
雌雄株香榧生育期光合特性与氮分配关系研究
2023-05-19 01:57:14欧明烛任启飞马菁华陈云飞任少华
种子 2023年2期
欧明烛, 任启飞, 马菁华, 刘 芳, 郑 听, 陈云飞, 任少华, 杨 朔, 周 艳
(贵州省植物园, 贵阳 550004)
香榧(Torreyagrandis)隶属红豆杉科(Taxaceae)榧属,常绿乔木、雌雄异株,是榧树(Torreyagrandis)经过人工选种繁育嫁接的中国特有珍稀干果树种,同时也是集药用、油用和观赏于一体的经济树种。大多数果树和经济林木的产量与植物性别性状有密切联系,不同性别植物具有不同的经济价值,原因在于雄性器官和雌性器官的区别,其中雄性植株在绿化林木中具有较大的经济价值,雌性植株则在种子和果实类植物中具有较大的经济价值[1]。因此,探究香榧雌雄性植株叶片在生育期生理生化指标之间的关系,对栽培优质香榧并提高产量具有重要意义。目前对香榧雌雄异株植物的叶片研究集中在光合特性变化、氮分配及氮利用效率等方面。当年生春梢叶片及结果枝叶片是果实发育的重要同化物供应源,二者对果实发育都具有重要作用[2-3]。由于雌雄异株植物对雄性和雌性生殖有不同需求,这类植物的资源分配模式可能因性别而产生差异[4]。光合有效辐射是影响早实核桃和晚实核桃净光合速率的主要环境因素,在生产中需注意株行距、适当修剪等[5]。植物的光合作用影响植物的生长发育,叶片氮含量影响叶片中光合色素含量,与光合作用关系密切。叶片氮分配是指氮在植物叶片各细胞结构及游离化合物中的分配比例,叶片氮分配方式对叶片光合能力的强弱起决定性作用[6]。光合氮利用效率(PNUE)是描述植物叶片生理特性、营养成分利用及生存策略的重要指标,在生产中提高光合氮利用效率能提高作物的氮素利用率和产量[7-8]。刘琏等[9]研究表明,无论树龄大小,从5月到8月,叶片氮含量减少,干物质积累使叶片发育成熟,8月到11月氮分配比例发生变化导致叶片PNUE相对较高。黄增冠等[10]从香榧不同叶龄的叶片,比较其光合能力及氮分配之间的关系,不同叶龄的光合特性有差异,叶龄增加使叶片对光适应性减弱,叶龄相同不同叶位香榧叶片的光合特性无显著差异。陈佳妮等[11]对香榧和榧树进行叶片光合特性比较,并研究两者之间的光保护机制,与榧树相比,香榧叶片虽具有较高的氮含量,但光合氮素利用效率低。宋洋等[12]研究表明,适当遮荫处理能提高香榧苗对弱光的利用率,促进香榧苗的生长发育,而过度遮荫可能导致光合能力下降。近些年来针对香榧的研究主要集中在药用、化学成分、香榧种子和假种皮等方面[13-15],缺乏对雌雄香榧的花、叶等部位之间关系的研究,尤其对雌雄香榧叶片光合特性及氮分配之间的比较研究相对较少。本研究通过对雌雄香榧营养生长阶段(无花、无果)与生殖生长阶段(开花、结果)叶片光合特性及叶片氮含量、氮分配等测定,初步探讨香榧不同生长阶段叶片光合特性的差异及氮分配调节机制,对改善香榧叶片光合性能,为进一步提高香榧果实产量提供理论依据。
1 材料与方法
1.1 试验材料
试验地位于贵州省植物园香榧良种采穗圃,地理位置(106°42′E,26°34′N),海拔1 210~1 410 m,年均气温15.3 ℃,相对湿度约为74%,年平均总降水量为1 174.7 mm,日照率低,气候温暖湿润,热量充足,雨量充沛,属于典型的北亚热带高原季风湿润气候区,具有典型高原山地喀斯特地貌特征[16]。供试材料为长势良好且一致的香榧幼树嫁接苗(2014年2+5嫁接苗,砧木为二年生榧树,穗条为五年生香榧),所有植株树高、胸径和冠幅基本一致。选择雌雄植株侧枝冠层阳面的健康成熟一年生无果(无花)(N+1)-WG(WH)叶片及一年生结果(开花)(N+1)-GG(KH)叶片作为测试叶,分别挂牌标记,测定时保持叶片自然生长角度不变,每株植物重复测定3次,测定时间为2020年10月中旬。
1.2 测定项目及方法
1.2.1光合-光响应曲线
采用Li-6800便携式光合仪(美国LI-COR公司)于晴天的09:00—12:00时,14:00—17:00时自然环境条件下测定香榧幼树结果(开花)和无果(无花)叶片的光响应曲线。测定时,设定叶室温度25 ℃,叶室相对湿度50%,气体流速为500 μmol/s,根据外界环境的实际光强设置人工光源2100RGB适应20 min,CO2浓度为400 μmol/mol,由CO2小钢瓶控制CO2浓度,光合有效辐射强度(PAR)梯度为2 100,1 800,1 500,1 200,900,700,500,400,300,200,100,50,20,10,0 μmol/(m2·s)。在每个梯度的光强下稳定4 min记录各项数据,重复3~4次,取测定结果的平均值。测定的主要参数包括:胞间CO2浓度(Ci),蒸腾速率(Tr),最大叶片净光合速率(Pmax),气孔导度(Gs)等。
根据SPSS软件计算出光响应曲线光饱和点(LSP)、光补偿点(LCP)、最大净光合速率(Pmax)、暗呼吸速率(Rd)。

表1 香榧幼树生长环境因子Table 1 The growth environmental factors of Torreya grandis young trees
1.2.2CO2响应曲线
测定条件与测定光响应曲线的条件相同,对选择光响应曲线测定的叶片进行充分足够的光合诱导,控制光强为1 800 μmol/(m2·s),CO2浓度为400 μmol/mol诱导20 min,CO2浓度设置为400、300、200、100、50、5、400、400、600、800、1 000、1 200、1 500、1 800 μmol/mol,测定参数:叶片净光合速率Pn[μmol/(m2·s)],根据CO2响应曲线求得[17],最大羧化速率Vcmax[μmol/(m2·s)]和最大光合电子传递速率Jmax[μmol/(m2·s)]。
1.2.3光合色素含量
称取0.02 g香榧幼树结果(开花)和无果(无花)鲜叶片,把叶片放入盛有8 mL提取液(95%乙醇)的离心管中密封遮光浸提约24 h。用紫外分光光度计(UV 754 PC,上海佑科)检测波长在664、649、470 nm下的吸光度,计算叶绿素含量用单位面积表示与光合速率一致,计算方法参考文献[18]进行。
1.2.4比叶重
全面提升基层党员干部的统战工作能力和水平,必须实现党员干部教育培训机制与统战教育培训体系的无缝对接,构建系统完善、科学管用的领导干部统战教育培训体系。系统的教育培训,既解决思想认识不到位的问题,又解决工作方法不专业的问题。
测定气体交换参数后,取下测定的叶片用植物图像分析仪(LA-S,杭州万深)测定叶面积,即用透明胶带将测定枝条上的叶片全部剪下粘贴于白纸上进行扫描,用软件计算测定叶面积。将叶片于85 ℃烘干至恒质量,称量干质量。
比叶质量(g/m2)=干质量/叶面积。
1.2.5叶片氮素含量及叶片光合器官中氮分配的计算
叶氮含量测定参考扩散法,扩散皿内圈盛硼酸溶液,外圈放待测液后加入氢氧化钠溶液,密封,产生的氨扩散在皿内,由硼酸溶液吸收,直接用标准酸测定。以饱和最大光合速率(Pmax)与叶氮含量(NA)的比值表示光合氮利用效率(PNUE)[19]。
PUNE=Pmax/NA。
参与叶片光合的氮分配阶段分为羧化系统(参与暗反应,主要为Rubisco)、类囊体(参与暗反应),其中类囊体细分为生物力能学组分与捕光系统(即①PSⅠ、②PSⅡ和③LHCⅡ),各组分氮素分配比例计算公式[20]如下:
(1)
(2)
(3)
PT=PC+PB+PL
(4)
式中,PC为分配到羧化系统中的氮素的比例;PB为分配到生物力能学组分氮素的比例;PL为分配到捕光系统氮素的比例;Vcr为Rubisco的比活度(单位Rubisco蛋白的CO2固定活性值[μmol/(g·s)]);NA为单位面积叶氮含量;Jmc为单位Cyt f的最大电子传递速率[μmol/(m2·s)];Cc为叶片叶绿素浓度(mmol/g);CB为捕光系统(PSI、PSII、LHCII)叶绿素与氮的比值(mmol/g);6.25为氮素转换成Rubisco蛋白的系数;8.06为氮素与单位Cyt f之间的转换系数。
2 结果与分析
2.1 光合特性指标及光合光响应曲线
如图1所示,无论是香榧雌树一年生无果叶片(C-N+1)-WG或雄树一年生无花叶片(X-N+1)-WH,雌树一年生挂果叶片(C-N+1)-GG或雄树一年生开花叶片(X-N+1)-KH对PAR的光响应曲线基本一致。当PAR<416[μmol/(m2·s)]时,Pn随PAR的增强迅速增大,此时PAR是香榧幼树叶片光合能力强弱的主要限制因素;当PAR>416[μmol/(m2·s)]时,Pn随PAR的增强,其上升速度开始减小并趋于平缓,最后达到光饱和状态。采用非直角双曲线模型对光合光响应曲线进行拟合,得到主要光合特性参数指标,结果见表2。雄树一年生无花叶片Pmax,Rd显著小于雌树一年生无果叶片(p<0.05),雌雄一年生无果、无花叶片与一年生挂果、开花叶片LSP无明显差异(p<0.05),雌雄一年生无果、无花叶片LCP无明显差异(p<0.05),雄树一年生开花叶片LCP显著小于雌树一年生挂果叶片(p<0.05)。雌雄一年生挂果、开花叶片Pmax,Rd,LSP,LCP均高于雌雄一年生无果、无花叶片。

表2 雌雄香榧幼树结果(开花)与无果(无花)叶片光响应曲线特征参数Table 2 Characteristic parameters of light response curves of leaves of female and male Torreya grandis young tree with fruit (blossom) and fruitless (no blossom)
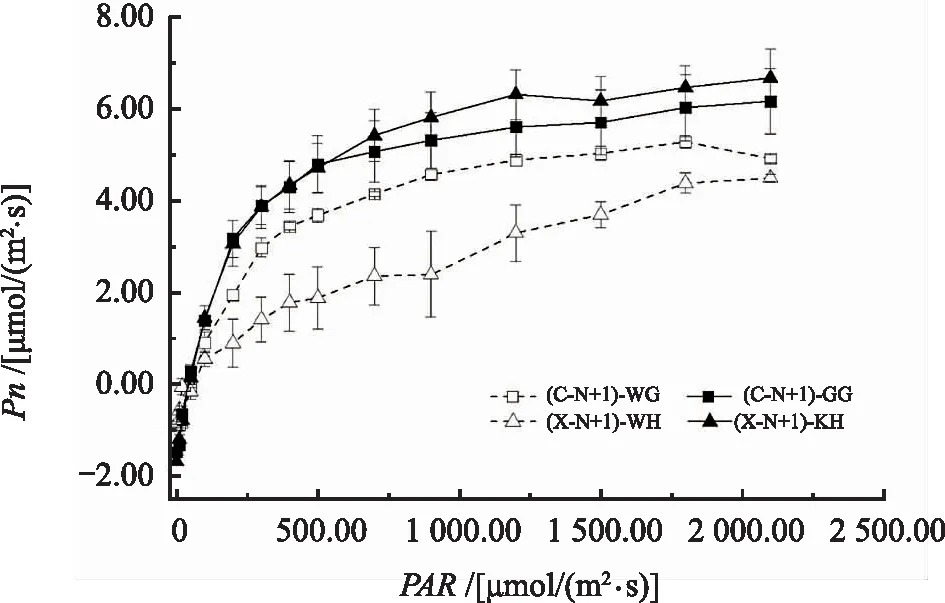
注:(C-N+1)-WG为雌树一年生无果叶片;(C-N+1)-GG为雌树一年生挂果叶片;(X-N+1)-WH为雄树一年生无花叶片;(X-N+1)-KH为雄树一年生开花叶片。图1 雌雄香榧幼树结果(开花)与无果(无花)叶片光响应曲线Fig.1 Photosynthetic response curves of leaves in female and male Torreya grandis young tree with fruit (blossom) and fruitless (no blossom)
2.2 光合CO2响应曲线
由表3可知,雌雄一年生无果、无花叶片与一年生挂果、开花叶片的Vcmax呈上升趋势,分别上升3.15%和15.79%。雌雄一年生无果、无花叶片与一年生挂果、开花叶片的Jmax呈上升趋势,分别上升28.19%和9.45%。雌雄叶片之间的Vcmax、Jmax无明显差异(p<0.05),除了雌树一年生挂果叶片的Jmax显著高于雄树一年生开花叶片之外;雌树一年生无果叶片的Jmax显著低于一年生挂果叶片,雄树一年生无花叶片与一年生开花叶片无明显差异(p<0.05)。

表3 雌雄香榧幼树结果(开花)与无果(无花)叶片CO2响应曲线模拟的光合参数Table 3 The modeled photosynthetic parameters in response to intercellular CO2 concentration in leaves of female and male Torreya grandis young tree with fruit (blossom) and fruitless (no blossom)
2.3 光合色素含量与最大净光合速率的关系
由表4可知,雌雄一年生无果、无花叶片与一年生挂果、开花叶片的Car含量相比呈上升趋势,分别上升3.70%和11.11%,雌雄一年生无果、无花叶片与一年生挂果、开花叶片的Chl(a+b)含量相比呈上升趋势,分别上升8.00%和12.24%。雌树一年生无果叶片与一年生挂果叶片的Car及Chl(a+b)含量之间无明显差异(p<0.05),雄树一年生无花叶片的Car及Chl(a+b)含量显著小于一年生开花叶片(p<0.05),雌雄一年生无果、无花叶片的Chl(a/b)显著小于一年生挂果、开花叶片(p<0.05),雌树Chl(a/b)显著高于雄树(p<0.05)。雌雄香榧叶片的最大光合净速率与总叶绿素含量之间呈显著性正相关(图2)。
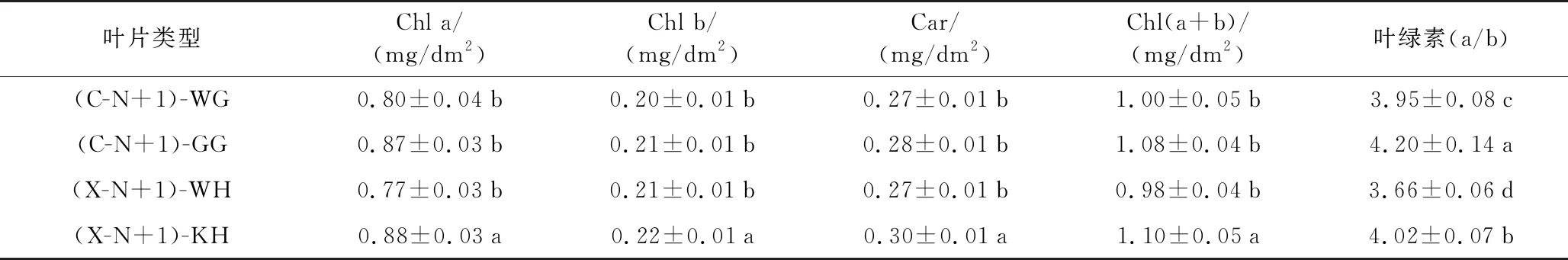
表4 雌雄香榧幼树结果(开花)与无果(无花)叶片光合色素含量Table 4 The photosynthetic pigment contents in leaves of female and male Torreya grandis young tree with fruit (blossom) and fruitless (no blossom)
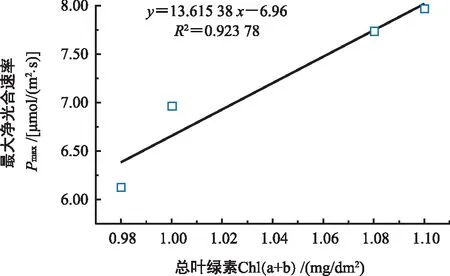
图2 香榧叶片总叶绿素(Chl(a+b))与最大净光合速率(Pmax)之间的相关性Fig.2 The relationship of total chlorophyll content(Chl(a+b)) and the maximum net photosynthetic rate(Pmax) in Torreya grandis
2.4 比叶质量
如图3,雌树叶片的SLW显著高于雄树叶片(p<0.05),而雌树一年生无果与一年生挂果叶片的SLW之间无显著差异,雄树一年生无花叶片的SLW显著高于一年生开花叶片(p<0.05)。
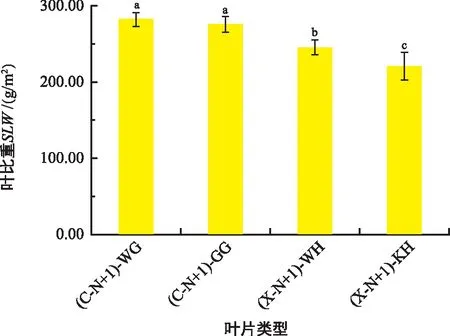
图3 雌雄香榧幼树结果(开花)与无果(无花)叶片SLW的变化Fig.3 SLW changes of female and male Torreya grandis young tree with fruit (blossom) and fruitless (no blossom)
2.5 叶氮含量及分配的关系
由表5可知,雌雄一年生无果、无花叶片与一年生挂果、开花叶片的叶氮含量NA呈上升趋势,分别上升30.45%和27.80%。雌雄一年生无果、无花叶片的NA,PC,PB,PL,PT都显著低于一年生挂果、开花叶片(p<0.05),除了雄树一年生无花叶片与一年生开花叶片的PC之间无显著差异之外(p<0.05);各叶片类型的NA,PB,PL之间无显著差异(p<0.05),雄树叶片的PC,PT都显著高于雌树叶片(p<0.05)。
2.6 光合氮素利用效率
如图4,雌树一年生挂果叶片的PNUE显著高于雄树一年生无花叶片及一年生开花叶片,雄树一年生无花叶片的PNUE显著高于雌树一年生无果叶片(p<0.05)。
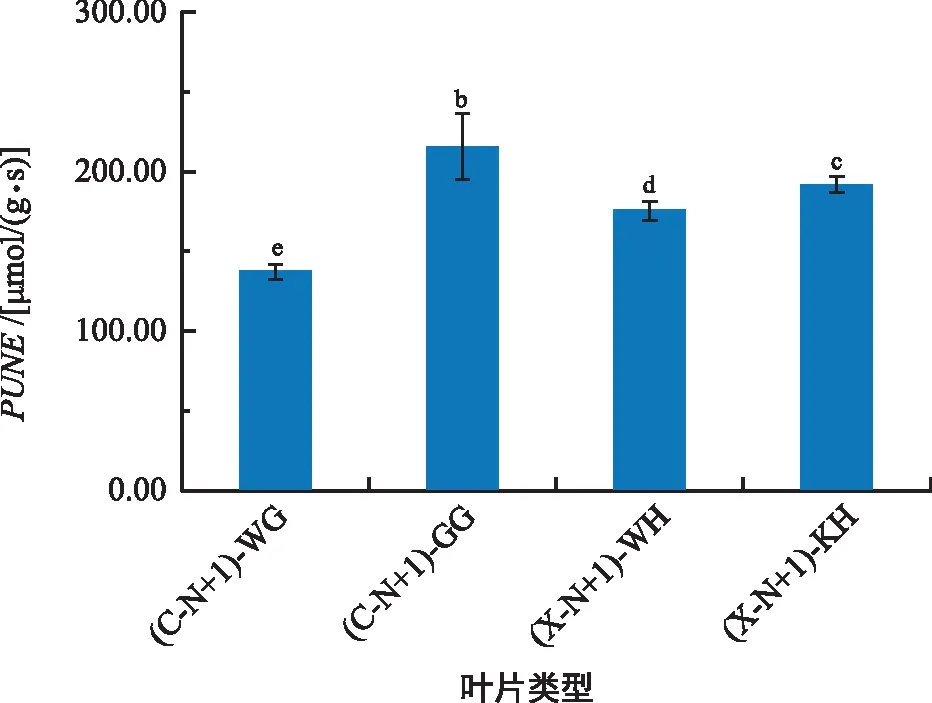
图4 雌雄香榧幼树结果(开花)与无果(无花)叶片PNUEFig.4 PNUE of female and male Torreya grandis young tree with fruit (blossom) and fruitless (no blossom)
3 讨论与结论
叶片是植物进行光合作用的重要器官,叶片性状特征能直接影响植物的生长发育水平及果实的产量和品质。最大净光合速率(Pmax)是衡量叶片光合能力的重要指标[21]。提高地石榴叶片的净光合速率可以增强对光能的利用率[22]。柴仲平等[23]研究表明,不同生长期香梨光合特性有明显差异,膨果期香梨叶片净光合速率、叶绿素含量、叶面积指数、蒸腾速率、气孔限制值和气孔导度都比坐果期高。本研究发现,雌雄香榧幼树一年生挂果、开花叶片的最大净光合速率(Pmax)高于一年生无果、无花叶片,表明光合作用与果实的形成呈正相关。
叶片的光饱和点和光补偿点反映了植物对光照条件的要求,是判断植物耐阴性的重要指标。一般阴性植物的LSP为500~1 000[μmol/(m2·s)]或者更低,而LCP小于20[μmol/(m2·s)],一般阳性植物的LSP在1 500~2 000[(μmol/(m2·s)]之间或者更高,而LCP在50~100[μmol/(m2·s)]之间。本研究结果显示,香榧叶片的LSP为563.60~391.34[μmol/(m2·s)],LCP为60.71~35.90[μmol/(m2·s)],符合阴生植物光饱和点与光补偿点的范围,雌雄香榧幼树一年生挂果、开花叶片的LSP值较高,表明生殖生长阶段光合能力强,同时也对强光的适应能力强(表2)。
植物叶片中最大羧化速率与叶氮含量、Rubisco含量有显著相关性[24]。最大羧化速率与叶氮分配给羧化系统的比例对光合速率起着决定性作用[25-26]。随着叶龄增加,光合速率降低是由于其叶氮含量和Rubisco/Chl比值低造成的[27]。本研究结果表明,雌雄香榧幼树一年生挂果、开花叶片的Vcmax、Jmax高于一年生无果、无花叶片,可能是由于无果枝和无花枝的叶片会优先将各类营养元素提供给结果枝和开花枝的叶片,比如氮素等元素,通过光合作用更有利于干物质的积累,所以导致一年生挂果、开花叶片的Vcmax、Jmax高于一年生无果、无花叶片。
叶片光合能力与光合色素含量、酶活性等有关。不同叶位的叶绿素Chl(a+b)含量与净光合速率之间均呈显著正相关[28]。本研究结果与其一致,一年生挂果、开花叶片的光合色素含量及Pmax均高于一年生无果、无花叶片,即光合色素与Pmax呈正相关。
比叶重(SLW)能反映植物对资源的利用方式[29],大多数研究认为叶片光合作用能力和叶氮含量都与SLW有关[24]。若植物叶片细胞壁的氮素分配增加,那么分配到叶绿体的氮素就会减少[30]。本研究结果显示,雌雄香榧幼树一年生无果、无花叶片的SLW高于一年生挂果、开花叶片(图3),这可能是结果枝和开花枝中的叶片氮素分配到光合机构大于分配到叶片细胞壁所导致一年生无果、无花叶片的SLW较高,也可能是导致香榧不同类型叶片的PNUE产生差异的主要原因。有研究表明,植物增加CO2扩散的阻力和分配到非光合组分的氮比率以及减少叶片透射光的比率,使PNUE降低,导致PNUE较低的植物通常会具有较高的SLW[31,8]。本研究结果显示,雌雄香榧幼树一年生无果、无花叶片的PNUE显著低于一年生挂果、开花叶片(图4),与SLW呈负相关。
许多研究表明,光合能力与植物叶片氮含量呈显著正相关[32-33],因为通常叶氮含量中大约有一半氮素会分配到光合机构中,比如Calvin循环和类囊体相关的酶[34-37]。研究发现,成熟后的叶片氮含量和光合速率逐渐降低[38]。本研究结果显示,雌雄香榧幼树一年生无果、无花叶片的叶氮含量(NA)及PC,PB,PT显著低于一年生挂果、开花叶片(表3),说明分配到结果枝和开花枝叶片的光合机构中氮含量较高,可能是氮元素含量高低直接影响光合作用强弱。
本研究中,雌雄香榧幼树在生育期(已开花结果枝)的叶片最大净光合速率(Pmax)、LSP、Vcmax、Jmax、Pmax、PNUE及叶氮含量值均高于未生育期(未开花结果枝),说明了光合作用与果实的形成呈正相关关系,光合色素与Pmax也呈正相关关系,所以在生殖生长阶段光合能力及对强光的适应能力强,而无果枝和无花枝的叶片会优先将各类营养元素提供给结果枝和开花枝的叶片,比如氮素等元素,而分配到结果枝和开花枝叶片的光合机构中氮含量较高,可能是氮元素含量高低直接影响光合作用强弱,通过光合作用更有利于干物质的积累。因此,在香榧生产过程中不仅要注意种植地中雌雄株的性配比,还应适当增加株行距及修剪,保持透光通风,合理利用有效光辐射,也要注意氮肥的施用。
猜你喜欢
恋爱婚姻家庭(2023年1期)2023-02-15 13:02:32
今日畜牧兽医(2022年10期)2022-12-23 06:22:28
今日农业(2022年1期)2022-11-16 21:20:05
辽河(2020年8期)2020-09-02 14:25:25
基层中医药(2018年8期)2018-11-10 05:32:08
浙江林业科技(2018年6期)2018-03-13 02:56:50
现代园艺(2018年3期)2018-02-10 05:18:21
绿色中国(2017年19期)2017-05-25 13:25:14
浙江柑橘(2016年3期)2016-03-11 20:12:51
Coco薇(2015年12期)2015-12-10 02:40:50
