4): 369-385.]
ZUO LH, SHANG AQ, ZHANG S et al., 2017. The first complete chloroplast genome sequences of Ulmus species by de novo sequencing: Genome comparative and taxonomic position analysis [J]. PLoS ONE, 12(2): e0171264.
(责任编辑 李 莉 王登惠)
