
胡萝卜果胶杆菌侵染下半夏转录组中SSR位点信息分析
2023-05-19 01:57:14舒福兴王馨瑶袁碧霞保正娟罗清清陈集双
种子 2023年2期
舒福兴, 黄 瑶, 贾 启, 王馨瑶, 唐 滢,袁碧霞, 保正娟, 罗清清, 陈集双
(1.遵义医科大学生物资源健康利用研究中心, 贵州 遵义 563000;2.遵义医科大学医学与科技学院临床学院, 贵州 遵义 563000)
半夏(Pinelliaternata(Thunb.)Breit)属于天南星科半夏属草本植物,具有止咳、降逆止呕、抗肿瘤等作用[1]。半夏主要分布于南亚地区,在中国主要分布于贵州、甘肃、河北等地[2]。由于半夏原生环境持续恶化以及被无节制挖掘,致使野生半夏资源减少,品种、数量均急剧下降。尽管多地进行了半夏人工种植,但面积依然不够大,并且面临半夏软腐病引起的绝收。研究表明,多种致病菌可引起半夏软腐病,其中胡萝卜果胶杆菌(Pectobacteriumcarotovorum)最常见、致病性强,接种20 h即可引起软腐病症状[3]。因此,筛选半夏软腐病抗病品种显得尤为重要。而获得软腐病抗病半夏品种首先得找到可以鉴定抗病品种的辅助育种方法。
简单重复序列SSR(Simple Sequence Repeat),又称微卫星标记,是一种广泛分布于真核和原核生物基因组中,由1~6个核苷酸组成的串联重复序列,其两侧是相同的基因组序列,但在非基因区域更频繁地出现。其具有多态性高、重复性高、覆盖面广等优点,广泛运用于遗传多样性、分子标记辅助育种[4],尤其是在许多作物中,选择数量性状位点QTL作图的标记方法长期以来一直是SSR[5]。SSR的来源常用的有两种:基因组SSR、转录组SSR(EST-SSR)[6-7]。最初的SSR标记引物开发需要构建DNA文库,该过程步骤繁琐、失败率高、耗资大、工作量大[8]。EST-SSR则摈弃繁琐的DNA建库过程,通过转录组开发标记引物,能够直接反应参与某一进程转录基因信息。目前,NCBI中收录的半夏EST序列较少,基于软腐病菌侵染的半夏转录组信息更少,难以满足SSR引物开发的需要。因此,本研究通过挖掘软腐病菌胡萝卜果胶杆菌侵染本团队特有的T 2+半夏品种,以获得此条件下的半夏转录组中SSR位点信息,为鉴别半夏软腐病抗病品种提供依据。
1 材料与方法
1.1 实验材料
实验材料为组培条件下胡萝卜果胶杆菌侵染的半夏嫩芽。胡萝卜果胶杆菌为本团队保存,菌样甘油保存于-80 ℃;T 2+半夏品种保留于遵义医科大学生物资源健康利用研究中心组培室,培养条件为光照强度2 000 lx,光照时间12 h/d,培养温度(25±1)℃。
1.2 实验方法
在恒温摇床中,利用LB液体培养基培养胡萝卜果胶杆菌;培养温度为28 ℃,转速为150 r/min;在波长为600 nm下测量菌液OD值,到0.6时停止培养,取出摇床待用。采用同一批接种、长势均一的半夏组培苗作为接种对象。实验设置0 h和20 h实验组,并在相应时间点设对照组。试验组用半夏组培苗与5 mL胡萝卜果胶杆菌培养液共培养,每组重复3次;对照组采用半夏组培苗与5 mL LB培养基共培养,每组重复3次。培养条件为光照强度2 000 lx,光照时间12 h/d,培养温度(25±1)℃。分别采集各组半夏嫩芽立即投入液氮中速冻,利用提前液氮冷冻的冻存管装好,埋在干冰中送往山东开源基因公司进行cDNA文库构建及llumina HiSeq X Ten平台测序获得转录组数据。获取的转录组数据采用MISA(http://pgrc.ipk-gatersleben.de/misa/)生物信息学SSR位点分析软件对转录组序列数据(Unigenes)进行检索和分析。参数设置参考蒋维昕等[9]的方法。
2 结果与分析
2.1 半夏转录组中SSR位点的数量
通过对半夏转录组中75 517条Unigene(总长度90 921 938 bp)进行SSR分析获得40 621个SSR,出现频率为36.09%,分布于27 252条Unigene,这些Unigene的数量为总Unigenes数量的36.09%。其中,含有2个及以上SSR位点的Unigene有8 916条。含有复合型SSR位点的Unigene有4 969条。半夏转录组中,平均每2.24 kb序列就会出现1个SSR位点,详见表1。

表1 半夏转录组中SSR基本信息Table 1 Basic information of SSR in the transcriptome of Pinellia ternata
2.2 半夏转录组中SSR位点的重复基元类型及占比
半夏转录组具有单核苷酸至六核苷酸等多种SSR重复基元种类。不同SSR重复基元类型在数量上差异悬殊,基元类型较多的是单、二、三核苷酸重复。其中,二核苷酸重复类型有16 936个,数量最多,占总SSR数量的41.69%;单核苷酸重复类型有13 293个,排第二位,占总SSR数量的32.72%;三核苷酸重复类型有9 829个,排第三位,占总SSR数量的24.20%;其余基元重复类型所占比例较低,三者占总SSR数量之和的1.39%。总体上,含SSR位点的序列数量随着核苷酸基元碱基数的增加逐渐变少。二核苷酸重复类型SSR的平均分布距离最小,为5.37 kb,五核苷酸重复类型SSR的平均分布距离最大,为1 466.48 kb,是二核苷酸重复类型平均分布距离的273.09倍,详见表2。
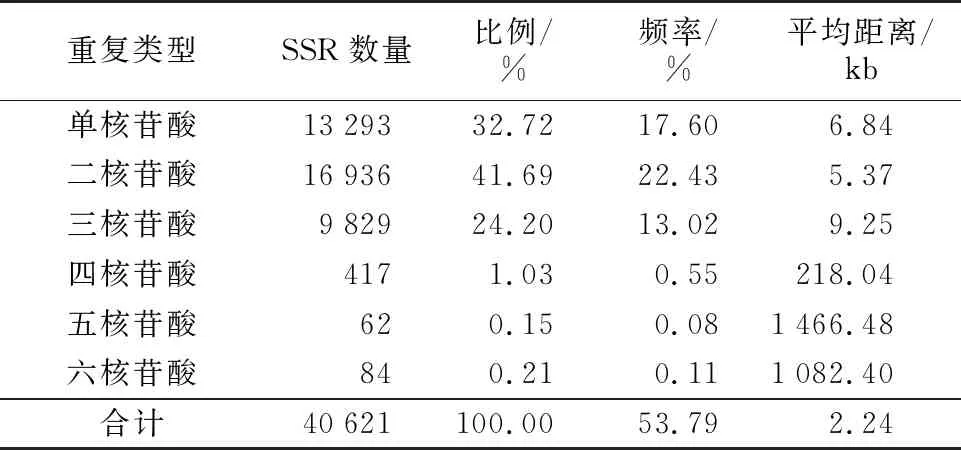
表2 半夏转录组中SSR的数量及重复基元类型Table 2 Type and repeat motifs of SSR loci in the transcriptome of Pinellia ternata
2.3 半夏转录组中SSR重复基元的碱基组成
分析发现,有40 621个SSR位点存在于半夏转录组中。这些SSR中一共发现106种重复基元类型。其中,单核苷酸至六核苷酸重复基元分别有2、4、10、25、28、37种。在所有SSR基元重复类型中,以AG/CT为碱基的基元重复类型为优势类群,占总SSR的30.93%;其次为单核苷酸重复基元,该重复类型中A/T基元占总SSR的29.72%。三核苷酸基元类型中,优势重复基元及占总SSR的比例分别为:CCG/CGG占比6.65%、AAG/CCT占比4.60%、AGG/CCT占比4.01%、AAG/CTT占比4.60%、其余四、五、六及其他核苷酸重复基元种类也比较丰富,但全部加起来仅占19.49%(详见图1)。
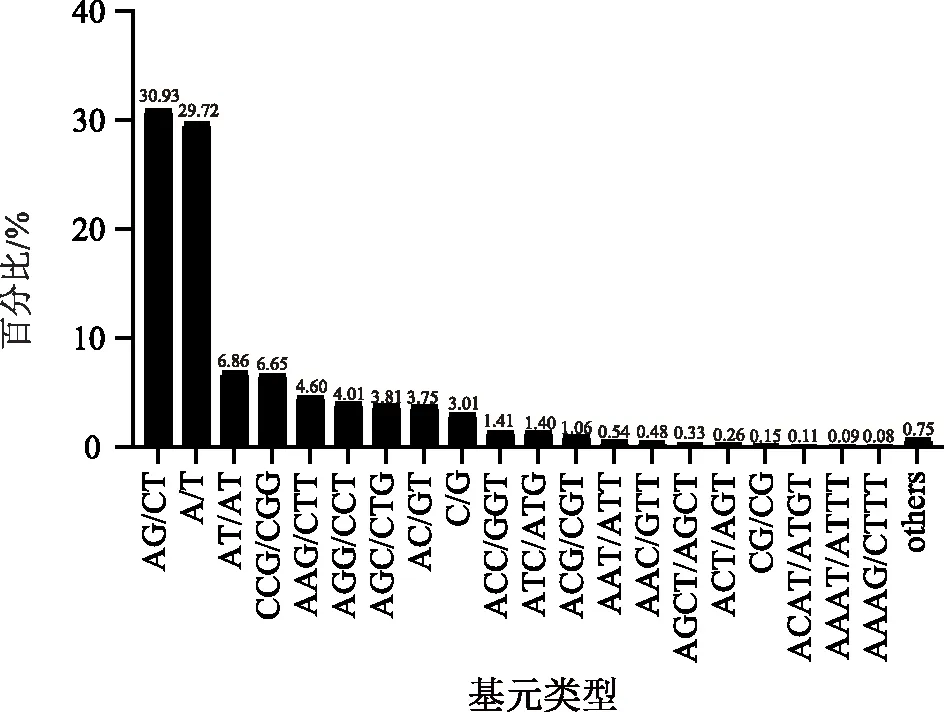
图1 半夏转录组中不同重复基元SSR的占比情况Fig.1 Percentage of SSRs with different repeats motifs in the transcriptome of Pinellia ternata
2.4 半夏转录组中SSR的多态性分析
2.4.1半夏转录组序列中SSR基元重复次数的分布
分析半夏转录组SSR位点的重复次数发现,基元重复次数的增加使不同类型核苷酸重复基元的数量和比例逐渐下降(图2和表3)。有37 008个SSR重复单元的重复次数集中在5~20次,占总SSR数量的91.11%(表3)。其中,有25 478个低重复率(5~10次)SSR位点,数量为总SSR数量的62.72%;中重复次数中,11~15次的SSR位点数为9 350个,占总SSR数量的23.02%,16~20次的SSR位点数为2 180个,占总SSR数量的5.37%。在SSR所有重复类型中,占比排列前三的依次为10次、6次、5次重复类型,分别有6 550、5 949、5 522个,各占SSR总数的16.14%、14.65%、13.59%。
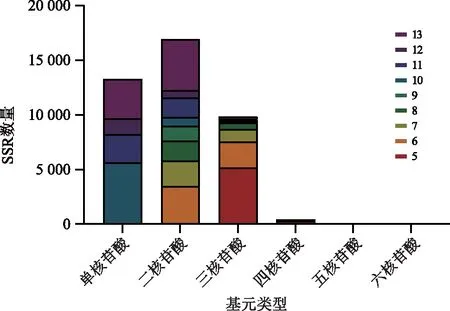
注:不同颜色代表不同重复单元次数,带有颜色的方框面积越大对应的SSR数量越多。图2 半夏转录组SSR重复类型统计Fig.2 Statistics of the SSR repeat types of the transcriptome of Pinellia ternata
2.4.2半夏转录组高多态性SSR分析
研究认为,具有较高多态性的SSR序列的长度往往大于等于20 bp,具有这一特点的SSR可以成为首选标记位点;多态性适中的序列长度在12~20 bp之间;而SSR序列长度一旦小于12 bp,则多态性表现极低[10]。因此,本研究以此为依据分析高多态性的SSR数量及占比(图3)。结果发现,有12 521个SSR序列长度集中于12~20 bp,占SSR总数的30.82%;有4 006个SSR序列长度处于20~30 bp,占SSR总数的9.86%。 有3 754个SSR序列长度超过30 bp,占10.53%;共计有20.39%的SSR序列长度符合高多态性SSR的特征。此外,半夏转录组SSR序列的平均长度为2.24 kb,其中G/C单核苷酸类型具有最长重复片段为66 bp,其出现3次。
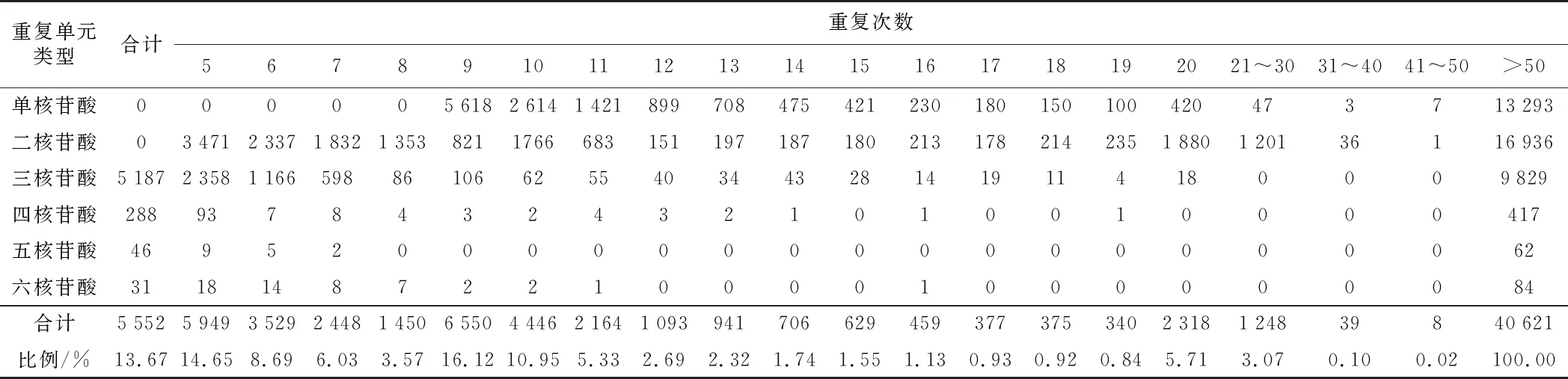
表3 半夏转录组不同重复次数的SSR数量分布Table 3 SSR quantity distribution with different repetitions in transcriptome of Pinellia ternata
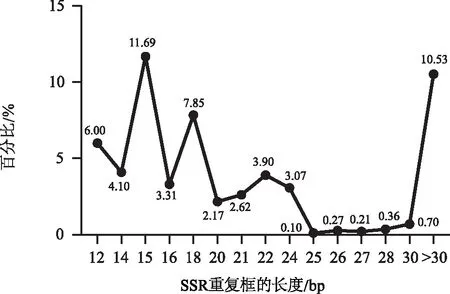
图3 半夏转录组不同长度SSR重复框的占比Fig.3 Proportion of SSR repeat fragments of different lengths in the transcriptome of Pinellia ternata
3 讨 论
不同品种半夏呈现出的软腐病抗病性有极大差异,有的表现为延迟发病,有的表现为易感病状态,还有的表现为抗病状态。目前,唐晓晶[11]用ISSR-PCR方法鉴定半夏近缘混淆品种及多来源品种;张君毅[12]、杨旻[13]利用ISSR分子标记技术分析半夏遗传多样性;上述寻找SSR的方法比较传统,过程繁琐信息不够全面。之后,马芃锐等[14]采用磁珠富集法获得芍药叶型半夏基因组序列841条,分析出SSR 406条,筛选出可以鉴定4种叶型半夏品种的引物7对。然而,在SSR挖掘数量以及可用性方面,磁珠法远不如转录组法。随着转录组测序技术的成熟,多种植物已经成功将其用于SSR开发[15-17]。因此,基于转录组信息分析半夏SSR分子标记具有可行性。
在以往的研究中,已有基于正常半夏转录组获得SSR分子标记的研究,例如王森等[15]在正常转录组中发现14 468个SSR,分布于12 000条Unigene 中;刘丹等[16]在正常转录组发现半夏中含有SSR位点的Unigene有14 640条,有19 270个SSR位点;这为半夏品种的鉴定打下基础。然而,这些基于非软腐病菌胁迫下的半夏转录组分析而来的SSR用于半夏软腐病抗病品种筛选具有很大缺点,例如: 1) 非软腐病抗病相关的SSR在筛选软腐病抗病品种过程中加大了分析工作量,甚至导致软腐病抗病品种筛选的失败; 2) 半夏未被软腐病菌侵染时,某些抗病基因可能未发生转录,那么使用这种转录组数据进行SSR分析,可能会造成部分抗病相关的SSR遗失。为克服这些缺点,本研究利用软腐病致病菌(胡萝卜果胶杆菌)侵染半夏T 2+,并获得半夏T 2+的转录组信息,在此基础上进一步分析SSR信息。结果发现,在27 252条Unigene中找到40 621个SSR位点,同时比较T 2+正常转录组中Unigene的数量(107 777个)发现[2],T 2+半夏在胡萝卜果胶杆菌侵染下,转录组中的Unigene数量显著降至75 517个,因此,利用本转录组筛选SSR用于抗软腐病半夏品种育种将更具针对性及必要性;尽管本次T 2+半夏转录组Unigene数量显著下降,但其SSR数量远高于王森等[15]和刘丹等[16]报道的SSR数量,这说明半夏品系之间转录组水平有极大的差异性,对其他半夏品种的EST-SSR研究也有必要。
本研究中,T 2+半夏转录组SSR基元重复类型种类多,出现的密度大,频率高。在27 252条Unigene中找到符合条件的SSR位点40 621个,Unigene出现频率为36.09%,这个频率远高于王森等[15]报道的13.47%和刘丹等[16]报道的17.06%;也高于同属天南星科的魔芋17.91%[18],造成这一差异的原因一方面可能是来自种属之间的差异;另一方面可能是转录组收集到了正常转录组未收集到的含有SSR的Unigene。本研究中,SSR的平均距离为2.24 kb,高于王森等[15]报道的4.33 kb及刘丹等[16]报道的3.39 kb,说明本研究发现的SSR作为分子标记具有很大潜力。
半夏转录组中以单核苷酸、二核苷酸、三核苷酸重复类型为SSR主要基元类型,这与刘丹等[16]报道的趋势一致。SSR基元重复次数、序列片段长度决定了SSR的多态性。随着SSR基元重复次数的增加,尤其是重复次数大于等于12次或者序列长度大于等于20 bp时将其开发为多态性标记的潜力将大大提升[19]。本研究中的SSR重复单元次数主要集中在5~20次之间,占比约为91.11%,其中重复次数大于等于12次的SSR占总SSR的比例为26.33%,表明这些SSR位点具有较大开发潜力。
总之,本研究利用胡萝卜果胶杆菌侵染下的T 2+半夏转录组筛选出的SSR重复基元类型多,出现频率高、密度大、其作为多态性标记开发的潜能大。这些SSR标记将缩短半夏抗软腐病品种选育的时间、增加半夏多样性研究以及遗传图谱绘制的手段。
猜你喜欢
农技服务(2023年11期)2023-11-27 12:54:30
高中数理化(2023年6期)2023-08-26 13:28:24
烟台大学学报(自然科学与工程版)(2021年1期)2021-03-19 08:39:24
中国果树(2020年2期)2020-07-25 02:14:24
中成药(2018年6期)2018-07-11 03:01:12
科学导报(2018年30期)2018-05-14 12:06:01
现代园艺(2017年13期)2018-01-19 02:27:58
中国粮油学报(2016年1期)2016-02-06 02:17:06
长江蔬菜(2014年22期)2014-12-08 08:57:12
Chinese Journal of Chemical Engineering(2014年3期)2014-07-24 15:40:13
