
基于Illumina MiSeq技术分析谷子根际丛枝菌根真菌群落多样性
2023-05-08 06:10段明
江苏农业科学 2023年6期
段 明
(山西农业大学实验教学中心,山西太谷 030801)
谷子[Setariaitalica(L.)Beauv]为禾本科狗尾草属,耐旱、耐贫瘠、水分利用率高[1]。目前,已成为单子叶作物抗逆研究的模式植物[2]。研究发现,丛枝菌根真菌(arbuscular mycorrhizal fungi,AMF)可使谷子根系高效利用土壤中磷元素,进而有利于产量的提高[3],AMF通过与宿主共生来提高植物的抗旱性。谷子抗旱性很强的特点除了与自身生理生态性质有关,其根系周围AMF的结构及多样性也不容忽视。因此,通过宏基因组测序分析不同品种谷子根系AMF种群特征,探索与谷子自身抗旱能力之间的相关性具有重要意义。AMF隶属球囊菌门,与绝大多数陆生植物都能形成普遍存在的内共生关系。在土壤微生物群体中,AMF的定殖依赖于特异信号的识别、传导和交换,这个过程涉及植物根细胞将菌丝的内化,进而AMF附着枝从根表面渗入到根内,将菌丝纵向蔓延在根系周围形成丛枝[4]。研究表明,AMF群落结构主要受非生物与生物两大因素影响。非生物因素如在森林、草原和沼泽生态系统中,AMF群落结构表现出显著差异[5]。生物因素主要指宿主本身对AMF的影响,植物会选择与之互利共生的AMF[6],但目前这种说法还存在争议。AMF最重要的生态功能就是与宿主建立共生关系来促进植株的生长,提高农业产量。研究发现,植物与AMF形成稳定的共生关系有利于它们的生存及对逆境胁迫的抗性。植物向AMF提供脂类以合成生命所需的脂肪酸;反过来,定殖于根系的AMF为植物提供水分和养分,促进对土壤氮磷及矿质营养的吸收和转运[7],从而促使植株生长发育,提高植株对胁迫的耐受性。此外,AMF对植株分子和生理生化过程的调控亦具有很大影响。研究表明,接种AMF后,植物的光合、渗透调节及抗氧化能力得到大幅提高,抗旱性与未接种的植物相比也显著增强[8];此外,植物体内抗旱相关基因的表达也受到AMF强烈的诱导[9]。近期一些研究表明,谷子根系中的AMF可有效提高对磷元素的吸收进而达到增产的效果[3],而接种根际固氮细菌也可显著促进狗尾草的生长发育[10],这2项研究均与谷子密切相关。谷子耐旱适应性强,然而,目前对谷子抗旱性的研究主要集中在基因表达调控方面。MiSeq高通量测序具有准确性高和通量大的特点,为谷子根际微生物的研究创造了条件,为此,采用高通量测序技术研究干旱胁迫下不同谷子品种根际AMF群落分布特征,以便充分发掘和利用AMF优势菌群,为谷子抗旱研究奠定基础。本研究通过宏基因组测序分析鉴定与干旱相关的丛枝菌根真菌,并解析干旱条件下谷子根系丛枝菌根群体的结构特征。
1 材料与方法
1.1 试验材料
谷子材料选用豫谷1号(YG)、沁黄(QH)、安陵(AN)3个品种。2018年,将3个品种种植于山西农业大学校外农作站旱棚中,苗期开始进行不浇水干旱处理,10 d后取样,通过抖根法至土壤不再下落,根际土壤即为黏在根系表面的土壤,去除土壤杂质,连同植物根系放于-80 ℃保存备用。3个品种进行3次重复,CK为不含谷子根系的土壤对照。
1.2 土壤宏基因组DNA的提取及检测
利用土壤专用的基因组试剂盒(天根)提取土壤中微生物基因组DNA,提取后放置于-80 ℃冰箱保存。
1.3 PCR扩增以及Illumina MiSeq测序
AMF的扩增采用引物AMV4.5NF(5′-AAGCTCGTAGTTGAATTTCG-3′)和AMDGR(5′-CCCAACTATCCCTATTAATCAT-3′)。扩增完成后,对PCR产物进行定量分析,在上海派森诺生物科技股份有限公司进行Illumina MiSeq测序。
1.4 高通量数据分析
利用FLASH软件与QIIME软件对测序结果进行比对分析。
2 结果与分析
2.1 土壤理化性质的测定
由表1可知,试验土壤在正常浇水与干旱条件下的含水量分别为7.64%和3.69%,测定结果显示,土壤的pH值分别为8.46和8.09,按土壤pH值分级标准,试验土壤呈碱性。按照土壤养分等级标准观察,速效氮含量为61.4~70.4 mg/kg,属于四级水平,速效磷含量为22.42~28.12 mg/kg,属于二级级水平,速效钾含量为183.6~185.3 mg/kg,属于二级水平,有机质含量为21.62~25.05 g/kg,属于三级水平。
2.2 OTU统计及分类学分析
按97%序列相似度进行归并和操作分类单元(OTU)划分,由图1可知,谷子根际AMF群落的多样性及种群丰度显著低于CK。统计结果表明,谷子根系与CK土壤中OTU数量在1 632~4 290之间,鉴定出126种AMF。根据试验要求,对不同样本绘制Venn图,分为2组:正常浇水谷子(YG、AN、QH)与干旱处理谷子(dYG、dAN、dQH)。由图2、图3可知,CK与YG、AN、QH进行聚类,共有OTUs 213个;CK与dYG、dAN、dQH进行聚类,共有OTUs 154个。



2.3 谷子根际AMF群落α-多样性分析
AMF的种类及丰度存在很大变异,通过检测谷子样品中AMF的丰富度和多样性,可根据检测结果计算得出Chao1指数、ACE指数、辛普森指数和香农指数。由表2可知,CK样品的Chao1指数为734.12,ACE指数为751.29;而谷子根际样品的Chao1指数在218.50~489.12之间,ACE指数在222.18~498.00之间,表明CK样品中AMF的丰富度与微生物群落多样性最高。

表2 不同样品中的AMF群落多样性指数
2.4 各分类水平的AMF群落数统计
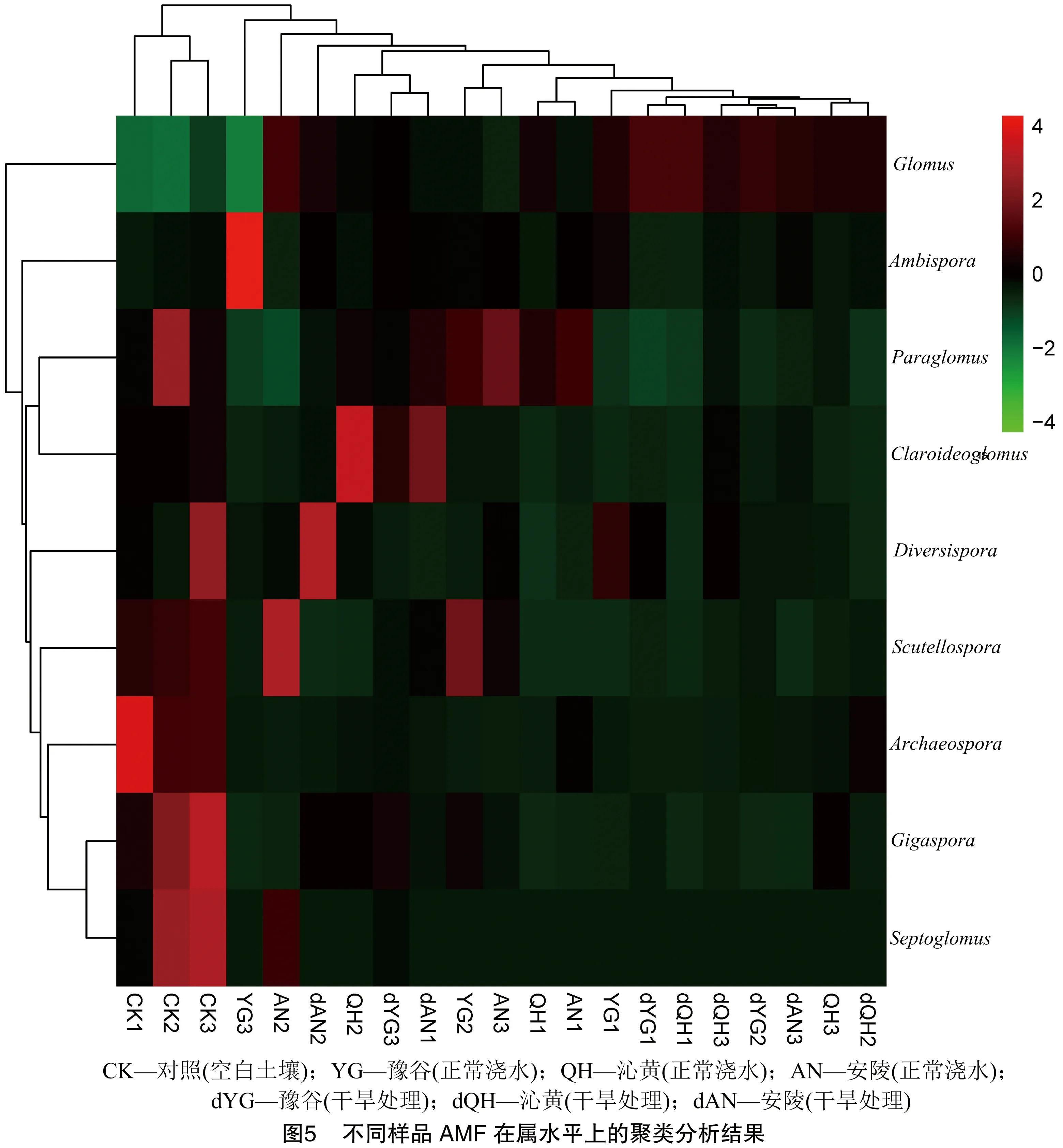
不同的分类学结果表明,在门水平上,所有样品序列都归入球囊菌门;在纲水平上,所有样品序列注释到明确分类地位的有3个纲,分别为球囊菌纲、类球囊菌纲和原囊菌纲;在目分类水平上,所有样品共注释到明确地位的有4个目,分别为球囊霉目、类球囊霉目、原囊霉目和多样孢囊霉目;在科分类水平上,注释到明确分类地位的有7个科,分别为球囊霉科、类球囊霉科、双型囊霉科、原囊霉科、巨孢囊霉科、近明球囊霉科和多样孢囊霉科。由图4可知,在属分类水平上,从CK以及谷子根际土壤样品中注释到明确分类地位的共有9个属,依次是球囊霉属、类球囊霉属、双型囊霉属、原囊霉属、近明球囊霉属、巨孢囊霉属、盾巨孢囊霉属、多样孢囊霉属和隔球囊霉属。其中,原囊霉属在CK中占优势,类球囊霉属在谷子根际样品中占优势。

使用R软件,对丰度较高的属进行聚类分析和绘制热图。由图5可知,红色为样本中丰度较高的属,绿色为丰度较低的属。
2.5 谷子根际AMF群落β-多样性分析
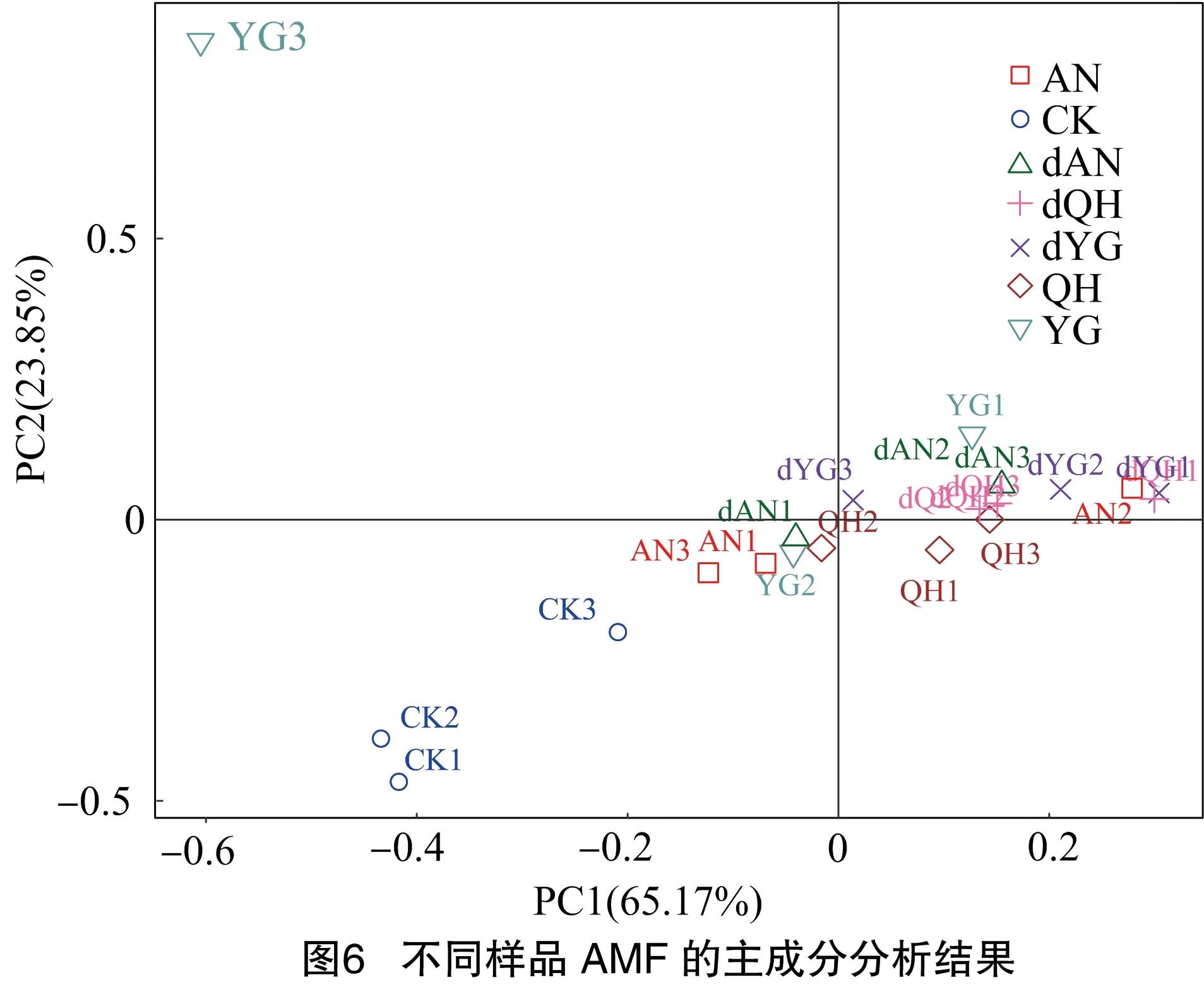
主成分分析(PCA)是基于欧氏距离评价样本之间的相似度,以不同样本中的AMF属进行作图,得到PCA图。由图6可知,主成分1(PC1)和主成分2(PC2)贡献率分别为65.17%、23.85%,可较好地反映样本间种群结构差异的影响因素。
其中,○组代表单纯土壤样品,CK1、CK2距离较近,但与CK3距离较远;□组代表正常浇水条件下的AN谷子根际样品,AN1、AN3距离较近,但与AN2距离较远;◇组代表正常浇水条件下的QH谷子根际样品,QH1、QH3距离较近,但与QH2距离较远;▽组代表正常浇水条件下的YG谷子根际样品,YG1、YG2距离较近,但与YG3距离较远;△组代表干旱条件下的AN谷子根际样品,dAN1、dAN2、dAN3距离较近;+组代表干旱条件下的QH谷子根际样品,dQH1、dQH2、dQH3距离较近;×组代表干旱条件下的YG谷子根际样品,dYG1、dYG2、dYG3距离较近。由此可得出以下结论:第一,CK与谷子根际样品能够较好地分开,表明寄主有无是影响AMF群落结构差异的主要因素;第二,同一谷子品种、不同处理以及同一处理、不同谷子品种并不能完全聚在一起,表明处理条件和品种类型是影响AMF群落结构差异的次要因素。
2.6 AMF系统发育树分析
为直观清晰地反映各分类单元在样本中的丰度及进化信息,利用软件MEGAN构建种群分类等级树。由图7可知,CK中AMF主要是原囊霉属(Archaeospora)和巨孢囊霉属(Gigaspora);YG中AMF主要是双型囊霉属(Ambispora);球囊霉属(Glomus)和类球囊霉属(Paraglomus)在所有样品中均匀分布。
2.7 样本AMF组成的差异分析
由表3可知,CK与谷子根际样品相比,在属水平上丛枝菌根存在显著差异;干旱处理后,YG与QH品种根际丛枝菌根在属水平上具有差异,AN品种根际丛枝菌根群落未发生显著差异。由图8可知,样本组间最显著的分类单元,分别为双型囊霉属、原囊霉属、近明球囊霉属、巨孢囊霉属、球囊霉属、类球囊霉属、盾巨孢囊霉属、隔球囊霉属。
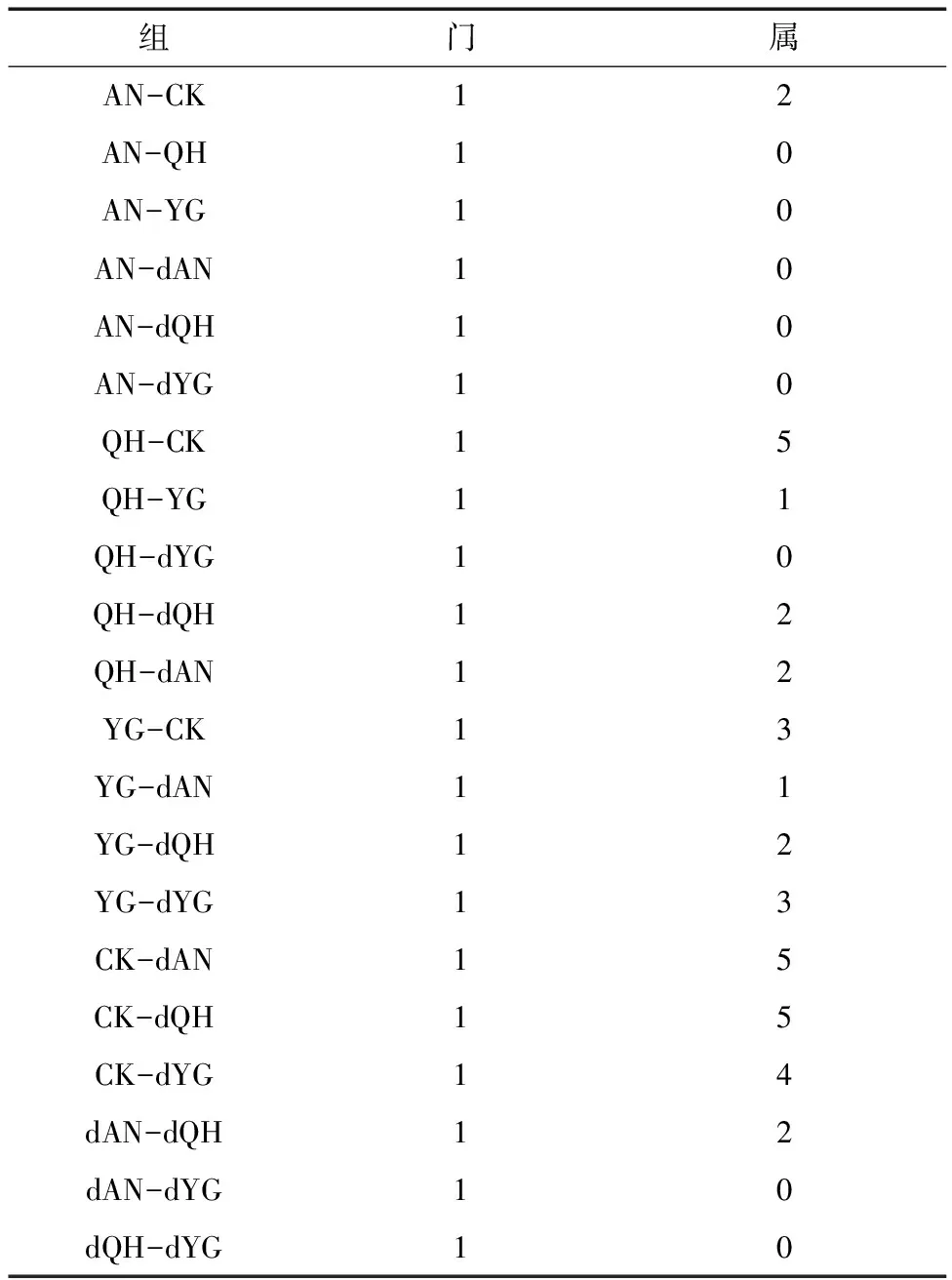
表3 样本间分类学组成的差异分析结果
3 结论与讨论
植物与微生物之间的互利共生关系源于胞内信号物质的交换。植物根系通过释放独脚金内酯(SLs)可诱导AMF产生分枝,并促进菌丝在根部的定殖[11]。AMF的定殖依赖于特异信号的识别、传导和交换,这个过程涉及植物根细胞将菌丝内化,进而AMF附着枝从根表面渗入至根内,将菌丝纵向蔓延在根系周围形成丛枝[12-13]。
土壤作为一种复杂的基质是植物赖以生存的场所[14],而根际是植物根系与土壤的交界面,是大量微生物的栖息地,微生物受益于植物根系的代谢产物[15]。AM是自然界分布较为广泛而古老的一种共生菌[16],它的共生体现在营养物质的交换,植物向它提供约20%的光合产物,反过来它帮助植物有效地吸收土壤中的矿质元素及水分[17-18],它们的相互作用影响着植物的生长发育、养分吸收及对胁迫的耐受性[19]。此外,土壤的组成及结构对植物生理及根系分泌物也有调控作用,不同植物根系形态的不同、分泌物数量及种类的不同均会对植物根系表面的微生物产生选择性,改变微生物群落组成[20]。

前期Tang等证明,YG与AN分别为抗旱性强与干旱敏感的谷子品种,QH介于两者之间;干旱条件下,与YG相比,AN的生长显著受到抑制;而在正常浇水条件下,AN的光合能力与生长状况要好于YG[2]。影响植物的抗旱性因素很多,诸如基因型、环境及植物与微生物的共生互作等。为此,本研究选择YG、AN和QH等3个抗旱性不同的材料,利用高通量测序技术对谷子根际丛枝菌根真菌群落进行了研究。多样性分析结果表明,CK与谷子根际丛枝菌根真菌多样性指数及丰度指数存在显著差异,宿主谷子的根际丛枝菌根真菌丰度指数和多样性指数均低于CK,特别是原囊霉属、巨孢囊霉属和隔球囊霉属显著减少,这表明宿主的生理代谢及根系分泌物在一定程度上影响了丛枝菌根真菌的分布特征。此外,无论正常浇水还是干旱条件下,这些指数在不同谷子品种之间存在普遍的相似性,并未表现出明显的差异,只是少数菌属在品种间表现出独特性。这表明在干旱条件下,根系的生理代谢依然维持在较高的水平,与正常浇水条件下的群落特征基本一致。分析原因可能有3点:第一,在自然生态系统中,植物往往能与AMF形成互惠互利的关系,这种关系的形成有利于提高植物抵抗各种胁迫的能力,是依赖于植物根系的菌丝协助获取矿质营养及水分而实现的。与其他植物不同,谷子耐旱、耐贫瘠,水分利用率高,而根系是其重要基础。因此,在干旱胁迫下,AMF依然保持种类多样的特征,与谷子建立很好的共生。第二,研究表明不同生态型的土壤类型能强烈影响AMF的群落结构及多样性水平[4],本试验土壤类型为基本农田用地,土壤呈弱碱性,根据有机质及氮磷钾含量分级标准来看,营养状况属于中高级,较为适宜作物生长。谷子根际AMF是否受到土壤理化性质的影响有待进一步探讨。第三,干旱的设置,造成品种间AMF的相似是否受到干旱时间和程度的影响有待进一步研究。Maestre等对除南极洲以外各大洲80个旱地的微生物多样性进行了全面分析,结果表明,随着干旱程度的增加,土壤微生物的多样性及丰度出现降低,这些结果主要是由于干旱对土壤有机碳含量造成的负面影响;干旱条件会改变土壤细菌群落结构,绿弯菌门和α-变形菌相对丰度增加,而酸杆菌门和疣微菌门的丰度减少,试验结果与以往研究报道情况相反,土壤pH值不是影响细菌多样性的主要因素,真菌群落以子囊菌为主[21]。然而,目前本研究结果表明,干旱条件下,谷子根系丛枝菌根真菌群落结构及分布并未发生显著变化,是否由于宿主与真菌互作的关系或谷子的抗旱性特征有关有待进一步研究。

由图6、图7可知,球囊霉属和类球囊霉属在所有样本中分布较为均匀,表明这2个AMF属受谷子根系分泌物影响较小;原囊霉属和巨孢囊霉属在谷子根际样本中含量很低,明显低于CK,表明这2个AM属受谷子根系分泌物影响较大。
综上所述,本研究对3个谷子品种(豫谷、沁黄、安陵)在正常生长与干旱条件下AMF的群落组成试验中,发现干旱条件下,AMF群落组成及丰度并未发生显著变化,与谷子本身耐旱有很大关系。在今后的研究中,应当从干旱程度及土壤选择等因素分析对AMF群落的影响,以期发现谷子抗旱与AMF的关系。本研究结果为进一步探索AMF的多样性,选择及鉴定合适的AMF菌种进行定殖谷子奠定了基础。
猜你喜欢
当代陕西(2021年21期)2022-01-19
中国土壤与肥料(2021年5期)2021-12-02
中成药(2018年3期)2018-05-07
天然产物研究与开发(2018年3期)2018-05-07
中国蔬菜(2016年8期)2017-01-15
现代农业(2015年5期)2015-02-28
植物营养与肥料学报(2011年5期)2011-11-06
植物营养与肥料学报(2011年6期)2011-10-24
植物营养与肥料学报(2010年3期)2010-11-16
植物营养与肥料学报(2010年6期)2010-10-26
