
热带海草海菖蒲果实、种子形态及幼苗生长特征
2023-05-19 01:49蔡嘉丽史云峰李照阳赵牧秋罗慧珏史志强韩秋影
种子 2023年2期
蔡嘉丽, 史云峰, 李照阳, 赵牧秋, 罗慧珏, 史志强, 韩秋影
(1.海南热带海洋学院崖州湾创新研究院, 海南 三亚 572022;2.海南省现代化海洋牧场工程研究中心, 海南 三亚 572022;3.海南省近岸海洋生态环境过程与碳汇重点实验室, 海南 三亚 572022)
海草是生长在温带与热带海洋沿岸的单子叶植物,成片分布的海草形成海草床[1]。海草床作为海岸带天然生态屏障,在红树林与珊瑚礁生态系统之间起着纽带作用,能促进海洋元素循环,净化水质并稳固海床,为多种海洋生物提供产卵、索饵等重要生境[2],每年可为全球提供近2万亿美元的经济价值[3]。然而20世纪以来,由于全球气候变化和人为干扰的影响,海草床正在以前所未有的速率退化和缩减[4]。因此,加强针对海草的研究并开展海草床的保护和恢复已刻不容缓。
海菖蒲(Enhalusacoroides)是一种水鳖科海草,雌雄异株,可通过水媒传粉,主要分布于印度洋和西太平洋区域[5]。海菖蒲于七千多万年前由陆地被子植物进化而来,并在几百万年前重新进入海洋并形成一套对海洋环境特有的适应策略[6],如带状的叶片结构、特殊的光能利用模式、可漂浮的丝状花粉、丰富的单宁细胞、较强的化感作用等[7]。与其他海草一样,海菖蒲主要通过根状茎进行无性克隆生长,其根状茎节间长很短,仅为5 mm,水平扩张速率缓慢,每年仅0.9~5.0 cm[8]。Jover等[9]研究表明,有性繁殖在维持海草种群的遗传多样性方面起着重要作用。但由于海菖蒲成功的有性繁殖较为罕见,且其频率和强度都不可预测[10],迄今为止,人们对海菖蒲果实、种子和幼苗的形态特征及其生态适应意义知之甚少。在我国海菖蒲主要分布于海南陵水、琼海和文昌沿岸海域[11],这些分布区域的底质类型差异显著,陵水海草床的底质为潟湖沿岸冲积形成的细砂质硅酸盐,而琼海和文昌海草床底质则为珊瑚礁坪向陆侧的钙质珊瑚砂[12]。不同类型底质由于具有不同的有机质含量和氧化还原电位,可能会对海草的有性繁殖产生重要影响。但截至目前,有关不同底质类型对海菖蒲种子萌发和幼苗生长影响的研究鲜少报道。
鉴于此,本研究拟探讨热带海草海菖蒲果实、种子和幼苗的形态特征及其生态适应意义,同时探索底质类型对海菖蒲种子萌发和幼苗生长的影响,以期为深入剖析海菖蒲有性繁殖策略以及应用种苗法修复海菖蒲草床提供参考。
1 材料与方法
1.1 果实采集
成熟的海菖蒲果实于2021年12月采自海南省陵水县黎安潟湖南侧海草分布区(18°24′46′ N,110°03′18′ E),用海水清洗果实表面杂质和附着物后放入装有原位海水的泡沫箱运回实验室。
1.2 果实形态特征观测
观察海菖蒲果实形态,分别测定其直径(取最宽处)和高。用吸水纸吸除果皮上残留的海水并称重(记为m果实),然后用钢针刺入果实柄端,将果实全部压入盛有一定体积(V0)海水的量筒中,并读取压入后液面位置(V1),由公式ρ果实=m果实/(V1-V0)计算果实密度[13]。
将果实放入装有海水的烧杯中,观察并记录其自顶部裂开并释放种子的过程。待48 h内果实全部自然开裂后,将果皮取出并分别测定其含水率和碳(C)、氮(N)含量(Elementar,vario EL cube,德国)。
1.3 种子形态特征观测
观察海菖蒲种子形态,分别测定其直径(取最宽处)和高。随机选择3~5粒种子为1组,用1.2的方法测定该组种子的平均密度ρ种子。
将1粒种子贴近水面轻轻放入1 L海水的量筒(水柱长度记为L水柱)中,记录种子下沉姿态及用时(记为t),由公式V下沉=L水柱/t计算种子下沉速率。
1.4 底质类型对海菖蒲种子萌发及幼苗生长的影响
试验用钙质沉积物取自海南省三亚市情人湾(18°13′10″N,109°29′20″E),风干后过2 mm筛备用,其碳酸盐含量为76.4%,粒径中值为378 μm。硅质沉积物取自海南省三亚市铁炉港(18°15′39″N,109°40′58″E),用1 mol/L盐酸浸泡过夜去除其中的碳酸钙,多次清洗至近中性,风干后过2 mm筛备用,其碳酸盐含量为0,粒径中值为270 μm。
分别取两种不同类型沉积物作为底质,平铺在水族箱(40 cm×20 cm×15 cm)底部(厚5 cm),选取大小均匀的海菖蒲种子[直径(1.5±0.1)cm]种入其中。播种时种子子叶端朝上,2/3埋入底质内,然后添加海水至水面距底质表面10 cm。每箱播种12粒种子,每种底质处理设3次重复。试验所用海水取自海南省三亚市附近海域,盐度30~33 psu;试验过程中使用小型造浪泵保持水族箱内海水缓慢流动;在培养期间,每3 d更换一次海水。
分别于播种后的第14、56、98、140、182天,从各箱取海菖蒲幼苗,并将其分割为叶、残留种子和根3部分(如图1 E),分别测定其形态指标、重量、含水率以及C、N含量。海菖蒲幼苗根部所粘附的砂粒(如图1 E)经自来水冲洗后,仍不脱落的用镊子小心全部取下,烘干并称重作为根系粘砂量。
1.5 数据处理
试验数据以平均值±标准差(SD)表示,应用Microsoft Excel 2019和IBM SPSS Statistics 26.0软件进行数据统计分析和绘图,不同底质处理海菖蒲幼苗之间的差异显著性采用独立样本t检验(p<0.05)。
2 结果与分析
2.1 海菖蒲果实形态与基本生物学特征
海菖蒲植株具有被褐色纤维束包裹着的粗壮根状茎,从根状茎上长出很多乳白色细根;其叶呈带状,果实附着在螺旋状的花梗上(图1 A)。海菖蒲果实呈绿色,卵圆形,上端有尖状的喙,下端有两瓣分开的花被,果皮上有密集的突起(图1 B)。果实直径(41.57±5.54)mm,高(61.45±11.49)mm;果实质量(33.11±8.92)g,其中果皮占(59±4)%,种子占(41±4)%;果实密度为(0.90±0.04)g/cm3,低于海水。在海水中,成熟的果实会从顶端的喙处裂成6~9个不规则瓣片(图1 C),而果皮基部仍然相连。果皮含水率(87.58±1.86)%,C含量(15.48±3.25)%,N含量(0.99±0.21)%。每个果实含有3~16粒不等的种子。

注:A为带有成熟果实的海菖蒲植株;B为海菖蒲成熟果实;C为开裂后的海菖蒲果皮和种子;D为海菖蒲种子形态结构;E为海菖蒲幼苗。图1 海菖蒲果实、种子和幼苗形态结构Fig.1 Morphology and structure of E. acoroides fruits, seeds and seedlings
2.2 海菖蒲种子形态与基本生物学特征
海菖蒲的种子为黄绿色,呈上小下大的锥形结构,顶部为子叶和叶原基,底部为下胚轴和胚根(图1 D)。种子直径(14.23±1.38)mm,高(14.76±2.22)mm;质量(1.15±0.30)g,密度(1.17±0.07)g/cm3;含水率(72.67±0.26)%;C含量(35.20±0.24)%,N含量(2.09±0.14)%。当其刚从果实释放到海水中时,外部被一层半透明膜质种皮所包裹,膜质种皮与种子间充满气泡,导致种子漂浮于海水表层(图2 A-1、2 A-2)。这层膜质种皮一般会在24 h内破裂并与种子本体分离,此时种子会由于密度高于海水而下沉(图2 A-3)。下沉过程中,种子由于具有锥形结构和不均匀的密度,总是保持子叶端向上的状态,直至沉降至水底(图2 B 1~4)。经测定,海菖蒲种子在海水中的沉降速率为(87.00±0.62)cm/s。

注:A-1为海菖蒲果实刚刚开裂释放出的种子,种子被一层半透明膜质种皮所包裹,在海水中呈漂浮状态;A-2为A-1的俯视图; A-3为种子释放24 h后,膜质种皮破裂并与种子分离,种子下沉至容器底部;B-1~B-4为海菖蒲种子在水柱中的下沉状态,其下沉过程中一直保持子叶端向上的姿态。图2 海菖蒲种子的漂浮和下沉Fig.2 Floating and sinking of E. acoroides seeds
2.3 底质类型对海菖蒲种子萌发及幼苗生长的影响
试验期间,播种的所有海菖蒲种子全部萌发,萌发率100%。培养过程中,海菖蒲幼苗叶片数为4~6片,且不同底质间没有显著差异(图3 A)。培养前期(≤56 d),幼苗叶片生长较为缓慢且在不同底质处理间无显著差异;56 d后,叶片生长开始加快,并且呈现出硅质底质处理叶长和叶宽显著高于钙质底质处理的趋势(图3 B、C)。培养过程中,海菖蒲叶片含水率在74.6%~85.5%范围内波动(图3 D);C含量从培养前期(56 d)的30%左右降低到培养中后期(98~182 d)的23%以下,且在培养后期(182 d)钙质底质处理叶片C含量显著高于硅质底质处理(图3 E);N含量从培养前期(56 d)的1.8%左右降低到培养中后期(98~182 d)的1.3%以下,且不同底质处理间无显著差异(图3 F)。
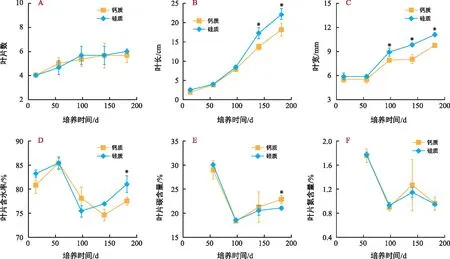
注:“*”表示差异显著(p<0.05)。下同。图3 试验期间海菖蒲幼苗叶片数量、形态、含水率和碳、氮含量的变化Fig.3 Changes of number, morphology, rate of water content, carbon and nitrogen content of the leaves of E. acoroides seedlings during the experiment
如图4 A、B,试验过程中海菖蒲幼苗的根数逐渐增加,根逐渐增长;在培养开始后的前98 d,幼苗根系发育(根数和根长)在不同底质处理间无显著差异;培养后期(≥140 d)硅质底质处理根数和根长均显著高于钙质底质处理。试验期间,幼苗根系含水率在75.3%~87.9%之间,且随培养时间的延长呈逐渐降低趋势(图4 C)。幼苗根系C含量由56 d时的32.0%~33.0%降低到140 d时的16.2%~18.8%,而后又升高到182 d时的18.4%~21.2%,且在培养后期(140~182 d)钙质底质处理显著高于硅质底质处理(图4 D)。培养过程中,幼苗根系N含量从56 d时的2.1%持续降低到试验结束时的0.4%以下,且不同底质处理间无显著差异(图4 E)。
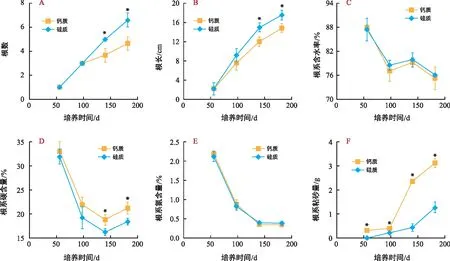
图4 试验期间海菖蒲幼苗根的数量、长度、含水率、碳、氮含量以及粘砂量的变化Fig.4 Changes of number, length, rate of water content, carbon and nitrogen content and sand sticking amount of the roots of E. acoroides seedlings during the experiment
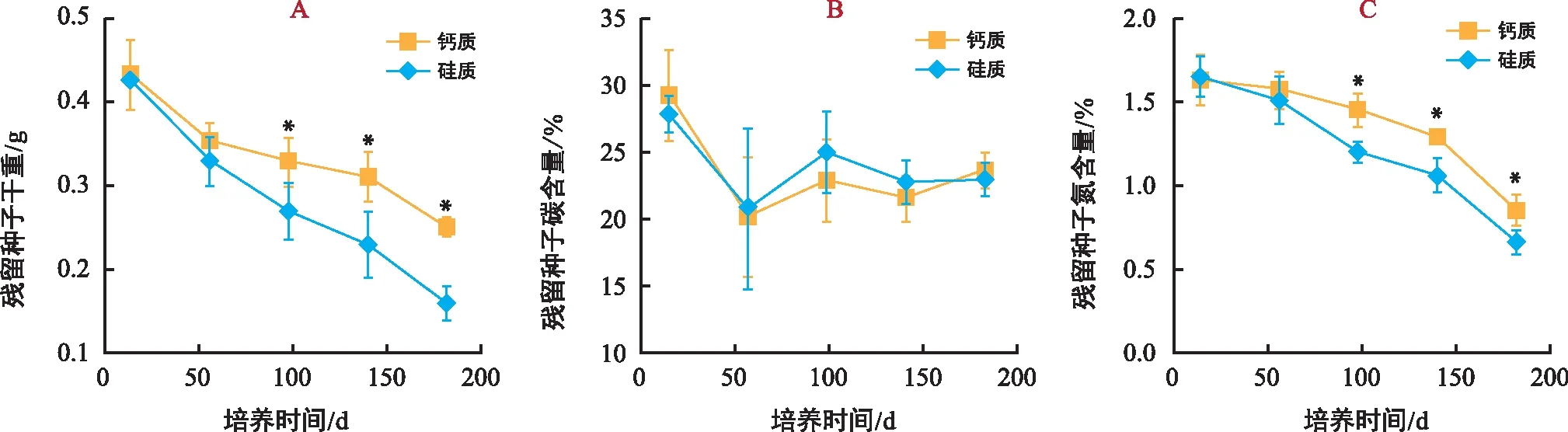
图5 试验期间海菖蒲残留种子干重及碳、氮含量的变化Fig.5 Changes of dry weight, carbon and nitrogen content of the residual seeds of E. acoroides during the experiment
伴随着海菖蒲幼苗根系的生长,在种子基部开始长出浓密的丝状根毛(图1 E)。这种丝状根毛能够粘附底质中的细小砂粒,水冲不掉,但两种不同底质处理的根系粘砂量存在显著差异(图4 F)。在第56天,钙质底质处理幼苗开始出现根系粘砂现象(0.31±0.02)g,但此时硅质底质处理根系尚无粘砂;培养的第98、140、182天,生长在钙质沉积物中的海草幼苗根系粘砂量分别是硅质沉积物处理的1.9倍、5.5倍和2.6倍。
培养期间,海菖蒲残留种子的干重和N含量持续降低(图5 A、C)。从14~182 d,硅质底质处理残留种子干重和N含量的平均降低速率分别为1.57 mg/d和0.059‰/d,显著高于钙质底质处理(1.09 mg/d和0.046‰/d)。试验过程中,不同处理残留种子的C含量呈前期(14~56 d)下降中后期(56~182 d)稳定的趋势(图5 B),且不同底质处理间无显著差异(p>0.05)。
3 讨论与结论
为了适应海洋环境和保持种群的遗传多样性,在漫长的进化过程中,海菖蒲获得了多样化的有性繁殖生态策略[14]。漂浮的果实和种子随海流向外扩散是海菖蒲种子传播并开拓新领地的重要方式,而漂浮状态的持续时间是其充分扩散的保障[15]。本研究发现,成熟的海菖蒲果实密度低于海水,其脱离母体后在海水中浸泡一段时间(48 h内)会自然开裂,而释放的种子尽管密度较高(1.17±0.07)g/cm3,但被含有大量气泡的半透明膜质种皮所包裹,释放后仍可保持漂浮状态,直至膜质种皮腐烂破裂释放气泡后种子方可下沉。这就为种子的充分扩散提供了时间保障。Lacap等[16]通过模型测算了不同天气条件下海菖蒲果实和种子的扩散距离,认为在正常天气条件下其散播距离在60 km左右,而台风期间其散播距离可达400 km。
海菖蒲种子具有上小下大的锥形结构,纵切面的颜色分布说明其顶端和基部存在明显的质地差异。这种结构和质地特征决定了海菖蒲种子在下沉过程中以及沉降到海床后总能保持直立姿态而不是随海流而滚动,使其基部与海底底质充分接触,进而确保根系的正常发育和种苗着床。海菖蒲幼苗初生根生长的同时,种子基部开始长出浓密的丝状根毛并粘附一定量的砂粒,也能够起到增强幼苗对底质的附着力,防止幼苗在波浪和海流环境中流失,提升有性繁殖成功率的作用。海草幼苗的丝状根毛可能是由种子内的糊粉层细胞发育而来[17]。本研究结果还表明,这种丝状根毛更倾向于粘附钙质材料而非硅质材料,原因可能是其通过分泌酸性化学物质将其粘附在碳酸盐砂粒的凹槽和细孔上,而这些分泌物对硅质沉积物的主要成分不能起到溶解作用。
本研究发现,当海菖蒲种子尚在果实内未释放前,其幼苗的叶片已开始萌发,说明海菖蒲的有性繁殖采取无种子库、无休眠期的胎生策略[18]。种子质量是种子储能大小的决定因素,一般小型种子能够维持幼苗生长的能力较低,而大型种子能够更充分保障幼苗初始生长过程所需的营养和能量[19]。不同种类的海草种子尺寸相差近2个数量级,从喜盐草属(Halophila)[20]的0.2 mm到波喜荡草属(posidonia)[21]的20 mm不等。本研究所获得的海菖蒲种子直径为(14.23±1.38) mm,说明其有性繁殖倾向于“大种子”策略,这可能是由于海菖蒲生长在营养较为缺乏的热带海区,其幼苗生长更多地依赖于种子的养分储备而不是通过幼苗的光合作用[22]。该繁殖策略与另一种热带大型海草大洋波喜荡草(P.oceanica)比较类似,其种子在萌发8个月后仍可为幼苗提供营养[23]。本研究结果也表明,在6个月的培养过程中,残留种子的干重和N含量持续下降,说明种子内部的营养和能量储存一直支持其幼苗的生长发育。
底质类型对海草的种子萌发和幼苗建成具有重要影响[24]。刘松林等[25]研究表明,沉积物粒径增大会显著降低海菖蒲幼苗的存活率和生长速率。Nugraha等[26]研究表明,海草幼苗在泥沙底质中比在珊瑚砂底质中生长得更快。本研究结果表明,海菖蒲幼苗在硅质底质中的生长速率明显高于钙质底质,其原因一方面可能是硅质沉积物较钙质沉积物含有更多的营养物质;另一方面可能是硅质沉积物粒径较小,质地较为松软,幼苗在扎根时不需要消耗太多能量,因此其根系生长速率更快并能从沉积物中吸收更多养分。
综上,海菖蒲为了提高在流动的海洋环境中的有性繁殖成功率,其果实、种子和幼苗进化出了一系列形态和生理策略。胎生缩短了海菖蒲种苗的发育时间,大型种子为幼苗的快速生长积累了更多营养和能量。成熟果实密度低于海水,果实脱离植株后在海水中漂浮一段时间后开裂,由膜质种皮包裹的种子释放后仍呈漂浮状态,这些特征为种子的有效扩散创造了条件。种子呈上小下大的锥形结构且下重上轻,幼苗根系发育时产生大量能够粘附钙质砂粒的丝状根毛,这些特征保障了海洋环境中种子的稳定和幼苗根系对底质的有效附着。此外,研究还表明,沉积物类型不影响海菖蒲种子的萌发率;但与钙质沉积物相比,粒径较小、质地松软的硅质底质更有利于海菖蒲幼苗的生长发育。
猜你喜欢
化工矿物与加工(2023年2期)2023-03-01
水生生物学报(2022年6期)2022-07-08
知识窗(2022年10期)2022-07-06
海洋通报(2022年2期)2022-06-30
学苑创造·B版(2022年10期)2022-05-30
海洋通报(2021年1期)2021-07-23
工程与建设(2019年1期)2019-09-03
智慧少年·故事叮当(2019年12期)2019-02-18
动漫界·幼教365(小班)(2018年5期)2018-05-14
海洋渔业(2017年5期)2017-11-07
